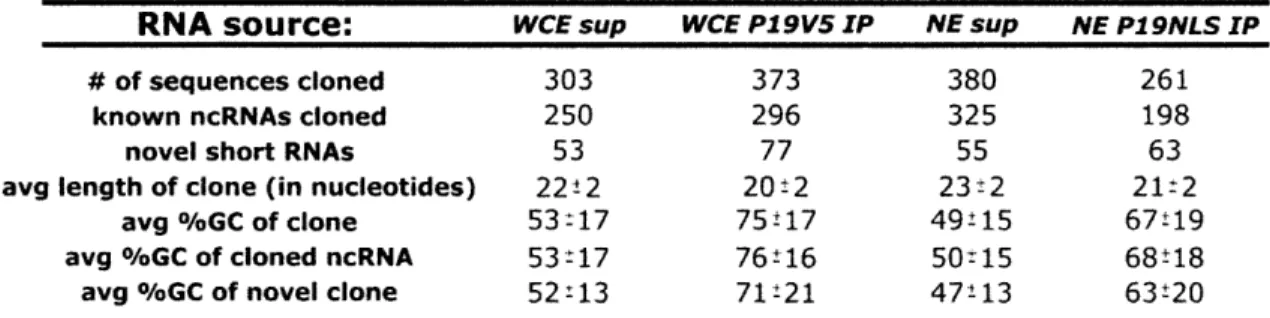
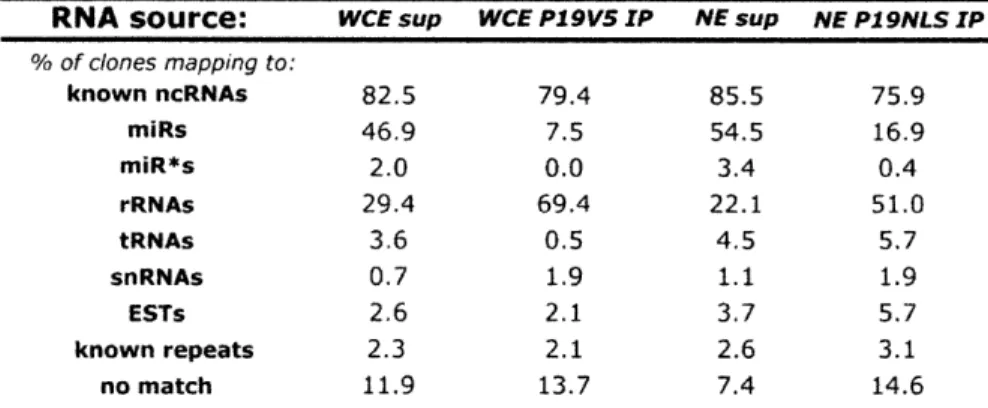
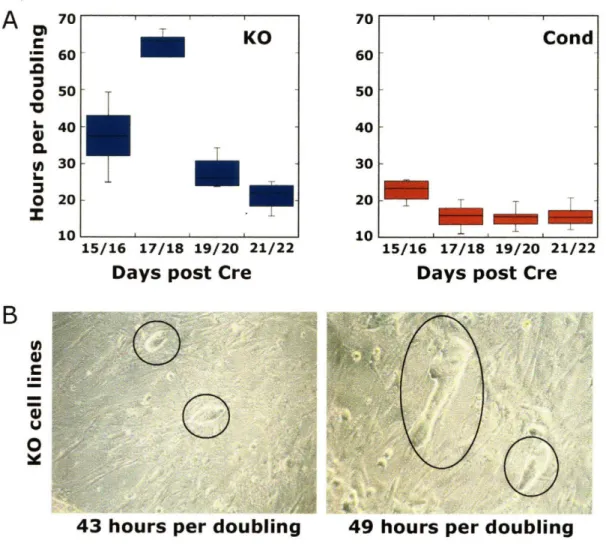
Dicer deletion and short RNA expression analysis in mouse embryonic stem cells
by
Joseph Mauro Calabrese
B.S. Chemistry, Biochemistry, and Molecular Biology (2001) University of Wisconsin-Madison
Submitted to the Department of Biology
In Partial Fulfillment of the Requirements for the Degree of Doctor in Philosophy in Biology
at the
Massachusetts Institute of Technology February 2008
C Massachusetts Institute of Technology All rights reserved.
Signature of Author:_
J
Department of Biology
December 18d, 2007 Certified by:C/'
Phillip A.
Sharp
Professor of Biology Thesis Supervisor Accepted by: Steven P. Bell Professor of BiologyChair, Biology Graduate Committee
MASSACHUSETT MImr1M
OF TEOHNOLOGY
FEB 1-2
2008
LIBRARIES
Dicer deletion and short RNA expression analysis in mouse embryonic stem cells By
Joseph Mauro Calabrese
Submitted to the Department of Biology in Partial Fulfillment of the Requirements for the Degree of Doctor in Philosophy in Biology.
ABSTRACT
RNA interference (RNAi) manages many aspects of eukaryotic gene expression through sequence-specific interactions with RNA. Short RNAs, 20-30 nucleotides in length, guide the various effector proteins of RNAi to silence fully or partially
complementary targets. The sequencing of endogenously expressed short RNA species coupled with genetic studies in various experimental organisms has revealed a role for RNAi in the silencing of protein-coding genes and repetitive elements in genomes. In mammals, it is unknown to what extent RNAi is involved in silencing processes other than the modulation of protein-coding gene expression, which is achieved through a class of short RNAs termed microRNAs (miRNAs).
The work in this thesis quantitatively describes the short RNAs expressed in mouse embryonic stem (ES) cells. ES cell lines are derived from the pre-implantation blastocyst and can be cultured in vitro for extended periods while still maintaining pluripotency. It was demonstrated that approximately 130,000 5' phosphorylated short RNA molecules are present in a single ES cell. 10% of these short RNAs represent non-random fragments of larger, abundant non-coding RNA species, and have no known function. Low abundance short RNAs were discovered that cluster bidirectionally around the transcription start sites of protein-coding genes. These RNAs associate with features
of active transcription, and may be evidence of widespread bidirectional initiation and pausing of RNA polymerase II in ES cells.
