Develop. Growth and Differ., 28 (I), 75-84 (1986)
A Freeze-Fracture Study of the Cortex of
Xenopus
laevis
Eggs
(amphibian egg/cortical endoplasmic reticulum/cortical granules/Freeze fracturelmembrane junctions)
MICHEL CHARBONNEAU*,**, ROBERT D. GREY*, RONALD J. BASKIN" AND DANIEL THOMAS*'s
* Department of Zoology, University of California Davis, Califonia 95616 and ** Laboratoire de Cytologie expdrimentale. Universite' de Rennes,
35042 Rennes Cedex, France
The organization of the cortex of Xenopus laevis eggs was investigated by freeze-fracture electron microscopy. The cortical endoplasmic reticulum (CER) formed a network surrounding and inter- connecting the cortical granules. It formed junctions with the plasma membrane and was confluent with the ER in subcortical regions. Intramembranous particles (IMP,) were only present in the P face of the CER, the E face being apparently devoid of pits and particles. Arrays of densely packed IMP,, having a mean diameter of 17 nm, were restricted to the microvillar region of the plasma membrane. The cortical granule membrane also contained IMP, (mean diameter, 21 nm) that were sparsely and randomly distributed. Several types of cortical granule seemed to exist based on an analysis of the distribution of the different IMP sizes.
Many of the early events of fertilization and activation of development in animal eggs are mediated by membrane systems in the cortex of the egg. In Xenopus Iaevis (anuran amphibian), as in other animals, these events are initiated by the contact and subsequent fusion of the plasma membranes of the two gametes, which triggers a rapid transient depolarization, termed the fertilization potential, that results from an increase in the chloride conductance of the egg plasma membrane (1, 2). The fertilization potential is followed by the release of calcium ions in the cytoplasm (3) resulting in exocytosis of cortical granules in a wavelike pattern that starts at the site of sperm entry (4).
The only previous report on the properties of the membrane components in the cortex of anuran eggs was confined to maturation (5). The cortical endoplasmic reticulum (CER) was first identified in thin sections of eggs (4) and described in greater detail by CAMPANELLA and ANDREUCCETTI (6), who suggested that it was a likely candidate for the storage and release of the calcium ions responsible for the early events of activation. Support for this hypothesis comes from the temporal correlation between the first appearance of the CER during oocyte maturation and the onset of calcium-dependent activatability (7, 8).
In addition, the CER makes junctions with the plasma membrane that are similar in morphology to those that join the terminal cisternae of sarcoplasmic reticulum with the T system (9). Finally, calcium has been reported to be detectable in the CER soon after activation of Xenopus eggs (10).
We have therefore examined the cortex of Xenopus eggs by freeze-fracture electron microscopy in order to understand better the three-dimensional organization of the CER and its relationship to other membrane systems. The present freeze-fracture study also provides
76 M. CHARBONNEAU ET AL.
an improved description of the junctions formed between the plasma membrane and the CER with respect to previous studies on thin sections of whole eggs (9) and of isolated cortices (8). We also report our observations of intramembranous particles (IMP,) in the cortical granule and plasma membranes and their spatial distribution.
MATERIALS AND METHODS
Adult females of Xempus laevis were injected intraperitoneally with 100 I U human Chorionic Gonadotropin (hCG) (Sigma Chemical Company, St. Louis, MO) and then 48 hr later with 800 to 1000 IU hCG. Mature jellied eggs were squeezed from the female 8 to 10 hr later into 1/10 OR-2 (11). After 10-15 min equilibration in the solution, the eggs were f3ed and the jelly layers and the vitelline envelope were manually removed during fixation. Some eggs were completely dejellied prior to fixation by gentle swirling in 2% cysteine, in 1/10 OR-2, pH 7.8, for 8-10 min before washing extensively for about 30 min by transferring them into large volumes of 1/10 OR-2, pH 7.4. Fixation time was 4-6 hr at 4°C in 2.5 % glutaraldehyde, either in 0.1 M phosphate buffer or in 1/10 OR-2. The eggs were then washed for about an hour in buffer and transferred to 20% aqueous glycerol for 4 hr or overnight at 4°C. The eggs were first frozen in liquid Freon 22 cooled with liquid nitrogen, then stored in liquid nitrogen. The specimens were fractured in a Balzers apparatus at -110°C; some were allowed to etch for 1-2 min. Platinum/carbon replication used an electron beam evaporation unit positioned at 45' and was followed by application of carbon from overhead. Replicas were cleaned in a mixture of 4% potassium bichromate, 53% HzS04, and 43% HzO, rinsed in distilled water, mounted on 300-mesh grids and examined in an Hitachi HU-I 1E electron microscope.
