Chemistry and material science at the cell surface
Texte intégral
Figure

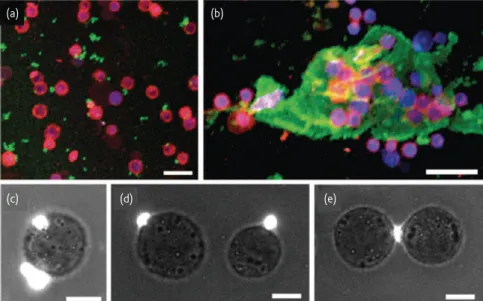
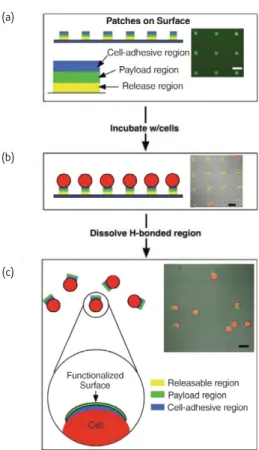
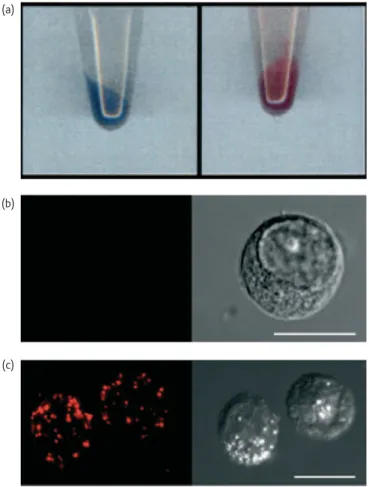

Documents relatifs
The ontophylogenesis theory considers the generation of a differentiated state as a constrained random process: randomness is provided by the stochastic dynamics of
The present study revealed that the present set-up based on ¯uorescent labelling of cells, long-term monitoring of cell shape and migration, using Confocal laser scanning
In vitro, human primary myoblasts have been shown to provide a very useful tool for the study of viral infection, since they can be differentiated into myotubes (Figure 1),
The Gene Expression Profile of Nodal T-Cell Lymphomas Identifies a Molecular Link between Angioimmunoblastic T-Cell Lymphoma (AITL) and Follicular Helper T Cells (TFH), and
4-10 This review will not cover the prolific recent findings in nematode effector functions but will instead focus on recent examples that illustrate the variety of plant
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des
“We take pretty seriously that idea of taking people on,” he says, “because more often than not, we’re making some kind of, you know, not forever marriage, but a
Therefore, even though activation of the Raf pathway is sufficient to decrease CD24 mRNA and protein, inhibition of oncogenic Raf/MEK signaling does not restore CD24 expression to