There are on the order of 300 different miRNA species expressed in ES cells, comprising 85% of the total pool of 130,000 5' phosphorylated short RNAs. Based on experiments correlating miRNA abundance to target repression, only about 30 of these miRNAs are expected to carry significant ES cell regulatory capacity. ES cells lacking all miRNAs do not significantly change their morphology or gene expression patterns, but do show a significant drop in growth rate compared to controls, suggesting that a major function of ES cell miRNAs may be to govern cell division. A detailed
comparison of short RNAs expressed in ES cells with and without the ribonuclease Dicer strongly suggests that miRNAs are the sole regulatory molecules that function through the RNAi pathway in ES cells. Considering previous work showing that repeating elements are frequently under Dicer-dependent repression, this observation raises the possibility that mammalian miRNAs may in certain contexts function to silence repeating genomic elements in addition to protein-coding genes.
ACKNOWLEDGEMENTS
To Phil, for training me to become a research scientist. I have learned so much in the lab it is impossible to recount the details in this space. Thanks for all of your help and
advice, and for providing me with so many opportunities to discover.
To my committee members, Dave Bartel and Rudolf Jaenisch for sage advice and expanding the way we thought about diverse problems.
To all of my co-workers in the Sharp lab over the years, so many of you have been teachers and role models, in addition to friends. Thanks for everything, especially the
Muddy Charles trips. Sharp lab rules.
To my friends who have made my life outside of lab exciting, thanks. Many highlights come to mind, including: the CCR retreats, Harvard Law parties, firing Ann and Keara, taquito bombs, the syrup race, grilled pecan sandies, flip cup, uneven pool tables, road
trips, good beadings, physical challenges on Boston Commons, Department-funded recruitment events at Pleasant Place, four 4th of Julys, and other barbecue-centered
events, too numerous to mention.
To Nicole, for all of your support and for providing so many pleasant distractions during times of stress.
To my parents, for all that you have done. Too few people in this world have had the opportunities and support that you have given me. Without your guidance,
Discovery consists of seeing what everyone else has seen and thinking what no one else has thought.
5
TABLE OF CONTENTS
A bstract ... ... 2
Chapter 1 RNA interference and the biology of mouse embryonic stem cells... 6
Animal miRNAs...8
miRNA biogenesis ... 8..
miRNA-mediated silencing mechanisms ... ... ... ... 11
m iRNA function ... 14
C. elegans antisense siRNAs... ... 17
RNAi-mediated transcriptional silencing in S. pombe... 20
RNAi-mediated viral and transcriptional silencing in plants ... 21
RNAi-mediated silencing of transposons and transgenes in C. elegans ... 23
RNAi-mediated transposon control in D.melanogaster ... 24
Mammalian RNAi and repetitive elements ... 26
M ouse embryonic stem cells ... ... 29
Chapter 2 Characterization of the short RNAs bound by the P 19 suppressor of RNA silencing in mouse ES cells ... 48
Chapter 3 RNA sequence analysis defines Dicer's role in mouse ES cells...89
Chapter 3 Appendix... ... 129
Chapter 4 Short RNAs in the sense and anti-sense orientation from transcription initiation sites in m ouse ES cells... ... 147
Chapter 5 Examining miRNA function in mouse ES cells ... 174
Chapter 1
RNA interference and the biology of mouse embryonic
stem cells
Introduction
To manage their gene expression programs, organisms employ many distinct mechanisms. In one set of regulatory mechanisms, termed RNA interference (RNAi), short RNAs 20-30 nucleotides (nt) in length, guide multi-protein complexes to suppress functions of complementary nucleic acid targets. Present in many single-celled, and likely all multi-celled eukaryotes, the processes of RNAi regulate protein-coding gene expression, initiate and maintain transcriptional silencing of specific genomic loci, and maintain genomic integrity and immunity via the silencing transposable elements. In Chapter 1, the various mechanisms of RNAi-mediated silencing are described, focusing heavily on studies conducted in mammals, though also touching on aspects of RNAi in many experimental organisms. Additionally, a basic introduction to the biology of mouse embryonic stem (ES) cells is included to provide appropriate background to the research described in this thesis.
RNAi in the control of protein-coding gene expression in animals
Animal microRNAs
RNAi is a master regulator of protein-coding gene expression, predominantly through a class of -22 nt long non-coding RNA genes termed microRNAs (miRNAs). miRNAs are sequence-specific guide molecules for protein complexes that prevent productive translation and destabilize mRNAs. miRNAs appear to be ubiquitously expressed in all multi-cellular eukaryotes, and have recently been identified in the unicellular eukaryote, Chlamydomonas reinhardtii (Molnar et al. 2007; Zhao et al. 2007a). Their roles are diverse, and in many cases, essential. From a growing set of genetic, biochemical, and computational analyses, it appears that many miRNAs control cell-fate specification and have pleiotropic effects on cellular environments, similar to cell-type-specific transcription factors (Kloosterman and Plasterk 2006). miRNAs have critical regulatory roles in plants as well as animals; however, significant differences exist between plant and animal miRNA biosynthesis and function. In the text below, only animal miRNAs are discussed.
miRNA biogenesis
miRNAs are transcribed by RNA Pol II as long primary transcripts, termed pri-miRNAs, that are capped, poly-adenylated, and frequently poly-cistronic (Cai et al. 2004; Lee et al. 2004; Rodriguez et al. 2004). Many miRNAs are located in defined intergenic transcriptional units (Saini et al. 2007), others are located in introns and likely co-expressed with host genes from single promoters (Baskerville and Bartel 2005). Pri-miRNAs are processed in the nucleus by the Drosha-DGCR8 heterodimer to generate
-70 nt long pre-miRNA hairpins with characteristic 5' phosphates and 3' 2 nt overhangs (Lee et al. 2003; Zeng et al. 2005; Han et al. 2006). Pre-miRNAs are then exported into the cytoplasm by Exportin-5 and Ran-GTP (Ying et al. 2003).