Particle size and density measurements were performed on micrographs at actual magnifications ranging from 100,000 to 150,000 diameters, using a digitizer (Houston Instruments) linked to a microcomputer (Zilog MCZ). For observation of thin sections of egg cortex, mature jellied eggs were fixed for 2 hr in 2.5 % glutaraldehyde plus 0.1 % tannic acid in 0.1 M phosphate buffer, washed in the buffer and post-fixed in 1 % OsO,. After the last wash, the samples were dehydrated in graded acetone and embedded in Epon-Araldite; thin sections were observed with a Hitachi HU-11E electron microscope.
RESULTS
Cortical Structures Revealed in Freeze-Etch Preparations
Fig. 1 illustrates the spatial relationships between the membrane-bound organelles in the cortex of a mature, unfertilized egg as observed in freeze-etch preparations. Immediately beneath the microvilli-studded plasma membrane are the prominent cortical granules which can be distinguished from other cortical organelles by their location, spherical shape, and size (1.5-2.5 pm). The cortical endoplasmic reticulum (CER), described in more detail in the next section, ramifies throughout the cortex. Pigment granules, mitochondria and yolk granules all tend to be positioned beneath the layer of cortical granules.
Fig. 1. Cortical region of an unfertilized egg of Xenopus [aevis, fractured approximately perpendicular to the plane of the plasma membrane, from which microvilli (Mv) extend. Cortical granules (CG) and cortical endo-
plasmic reticulum (CER) occupy much of the 1-1.5 pm of cytoplasm immediately beneath the plasma membrane. P, pigment granule. On all freeze-fracture micrographs, the arrow located near the figure number indicates the direction of shadowing.
Fig. 2. Higher magnification of the cortical area shown in the right-hand portion of Fig. 1. Particles can be seen in the membrane of the cortical granule. The extensive cortical endoplasmic reticulum (CER) extends a network around the cortical granule; both the P face (single arrows) and the E face (double arrows) of the CER can be seen. PM, plasma membrane. 49,900 x
.
FREEZE-FRACTURE OF X E N O P U S EGG
78 M. CHARBONNEAU ET AL.
Fig. 3-5. Examples of junctions (star) between the cortical endoplasmic reticulum (arrows) and the plasma membrane. Fig. 3 : The P face of the junction is shown in a region of the CER that expands laterally toward the cortical granules and centrally toward the subcortical cytoplasm. 52,500 x
.
Fig. 4: A junction region (star) is seen to the left of the cortical granule; a lateral expansion of the CER in a region between cortical granules lies to the right. 31,500 x . Fig. 5: The CER appears as a disconnected column that extends from the region of junctions (stars) with the plasma membrane to the areas juxtaposed to the cortical granule. 43,750 X.
Fig. 6 1 1. Organization of the egg cortex compared in freeze-fracture replicas (Figs. 6,8,10) and in thin sections (Figs. 7,9, 11). CER and arrows: cortical endoplasmic reticulum; asterisks: junctions between the CER and the plasma membrane; other symbols: as in Fig. 1 . Fig. 6: cisternae of CER completely surround the two cortical granules and make junctions (asterisks) with the plasma membrane. 18,250 x . Fig. 7 : an organization similar to that of Fig. 6 is observed in this sectioned material. 18,250 x . Fig. 8 a junction (asterisk) is seen to the right of the cortical granule; expansions of the CER lie around and between the two cortical granules and between the pigment granules. 21,900 x . Fig. 9: the junction located to the upper right of the left cortical granule and its extension to the area between the cortical granules are identical to the ones shown in Fig. 8. 2 1 , 9 0 0 ~ . Fig. 10 shows the two types of junctions already described (see Figs. 3-5 and text) and cisternae of CER interconnecting the plasma membrane and two cortical granules. 25,600 x . Fig. 11 : an extensive network of CER surrounds the cortical granules and makes junctions with the plasma membrane. 25, 600 x
.
80 M. CHARBONNEAU ET AL.
The Cortical Endoplasmic Reticulum (CER)
The CER is best observed in regions where it surrounds cortical granules. As in sectioned preparations, the CER associated with cortical granules appears as a fragmented series of vesicles, usually fractured to reveal the P face (Fig. 2), which contains IMP,; the E face appears to be free from both particles and pits. Whether the absence of pits in the E face is due to contamination resulting from long etching times, as has been demonstrated for sarcoplasmic reticulum (12), has not been determined. In regions between cortical granules, the CER appears to be larger, and fracture planes typically expose the smooth, particel-free E face (Figs. 2-5, 6, 8, 10). The anastomosing nature of the CER is shown in Fig. 12 which also illustrates the communication between the CER and ER in the subcortical regions of the egg.
