Female-led infanticide in wild chimpanzees
2
0
0
Texte intégral
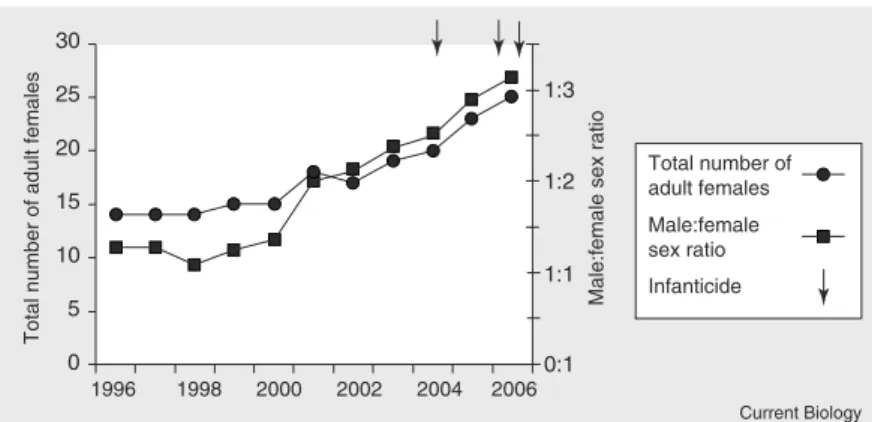
Figure
Documents relatifs