After nuclear export, the pre-miRNA hairpin is processed by the cytoplasmic enzyme Dicer to generate a ~22 base pair RNA duplex consisting of the mature miRNA paired to its complement, termed the miRNA* (Grishok et al. 2001; Hutvagner et al. 2001; Ketting et al. 2001). This duplex is likely short-lived, as miRNA* levels are up to
100 fold lower than levels of corresponding miRNAs (Ruby et al. 2006). Overexpression of an RNA-duplex binding protein in mammalian cells fails to capture miRNA-miRNA* duplexes, consistent with their proposed short life span and suggesting that these
duplexes are bound by protein components in the cytoplasm (described in Chapter 2). After Dicer processing, the mature single-stranded miRNA is then displaced from the miRNA* and incorporated into an active silencing complex.
The Argonaute proteins bind miRNAs in the core of the multi-subunit RNA-induced silencing complex (RISC), the protein complex that mediates RNAi-based silencing (Liu et al. 2004). Mammals have eight Argonaute paralogues, divided equally between the Ago and Piwi subfamilies (Carmell et al. 2002). Ago subfamily members are thought to strictly associate with miRNAs, while at least two Piwi subfamily members associate with a separate class of RNAs, termed piRNAs (Liu et al. 2004; O'Donnell and Boeke 2007). Of the 4 Ago proteins, only Ago2 is capable of cleaving target transcripts that are perfectly complementary to bound miRNAs (Liu et al. 2004). Transcript cleavage by Ago2 is multi-turnover and occurs on the target RNA directly across from the 10h nucleotide measuring from the 5' end of the miRNA (Hutvagner and
Zamore 2002; Martinez and Tuschl 2004). Ago2 does not explicitly depend on this cleavage activity to function, as expression of a cleavage-deficient Ago2 mutant protein is able to fully rescue the phenotypic defects of a mouse Ago2 hematopoietic knockout (Tang et al. 2007). The other Ago proteins lack significant cleavage activity and likely function mainly to prevent translation of target mRNAs (Pillai et al. 2004).
Many proteins associate with the Argonautes either as RISC loading or accessory factors. In HEK 293T cells, the double-stranded RNA (dsRNA) binding protein TRBP associates with Dicer and Ago2 and is likely required for proper loading of miRNAs into the RISC (Chendrimada et al. 2005). Biochemical studies mainly conducted in cells from
Homo sapiens and Drosophila melanogaster have shown many other proteins associate
with RISC as accessory factors, including: the fragile-X-mental-retardation protein (FMRP), tudor staphylococcal nuclease (TSN), the vasa intronic gene (VIG), Mov 10, elF6, and Gemin3 and Gemin4 (Meister et al. 2005; Sontheimer 2005; Chendrimada et al. 2007). The function of many of these proteins in miRNA-mediated silencing, and
whether they consistently associate with Ago and the RISC in multiple cell types, remains unclear.
Loading of single-strand mammalian miRNAs into the RISC is thought to depend on the difference in thermodynamic end stabilities between the two ends of the
miRNA/miRNA* duplex. Analysis of functional short-interfering RNAs (siRNAs) and both vertebrate and invertebrate miRNAs has shown that the short RNA whose 5' terminus is located at the end of the duplex that is least thermodynamically stable is preferentially incorporated into the RISC (Khvorova et al. 2003; Schwarz et al. 2003). This difference in thermodynamic stability can in many cases accurately predict which
strand of the pre-miRNA hairpin will be the miRNA and which will be preferentially degraded as the miRNA*; however, exceptions exist where differences in thermodynamic stability alone are insufficient to predict miRNA duplex strand choice (Khvorova et al. 2003; Schwarz et al. 2003).
In addition to miRNA duplex end stabilities, base pairing between the body of the miRNA and miRNA* can affect how miRNAs are loaded into different RISCs. In D.
melanogaster, miRNAs duplexes with perfect complementarity across from the site of
Ago2 cleavage are preferentially incorporated into Ago2-containing RISCs, while those with bulges in the would-be cleavage region-positions 9 through 11 measuring from the miRNA 5' end-are preferentially incorporated into Agol-containing RISCs (Forstemann et al. 2007; Tomari et al. 2007). In mammals, this differential incorporation of miRNAs into RISCs does not appear to occur, as immunoprecipitation of different Argonautes followed by miRNA microarray analysis shows that Ago 1, 2, and 3 bind all miRNAs equally well (Liu et al. 2004).
Dynamic changes in miRNA levels have been observed along developmental axes and changes in physiological state (Cheng et al. 2007; Neilson et al. 2007; Xu et al. 2007), suggesting the existence of active processes for the regulation of mature miRNA levels. Nevertheless, some mature miRNAs appear to be very stable in non-dividing cells (Song et al. 2003). Also, levels of mature miRNAs are often uncoupled from levels of pre- and pri-miRNAs, suggesting that miRNA processing itself is a regulated process
(Obernosterer et al. 2006; Thomson et al. 2006).
The mechanisms of miRNA-mediated gene silencing processes appear to be diverse. The founding miRNA, Caenorhabditis elegans lin-4, was observed to prevent translation of its target mRNA, lin-14, through an interaction that required partially complementary sequences in lin-14's 3' untranslated region (UTR). This translational repression did not significantly change lin-14 mRNA levels or the location of lin-14 mRNA in a polysome sedimentation profile (Lee et al. 1993; Wightman et al. 1993; Olsen and Ambros 1999). A large body of subsequent work shows that translational repression of mRNAs via partially complementary sequence interaction is the
predominant mechanism of miRNA-mediated gene silencing in animals (Bartel 2004). However, at least one animal miRNA, miR-196, functions by cleaving perfectly
complementary target transcripts (Yekta et al. 2004). Moreover, it has been observed that many miRNAs can destabilize target mRNAs, likely by causing relocation of mRNAs to cytoplasmic processing bodies (P-bodies) (Bagga et al. 2005; Lim et al. 2005).