Regions of close apposition between CER and plasma membrane (Fig. 3-5) resemble the junctions seen in thin sections (8, 9). Some of these junctional regions involve an irregularly shaped cisterna which is confluent with the CER (Figs. 3, 5). In many cases, however, the internal membrane in close apposition with the plasma membrane appears to be part of a simple, ovoid vesicle with no apparent connection with other components of the CER (upper left areas of both Figs.
4
and 5). However connections between these vesicles and the CER could lie beneath the fracture plane. We were unable to resolve any counterpart of the electron-dense “feet” seen in thin sections to join the CER and the plasma membrane.In order to further illustrate the characteristics of the CER and of the CER-plasma membrane junctions, we selected 6 areas of egg cortex from freeze-fracture replicas (Figs. 6, 8, 10) and from thin sections (Figs. 7, 9, 11). Such a comparison makes it easier to localize the junctions on the freeze-fracture replicas and to observe the relationships between the CER and the cortical granules.
Cortical Granules
The membrane that bounds the cortical granules contains sparsely distributed IMP, (ca. 256h70/pm2) that associate with the P face in fractured preparations (Figs. 2, 13). Measure- ments of 764 particles (in 37 cortical granules from six eggs, each from a different female) yielded a mean diameter of 21.2k3.2 nm (SD).
The diameters of IMP, in the membranes of 29 different cortical granules in a single egg
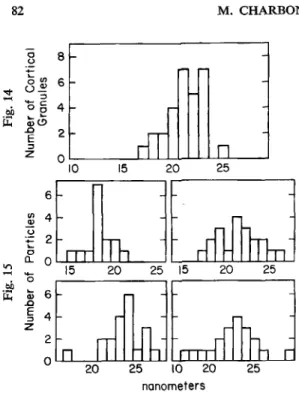
were measured in order to ascertain their homogeneity. A broad and asymmetric distribution of granules was found according to the mean diameter of IMP, (Fig. 14). Significant differences
were seen when the mean diameters of IMP, in these 29 granules were c0mpared.l In order to know whether the asymmetry of the distribution of the values of IMP, in the cortical granule population (Fig. 14) was also present at the level of each cortical granule, we selected data from
six of these cortical granules and performed the same statistical analysis. Fig. 15 illustrates the range and means of four of these samples. For any one granule, the distribution of IMP sizes was fairly symmetrical, and there was no statistically significant difference between IMP size. The asymmetry of the mean size distribution observed among the cortical granule popu- lation was due therefore to the existence of cortical granules with different IMP size classes. At least two types of cortical granules can be discerned on the basis of size, shape and morphology
FREEZE-FRACTURE OF XENOPUS EGG 8 1
Fig. 12. Brackets mark the disjointed columns of CER that extend from a junction with the plasma membrane to the subcortical cytoplasm. Symbols as in Fig. 1. 18,250X.
Fig. 13. P face of a cortical granule mem- brane containing intramembrane particles. 30,600 x .
a2 M. CHARBONNEAU ET AL. 2 0 25 15 0 15 2 0 25 15 20 25 6 4 2 2 0 25 10 20 25 nanometers
of contents in sectioned Xenopus eggs (4, 6 ) .
Fig, 14-15. Distribution of intramembrane pa rticle (IMP) diameters in the membrane bounding cortical granules, subdivided into 1 nm classes. F&. 14: Distribution of the mean values in 29 cortical granules from a single egg. Abscissa: IMP diameter (nm); ordinate: number of cortical granules. In this egg, 606 particles from 29 cortical granules were examined; mean diameters were 21.7k 1.8 nm (N=29) and 21.8b2.9 (N=606). The variations in IMP size observed between cortical granules were statistically significant as demonstrated by analysis of variance (see text). Fig. 15: Distribution of particle diameters in each of four different cortical granules from the same egg as in Fig. 14. Abscissa IMP diameter (nm); Ordinate: number of particles. Six cortical granules (data for only four of which are shown) were arbitrarily chosen from among the 29. The selection included two with a mean value in the range of 17-1 8 nm, two in the range of 21-22 nm and two in the range of 23-24nm. For all six granules, the variations observed in IMP size within each granule were not statistically significant, as demonstrated by a standard one-factor analysis of variance.
IMP, observed in unfertilized eggs of sea urchins (13, 14) are generally smaller than in
Xenopus, but the cortical granules are similar in these species with respect to density and lack of any obviously regular arrangment of particles.
Plasma Membrane
The plasma membrane of mature, unfertilized Xenopus eggs contains IMP, that associate with the E face in fractured specimens (Fig. 3-5) (see also ref. 5). IMP, are most numerous in the microvilli (1331 237 particles/pm2). IMP diameters measured 17.7f 3 nm (n =495 parti- cles, from 27 microvilli from 5 eggs, each from a different female) in microvillar regions of the
plasma membrane and 15.2 & 3 nm (n = 150, in 3 eggs, each from a different female) in the regions between microvilli. These sizes are significantly larger than the 10 nm average diameters reported by BLUEMINK et al. (5). The reasons for this difference are not clear, but may be due
to the different procedures used to prepare freeze fracture replicas.