Sequence information in 3' UTRs predominantly dictates the type and extent of miRNA-mediated mRNA repression. Comparative genomics studies have shown that miRNA target sites on mRNAs are most conserved over bases 2-8 measuring from the 5' end of the miRNA, termed the "seed" region of the miRNA (Lewis et al. 2003; Lewis et al. 2005). Experiments testing the repressive capability of both artificial and natural miRNAs are in agreement with this, having demonstrated that perfect base pairing between the 5' end of the miRNA and the 3' UTR is a strong determinant of repression
(Doench and Sharp 2004; Brennecke et al. 2005). Additionally, these experiments showed that extensive pairing to the 3' end of the miRNA can compensate for weak 5' pairing, and that miRNA sites in close proximity synergize with each other,
demonstrating that a single UTR may be subject to regulation from many miRNAs (Doench et al. 2003; Doench and Sharp 2004; Brennecke et al. 2005). More recently, it has been shown that target site accessibility, local A/U content, target-site-proximal conservation, and location of the target site relative to the stop codon, are all additional determinants of miRNA-mediated repression (Grimson et al. 2007; Kertesz et al. 2007; Nielsen et al. 2007).
Though it is accepted that the majority of animal miRNAs function by preventing productive translation of their target mRNAs, the apparent mechanisms of translational inhibition by miRNAs vary depending on the experimental system used by researchers. Studies using various in vitro cell extracts or in vitro transcribed mRNAs have shown miRNAs inhibit translational initiation in a manner dependent on a 7-methyl-guanine (m7G) cap structure and a poly-A tail (Humphreys et al. 2005; Pillai et al. 2005; Wang et al. 2006a; Mathonnet et al. 2007; Wakiyama et al. 2007). Argonaute proteins have a m7G cap binding domain that is similar to the that of the cap binding protein eIF4E, and this domain is required for Ago-mediated translational repression of mRNAs (Kiriakidou et al. 2007). Furthermore, addition of recombinant eIF4E to extracts interferes with
miRNA-mediated translational inhibition (Mathonnet et al. 2007). Together, these studies support a model by which miRNA-guided RISCs bind m7G cap structures to prevent translational initiation.
Apparently at odds with these findings are a number of studies analyzing cells that suggest miRNAs inhibit translation at a step post-initiation (Olsen and Ambros 1999;
Seggerson et al. 2002; Maroney et al. 2006; Nottrott et al. 2006; Petersen et al. 2006). In these works, miRNAs and mRNAs actively repressed by miRNAs co-sediment with
polyribosomes in sucrose gradients, and this co-sedimentation can be disrupted by
puromycin, suggesting it depends on actively translating ribosomes (Maroney et al. 2006; Nottrott et al. 2006; Petersen et al. 2006). Further, miRNAs inhibited translation of a reporter gene driven by the cricket paralysis virus IRES, which allows loading of elongation-competent 80S ribosomes on mRNAs without the requirement for canonical initiation factors and initiator tRNAs, again suggesting that miRNAs repress translation post-initiation (Petersen et al. 2006). One potential explanation for these discrepancies could be that miRNAs may inhibit translation both pre- and post-initiation, but certain experimental conditions, such as those that utilize in vitro transcribed mRNAs, are differentially sensitive to these two modes of inhibition.
miRNAs can also induce mRNA destabilization by targeting mRNAs for deadenylation, and potentially decapping. Zebrafish miR-430 promotes clearance of maternal mRNAs at the onset of zygotic transcription via deadenylation, and the miRNA
let-7 promotes translation-independent deadenylation of a reporter mRNA in vitro
(Giraldez et al. 2006; Wakiyama et al. 2007). Separate experiments show that Argonaute proteins associate with P-bodies and decapping enzyme in tissue culture cells, indirectly linking miRNA-mediated repression to decapping (Jakymiw et al. 2005; Liu et al. 2005; Pillai et al. 2005; Sen and Blau 2005). Further, decay products have been detected of miRNA-targeted mRNAs that are consistent with the 5' to 3' exonucleolytic degradation mediated by Xrnlp, the nuclease that destroys uncapped mRNAs (Bagga et al. 2005).
The current miRNA database has annotations for 533 human and 442 mouse miRNAs (Griffiths-Jones 2004). Given the large number and apparent ubiquitous expression of miRNAs in animals, the potential for miRNA-mediated gene regulation is large. The founding miRNAs were identified in forward genetic screens via their
phenotypic influence on genetic pathways (Chalfie et al. 1981; Reinhart et al. 2000); however, the small amount of sequence complementarity needed for miRNA-mediated repression suggests that most miRNAs affect many cellular pathways rather than one specifically.
miRNAs tune translation from expressed mRNAs to define protein output of targeted genes. In one specific example, reduction of atrophin levels by Drosophila miR-8 is required for normal central nervous system function. Importantly, further reduction or overexpression of atrophin in otherwise wild-type flies result in a mutant phenotype, indicating that miR-8 reduces atrophin expression to a level appropriate for normal function (Karres et al. 2007). miR- 150 expression in differentiating B cells represents another example of miRNA-mediated tuning of protein expression (Xiao et al. 2007). miR-150 targets the transcription factor c-Myb during lymphocyte development. Similar to the situation described for atrophin and miR-8 in Drosophila, the relief of miR- 150-mediated c-Myb repression or a reduction of c-Myb protein levels both result in the impairment of B cell development (Xiao et al. 2007).
Many miRNAs are reciprocally expressed with target mRNAs, suggesting that in certain cases, miRNAs function as master regulators of cell-type specific transcriptional and translational output (Farh et al. 2005; Stark et al. 2005). Genes that are highly expressed in specific tissues have evolved to avoid targeting by abundant tissue-specific
miRNAs, whereas genes that are conserved targets of tissue specific miRNAs are frequently expressed at low levels, or in tissues adjacent to the tissue specific miRNA-such that gene expression boundaries and cell identity appear to be maintained by
miRNA expression (Farh et al. 2005; Stark et al. 2005). Additionally, genes that need to be ubiquitously expressed, such as ribosomal protein genes, tend to have short UTRs that avoid miRNA targeting completely (Stark et al. 2005). Consistent with a role for
miRNAs in the restriction of tissue identity, miRNA levels are generally down-regulated in tumors, and impaired miRNA processing enhances tumorigenesis, a process in which diverse collections of rapidly evolving cells need to adopt multiple cellular identities (Lu et al. 2005; Kumar et al. 2007).
Genetic knockouts of specific mouse miRNAs reveal a striking intolerance for loss of tissue-specific miRNA expression. The most poignant example of this thus far is knockout of miR- 1-2, which is expressed specifically in muscle cells. Approximately 50% of miR-1-2 knockout mice die from severe cardiac dysfunction at or before weaning, indicating a critical role for miR-1-2 in the heart (Zhao et al. 2007b). Also, deletions of the lymphoid specific miRNAs miR-150 and -155 result in severe defects in B-cell and T-cell differentiation, respectively (Rodriguez et al. 2007; Thai et al. 2007; Xiao et al. 2007).
Deletion of Dicer from mouse tissues results in catastrophic abnormalities in all cases examined, again suggesting that miRNA function is critical in many tissues; however, because Dicer is required for the biogenesis of several other regulatory RNAs in non-mammalian organisms, the phenotypic consequences of Dicer loss may not be solely due to loss of miRNA expression and must be carefully interpreted. Dicer
knockout mice die at the earliest stage examined, E7.5, and oocyte-specific Dicer
deletion results in arrest at oocyte meiosis I, showing the necessity of Dicer activity at the earliest stages in mouse development (Bernstein et al. 2003; Murchison et al. 2007; Tang et al. 2007). Tissue-specific deletions of Dicer in the limb, lung, immune system, heart, and epidermis all show catastrophic mutant phenotypes, consistent with a requirement for Dicer function in these tissues (Harfe et al. 2005; Andl et al. 2006; Harris et al. 2006; Yi et al. 2006; Zhao et al. 2007b).
To conclude, there is a role for miRNA-mediated gene regulation in a large number of biological processes. Not discussed here are documented roles for miRNAs in a range of biology, including apoptosis, metabolism, cell division, metastasis, local translation at synapses, and management of circadian rhythms (Kloosterman and Plasterk 2006; Cheng et al. 2007; Wu et al. 2007; Xu et al. 2007). Additionally, it is possible that miRNAs have a generalized role in regulating gene expression during stress (Leung and Sharp 2007). Recent observations show that Ago2 and the RISC component FXR1 are curiously required for the up-regulation of TNFa protein in human cells after serum starvation (Vasudevan and Steitz 2007); whether or not this up-regulation is miRNA-dependent is currently unclear.
C.elegans antisense siRNAs
In the nematode C. elegans, the expression of short RNAs antisense to protein-coding mRNAs is thought to modulate protein-protein-coding gene expression in a manner separate from miRNAs, likely by guiding direct mRNA cleavage (Ambros et al. 2003). Several proteins are implicated in the biogenesis of these endogenous siRNAs, including Dicer, an RNA-dependent RNA polymerase (RdRP), an RNA helicase, an RNAse D
homologue, a nucleotidyltransferase, and the conserved RNA phosphatase Pir- 1
(Duchaine et al. 2006; Lee et al. 2006a; Sijen et al. 2007). The mechanistic details of C.
elegans siRNA biogenesis remain unclear. Potentially, target mRNAs serve as templates
for an RdRP to generate double-stranded RNA species with 5' tri- or di-phosphates. Presumably, these phosphates need to be removed before Dicer processing, as pir-1 phosphatase mutants accumulate long RNAs anti-sense to target transcripts (Duchaine et al. 2006). Dicer processing then likely generates primary siRNAs that are low in
abundance and serve as guides to initiate a second round of RdRP synthesis, this time resulting in abundant short 21-27 nt siRNAs that likely function to silence
complementary mRNAs (Ruby et al. 2006; Sijen et al. 2007).
C. elegans siRNAs have 5' tri- or di-phosphates, different from the 5'
mono-phosphates of miRNAs. Unlike miRNAs, most C. elegans siRNAs do not serve as substrates for T4 RNA ligase in vitro, which requires a 5' mono-phosphate; however, they do show a shift in mobility after treatment with alkaline phosphatase, indicating the presence of at least one terminal 5' phosphate (Pak and Fire 2007; Sijen et al. 2007). C.
elegans siRNAs also serve as substrates for in vitro capping reactions that require 5'
tri-or di-phosphates, and exhibit gel mobility patterns that mimic the mobility of synthetic 5' tri- and phosphorylated RNAs, strongly suggesting they are marked with 5' tri- or di-phosphates (Ruby et al. 2006; Pak and Fire 2007; Sijen et al. 2007).
The 5' end modification of C. elegans siRNAs is noteworthy because it greatly reduces endogenous siRNA sequencing frequency in short cDNA libraries that have been prepared by selecting for the canonical 5' and 3' end modifications of animal miRNAs, 5' monophosphates and 3' hydroxyls. Ruby and colleagues, selecting for short RNAs with
5' monophosphates and 3' hydroxyls, found miRNAs to be 100-fold more abundant than anti-sense siRNAs in mixed-stage C. elegans (Ruby et al. 2006). In contrast, using a cDNA library preparation method that was independent of 5' modification, Ambros and colleagues found miRNAs and siRNAs to be approximately equal in abundance (Ambros et al. 2003). It is currently unclear whether other organisms express endogenous short RNAs with similarly modified 5' termini; however, these studies set the precedent for short RNA species eluding discovery because of end modifications incompatible with cDNA library preparation methods.
RNAi in the control of heterochromatin and transposable elements
RNAi has a conserved role in the silencing of transposable elements and the establishment of heterochromatin at repetitive loci in eukaryotic genomes. These RNAi-based silencing processes are diverse, and understood in varying detail, discussed in the text below. There are many cases in which RNAi prevents the replication of exogenous RNA viruses on a post-transcriptional level. Also, at least in plants, RNAi
transcriptionally silences exogenous viruses that have integrated into the genome. In a related set of silencing mechanisms, RNAi prevents the spread of endogenous
transposable elements on both transcriptional and post-transcriptional levels. These types of endogenous transposable elements represent a large portion of many eukaryotic
genomes, and usually express one or several proteins that function in concert with cellular machinery to replicate. In certain cases, formation of heterochromatin around these elements is a direct consequence of the protective role of RNAi. In many cases,
repeats are silenced not as an act of genomic defense but instead as a means to coordinately regulate nuclear domains for maintenance of genome structure or in
response to developmental cues, suggesting exaptation of this defense pathway. Notably, despite extensive conservation of RNAi components in eukaryotes, it is unclear to what extent RNAi mediates the silencing of repetitive elements and the formation of
heterochromatin in mammals.