DISCUSSION
To our knowledge, this paper is the first to report on the three-dimensional ultrastructure of the cortical region of a vertebrate egg by means of freeze-fracture. Although the anuran egg is a very popular material in developmental biology, this lack of data is probably due to technical reasons. The replica of a whole section of a freeze-fractured egg is very susceptible to breakage. Since the cortical portion of the cytoplasm is very small compared to the rest of the egg, replicas of it are very difficult to collect on the grids. Nevertheless, careful examination of the small broken replicas in the electron microscope can lead to the selection of regions of interest.
FREEZE-FRACTURE OF XENOPUS EGG 83
Knowledge of the precise organization of the cortex of the unfertilized egg is of basic interest in the comprehension of the mechanisms responsible for the transduction of the activating signal generated by the fertilizing sperm.
In addition to the numerous observations already carried out on thin sections of the anuran egg cortex, the present study reveals the existence of a number of new characteristics. (i) The microvillar regions of the plasma membrane contain regularly arranged and closely packed arrays of IMP,, whereas the IMP, in the regions between the microvilli are much less numerous and are randomly distributed. The existence of such membrane domains might be related to the presence of preferential sites of fusion between the sperm and the egg or, alternatively, to the fact that the microvilli, which elongate upon activation, might have a particular compo- sition enabling this process. (ii) The cortical granule membrane contains sparsely and randomly distributed IMP,; their size is homogeneous within a single granule but varies within the granule population of a single egg. The existence of several types of cortical granules on the basis of their membrane composition is to be regarded together with the previously reported regional differences in the granule matrix density (6, 4). (iii) The CER is shown here for the first time in three dimensions; its P face contains IMP, whereas its E face appears to be devoid of any
particle. Further characterization of the CER, preferably in the form of isolated vesicles, should reveal the importance of these IMP, which might represent some specific ER protein. (iv) Membrane junctions between plasma membrane and endoplasmic reticulum have been described only twice in eggs, once in whole eggs (9) and once in isolated cortices (8). In this latter study, the CER was shown to tightly adhere to the plasma membrane at junctional sites, whereas it did not retain any association with cortical granules. From the present work the possibility arises that the membrane junction may exist under diverse configurations. The improved description of the structure of these junctions compared to the original description (9) demonstrates that further characterization is needed before a precise role can be attributed to them.
We thank Misses F. de Sallier Dupin and G. Boguais for typing the manuscript and Mr. B. Morille for MC was supported by a research fellowship from the Ministere des assistance in preparing the illustration.
Relations Extkrieures, France.
1. 2. 3. 4. 5. 6. 7. 8. 9. 10. 11. REFERENCE !
CROSS, N. L. and ELINSON, R. P. (1980). Develop. Biol. 75, 187-198.
GREY, R. D., BASTIANI, M. J., WEBB, D. J. and SCHERTEL, E. R. (1982). Develop. Biol. 89, 475-484. BUSA, W. B. and NUCCITELLI, R. (1985). J. Cell Biol. 100, 1325-1330.
GREY, R. D., WOLF, D. P. and HEDRICK, J. L. (1974). Develop. Biol. 36, 4461.
BLUEMINK, J. G., HAGE, W. J., VAN DEN HOEF, M. H. F. and DICTUS, W. J. A. G. (1983). Eur. J. Cell Biol. 31, 85-93.
CAMPANELLA, C. and ANDREUCCETTI, P. (1977). Develop. Biol. 56, 1-10,
CHARBONNEAU, M. and GREY, R. D. (1984). Develop. Biol. 102,90-97. GARDINER, D. M. and GREY, R. D. (1983). J. Cell Biol. 96, 1159-1163.
ANDREUCCETTI, P., DENIS-DONINI, S., BURRINI, A. G. and CAMPANELLA, C. (1984). J. Exp. Zool. 229,
WALLACE, R. A., JARED, D. W., DUMONT, J. N. and SEGA, M. W. (1973).
, TADDEL, C. and TALEVI, R. (1984). J. Exp. Zool. 229, 283-293.
~ _ _ _
295-308.
84 M. CHARBONNEAU ET AL.
12. RASH, J. E., GRAHMA, W. F. and HUDSON, C. S. (1979). In: “Freeze Fracture Methods, Artefacts and Interpretations,” (J. E. Rash and C . S. Hudson, Eds), Raven Press, N. Y., 111-122.
13. LONGO, F. J. (1981). Develop. Biol. 84, 173-182. 14. POLLQCK, E. G. (1978). Amer. Zool. 18, 25-69.