RNAi-mediated transcriptional silencing in Schizosaccharomyces pombe
RNAi-mediated transcriptional silencing is best understood in S. pombe, which has only one member from each of three major gene families involved in RNAi.
Targeted deletion of the sole Dicer (dcrl), Argonaute (ago 1), or RdRP (rdp 1) de-silences centromeric repeats and results in defects in mitotic chromosome segregation and
telomeric clustering, indicating a role for RNAi-mediated transcriptional silencing in genomic integrity and high-order nuclear structure (Volpe et al. 2002; Hall et al. 2003; Sugiyama et al. 2005). RNAi is also needed for heterochromatin establishment at the repetitive mating-type locus, which is a 20kb region harboring a copy of the centromeric repeat cenH flanked by inverted repeats that serve as boundary elements to
heterochromatin formation (Hall et al. 2002; Jia et al. 2004).
A detailed mechanistic model has emerged for RNAi-mediated establishment and maintenance of heterochromatin in S. pombe (Colmenares et al. 2007). Nascent
transcripts generated by RNA Polymerase II (RNA Pol II) at heterochromatic loci are bound by complementary short RNAs carried in the RNA-induced-transcriptional-silencing, or RITS, complex (Buhler et al. 2006; Irvine et al. 2006). The trimeric RITS
complex, composed of Ago 1 bound to a heterochromatic siRNA, the chromodomain-containing protein Chpl, and a protein of unknown function, Tas3 (Verdel et al. 2004), then recruits an RdRP-containing complex (the RDRC) to the heterochromatic locus in a manner that requires both the catalytic cleavage activity of Agol and the histone3-lysine-9 (H3Khistone3-lysine-9) binding activity of Chpl (Motamedi et al. 2004; Noma et al. 2004; Irvine et al. 2006). Heterochromatin formation and siRNA production both require the H3K9
methyltransferase Clr4, highlighting the importance of Chp l's interaction with chromatin (Noma et al. 2004). Once tethered to the nascent transcript, the RDRC creates a double-stranded RNA that is processed by Dicer to generate more heterochromatic siRNAs and start the cycle anew (Colmenares et al. 2007). Interestingly, a centromeric repeat exogenously introduced into euchromatin is sufficient to induce RNAi-mediated heterochromatin formation, suggesting either a sequence specificity to the RNAi-mediated induction of heterochromatin, or that siRNA-loaded RITS can act in trans to silence dispersed repeats (Hall et al. 2002).
RNAi-mediated viral and transcriptional silencing in plants
The RNAi pathway has a significant role in plant antiviral immunity. Many plant viruses encode single- or double-stranded RNA genomes that are recognized by RNAi machinery as deleterious and serve as substrates for the generation and amplification of targeting siRNAs. In this process, the viral genome is converted into double-stranded
RNA via an RdRP and subsequently converted into siRNAs by a plant Dicer, after which the viral siRNAs are incorporated into a viral-targeting RISC that likely contains plant Agol at its core (Xie and Guo 2006; Zhang et al. 2006). An interesting aspect of this
silencing mechanism is that viral siRNAs are not only present in the infected cells, but spread throughout the plant and protect distal portions of the plant from subsequent viral infections (Hamilton et al. 2002). Accordingly, plants with mutations in various Dicers (dcl2 and dcl4) and RdRPs (rdr I and rdr6) are highly susceptible to local and systemic viral infection (Mourrain et al. 2000; Deleris et al. 2006). Emphasizing its protective importance, potentially all plant RNA viruses have evolved to express proteins that are potent inhibitors of the plant RNAi antiviral response (Voinnet 2005).
The P19 protein from Tombus family of viruses is the best-characterized viral inhibitor of RNAi (Scholthof 2006). P 19 functions by binding and sequestering viral-targeting siRNAs that in their free form would be incorporated into a plant RISC. X-ray crystallographic and biochemical studies show that head-to-head dimers of P19 bind siRNA duplexes with low nanomolar affinity (Vargason et al. 2003; Ye et al. 2003; Lakatos et al. 2004). Many viral inhibitors of RNAi are proposed to function similarly (Voinnet 2005); still others inhibit different steps of the RNAi pathway, as demonstrated by the 2b protein from the Cucumber mosaic virus, which inhibits plant AGO 1 (Zhang et
al. 2006).
RNAi also plays a significant role in the establishment of heterochromatin surrounding repetitive elements in plant genomes. One branch of this pathway mediates the formation of heterochromatin around invading viruses and exogenously introduced transgenes that have integrated into plant genomes, and has genetic requirements similar to those described above for post-transcriptional silencing of viruses, namely dcl2, rdrl, and rdr6 (Dalmay et al. 2000; Fagard et al. 2000; Mourrain et al. 2000). The other branch serves to silence endogenous repetitive elements, and requires separate RNAi paralogues
to function. Certain classes of transposons and ribosomal DNA loci are de-silenced in mutant strains of Dicer (dcl3), Argonaute (ago4), and RdRP (rdr2). This de-silencing is accompanied by disappearance of short RNAs corresponding to the repeats, and a reduction in H3K9 and DNA methylation at the repetitive loci (Lippman et al. 2003; Zilberman et al. 2003; Chan et al. 2004; Xie et al. 2004). Synthesis and subsequent loading of heterochromatic siRNAs into silencing complexes also depends on RNA Polymerase IV, a DNA-dependent RNA polymerase that localizes with Ago4 in nuclear Cajal bodies (Onodera et al. 2005; Li et al. 2006; Zhang et al. 2007).
RNAi-mediated silencing of transposons and transgenes in C. elegans RNAi is required for the suppression of transposon replication and the transcriptional silencing of transgene arrays in the C. elegans germline and soma. Genetic screens first uncovered a role for RNAi in these processes, showing that the RNAseD homologue mut-7 and the Argonaute-like protein ppw-2 are required for germline suppression of transposition in C. elegans (Ketting et al. 1999; Vastenhouw et al. 2003). Subsequent work showed that many genes required for the suppression of transposition are also required for co-suppression, the process by which high-copy transgenes can induce the silencing of related endogenous genes in trans (Ketting and Plasterk 2000). Candidate-based RNAi screens uncovered additional genes required for germline suppression, including many chromatin modifiers, suggesting that co-suppression is at least partly a transcriptional silencing process (Robert et al. 2005). Transcriptional silencing of transgene arrays in the C. elegans soma requires a different
set of RNAi paralogues, including Dicer (dcr-1), the double-stranded RNA binding protein rde-4, the Argonaute protein rde-1, and the RdRP rrf-1 (Grishok et al. 2005).
RNAi-mediated transcriptional silencing, transposon control, and viral defense in D.
melanogaster
Transcriptional and post-transcriptonal control of endogenous repeats by RNAi
D. melanogaster RNAi pathway components are required for the transcriptional
and post-transcriptional control of repetitive elements. Post-transcriptional silencing of the tandemly repeated Stellate genes and other classes of retrotransposons requires at least one of two genes from the Piwi-subfamily of Argonaute proteins, Aubergine and Piwi, and two DExH-box helicases also involved in RNAi, Spindle-E and Armitage (Aravin et al. 2001; Vagin et al. 2006). The silencing of transgene arrays on the transcriptional and post-transcriptional levels has similar genetic requirements (Pal-Bhadra et al. 2002; Pal-(Pal-Bhadra et al. 2004). Moreover, piwi, aubergine, and spindle-E mutant flies show genome wide defects in heterochromatin, including reduction in H3K9 methylation levels and delocalization of the heterochromatin binding protein, HP 1 (Pal-Bhadra et al. 2004). Curiously, a repetitive locus associated with the telomere on the right arm of D. melanogaster chromosome 3, the 3R-TAS locus, is constitutively
euchromatic; however, in piwi mutants 3R-TAS becomes heterochromatinized despite a genome-wide decrease in H3K9 marks, suggesting that Piwi may also inhibit the spread of heterochromatin in addition to nucleating its formation (Yin and Lin 2007).
RNAi components are also required for the nuclear clustering of Polycomb genes in D. melanogaster (Grimaud et al. 2006). Using transgenic flies carrying multiple copies of the Polycomb response element Fab-7 integrated at different genomic locations, Grimaud and colleagues showed that Piwi, Ago 1, and Dicer-2 (Dcr-2) frequently co-localize with nuclear clusters of Fab-7 (Grimaud et al. 2006). Further, piwi, agol, and
dcr2 mutant flies still maintained silencing but could no longer organize Fab-7 repeats
into punctate nuclear foci, indicating a role for RNAi in the high-order nuclear organization but not silencing of Polycomb response elements (Grimaud et al. 2006).
Sequence analysis of short RNAs associated with Piwi subfamily proteins (or piRNAs, for piwi-interacting RNAs) reveals several interesting characteristics (Brennecke et al. 2007; Gunawardane et al. 2007). D. melanogaster piRNAs are on average longer than miRNAs, 23-27 compared to -22 nt. Though the majority are complementary to highly repetitive elements, analysis of those with unique genomic locations reveals that piRNAs are generated from large, discreetly located clusters that harbor diverse classes of transposons (Brennecke et al. 2007). Strikingly, piRNAs associated with different Piwi subfamily members show characteristic first and tenth nucleotide biases, such that the large majority of Piwi- and Aubergine-associated RNAs begin with 'U' residues, while Ago3-associated RNAs frequently contain 'A' residues at their tenth nucleotide (Brennecke et al. 2007; Gunawardane et al. 2007). Even more surprising is the staggered overlap of piRNAs associated with different Piwi-subfamily members. It was observed that a large number of Ago3 piRNAs were complementary to the first 10 bases of piRNAs associated with Aubergine and Piwi (Brennecke et al. 2007; Gunawardane et al. 2007). Together with the observations that (1) the first and tenth
nucleotides of Aubergine/Piwi- and Ago3-associated RNAs are complementary ('U' and 'A', respectively), (2) Piwi proteins are capable of cleaving complementary transcripts, and (3) piRNAs appear to exist in the absence of Dicer, the data suggest a model in which piRNAs are predominantly generated by other piRNA-containing protein complexes rather than Dicer cleavage of long, double-stranded RNA (Vagin et al. 2006; Brennecke et al. 2007; Gunawardane et al. 2007). Considering this model, one major puzzle is how this putative piRNA-induced biosynthetic loop is initiated.
RNAi-mediated antiviral immunity in D. melanogaster
D. melanogaster antiviral immunity depends on the RNAi pathway, analogous to the antiviral role of RNAi in plants. Mutations in Dicer-2, the Dicer-2 binding protein R2D2, or Ago2, render flies hypersusceptible to infection and lethality by various exogenous viruses (Galiana-Arnoux et al. 2006; van Rij et al. 2006; Wang et al. 2006b). At least 2 Drosophila viruses encode proteins, necessary for successful infection, that function by potently suppressing Dicer-2/R2D2 processing of double-stranded RNA, further evidence that D. melanogaster RNAi participates in antiviral immunity (Li et al. 2002; Galiana-Arnoux et al. 2006; van Rij et al. 2006).
Mammalian RNAi and repetitive elements
As described in the preceding sections of Chapter 1, repetitive regions of many eukaryotic genomes are frequently maintained in a silent state via the RNAi pathway. Though mammalian repeating elements are frequently associated with heterochromatin (Thurman et al. 2007), the extent that mammalian RNAi is involved in
heterochromatization of repetitive elements is currently unclear. Mammals do encode Piwi subfamily proteins that share characteristics with their Drosophila homologues, potentially indicating the existence of a germline RNAi pathway to repress the expression of mammalian repetitive elements. Also, studies examining aspects of early mouse development potentially implicate the RNAi pathway in the silencing of mammalian repeats outside of the germline.
Mammalian piRNAs
Like their Drosophila homologues, mammalian Piwi subfamily proteins associate with germ cell-specific RNAs termed piRNAs. These RNAs are 29-30 nt long and a large majority have 5' 'U' residues (Aravin et al. 2006; Girard et al. 2006; Lau et al. 2006). Unlike Drosophila piRNAs, mammalian piRNAs are generally not repetitive and frequently map uniquely to the genome (Aravin et al. 2006; Girard et al. 2006; Lau et al. 2006). piRNAs are produced from genomic clusters spanning approximately 20-100 kb, and exhibit a striking strand bias within these clusters, such that sequences associated with a particular Piwi protein almost never overlap in polarity (Aravin et al. 2006; Girard et al. 2006; Lau et al. 2006). This polarity bias is consistent with a mammalian piRNA biosynthetic pathway similar to that proposed in Drosophila, where different Piwi paralogues associate with partially complementary sequences and appear to
synergistically synthesize piRNAs (Brennecke et al. 2007; Gunawardane et al. 2007). piRNA sequences are not conserved between mouse, rat, and human; however, syntenic genomic regions give rise to piRNA clusters in these three organisms, suggesting that
genomic location rather than sequence may be important for piRNA function (Aravin et al. 2006; Girard et al. 2006; Lau et al. 2006).
Ablation in mice shows a role for Piwi proteins in spermatogenesis and implicates them in the germline silencing of repetitive elements. There are 4 Piwi proteins in mice: Miwi, Miwi2, Mili, and PiwiL3. PiwiL3 has not been studied in any context. Mice lacking Miwi, Mili, or Mili2 are male-specific infertile and show various defects in spermatogenesis (Deng and Lin 2002; Kuramochi-Miyagawa et al. 2004; Carmell et al. 2007). Additionally, Mili and Miwi2 null mice show reduced DNA methylation at repetitive elements, implicating these genes in a germline silencing pathway that
methylates repetitive elements (Aravin et al. 2007; Carmell et al. 2007). While consistent with Piwi function in Drosophila, the proposed role of Miwi2 and Mili in the silencing of repeats is at odds with sequence data showing that Mili associates predominantly with short RNAs that are not repetitive. It would seem possible, however, that the minority of Mili-associated piRNAs that are repetitive could guide silencing in trans. Alternatively, the observed DNA methylation defects in the germlines of Mili and Miwi2 null mice may
be an indirect consequence of Piwi protein loss.
Proposed roles for Dicer in the silencing of endogenous repeats
Studies of cells deleted or hypomorphic for Dicer indicate a potential role for mammalian RNAi in endogenous repeat silencing outside of the germline. DNA derived from the long interspersed nuclear element, or LINE, makes up approximately 20% of the mouse and human genome (Lander et al. 2001; Waterston et al. 2002). Full-length LINE repeats are -6kb in length and encode a reverse-transcriptase and chaperone protein that
replicate the LINE genome after it has been transcribed by cellular RNA polymerase II. Human cells treated with siRNA to reduce Dicer levels show a mild increase in frequency of LINE retrotransposition, suggesting that RNAi may repress LINE replication (Yang and Kazazian 2006). Knockdown of Dicer via RNAi in one-cell mouse embryos results in a -50% increase in steady-state levels of different classes of endogenous long terminal repeat (LTR) retrotransposons, which have a different genome structure but replicate via similar mechanisms as LINE repeats (Svoboda et al. 2004). LTR elements are also abundant repeats, representing approximately 10% of the human and mouse genomes (Lander et al. 2001; Waterston et al. 2002). Similar to observations from early mouse
embryos, Dicer knockout mouse oocytes accumulate retrotransposon RNA, and display elevated levels of transcripts containing specific classes of repeats (Murchison et al. 2007). Finally, Dicer knockout mouse ES cell lines have been reported to show increases in steady-state levels of centromeric repeat transcripts (Kanellopoulou et al. 2005;
Murchison et al. 2005). All of these studies implicate Dicer in the control of repetitive elements in mammalian genomes, but it is currently unclear whether or not these effects are direct. Importantly, it has not yet been shown that mammalian Dicer generates the putative repeat-derived siRNAs that have been proposed to mediate the above-mentioned repressive effects.
A brief introduction to mouse ES cells
RNAi is essential for normal function in all mammalian tissue types examined, yet in the majority of these cases, the essential regulatory functions performed by RNAi are unclear. The functions of specific animal miRNAs have been difficult to determine, not
only because they require so little sequence complementarity to influence target gene expression, but because they likely each affect several target genes, making it difficult to computationally determine functionally relevant targets within specific tissues. RNAi also functions through several different types of silencing molecules, affecting gene expression in diverse ways. In mammals, it is unclear how RNAi influences gene expression other than through miRNA-mediated silencing pathways. For example, it seems likely that mammalian piRNAs mediate silencing processes separate from miRNAs, but what these silencing processes are is unclear. The work described in this thesis focuses on defining the roles of RNAi in mouse ES cells. ES cells have a number
of interesting properties relevant to both medicine and the biology of the early embryo; thus an understanding of RNAi-mediated gene regulation in ES cells will likely have broad applications.
ES cells are cultured derivatives of the pre-implantation inner cell mass (ICM) of the blastocyst. The ICM is composed of the progenitor cells that will eventually give rise to a fully developed embryo (Niwa 2007). At the developmental stage from which ES cells are derived, the ICM is in an undifferentiated, epigenetically plastic state; genome-wide DNA methylation levels are a fraction of what they will be in differentiated cells (Kafri et al. 1992; Rougier et al. 1998), and in female ICMs, paternal X chromosomes are re-activated to allow random inactivation in the epiblast (Mak et al. 2004).
ES cells retain several characteristics of the ICM from which they are derived, most notably they exhibit a large degree of epigenetic plasticity and are pluripotent. ES cells can survive with two active X chromosomes, and in the complete absence of DNA methylation, highlighting their epigenetic plasticity (Rastan and Robertson 1985; Lei et
al. 1996; Okano et al. 1999). A direct consequence this plasticity is ES cell pluripotency, defined as the ability of ES cells to give rise to all tissues in a fully developed embryo (Beddington and Robertson 1989). ES cell pluripotency can be maintained for extended periods in culture, and under appropriate conditions, ES cells can be differentiated into a number of cell types in vitro, raising the possibility that human ES cells may someday be used as tissue sources in regenerative therapies (Pera and Trounson 2004; Keller 2005).
A number of factors have been implicated in the maintenance of ES cell pluripotency. The cytokine LIF (for leukemia inhibitory factor) activates a STAT3-dependent transcriptional program that is important for maintenance of pluripotency (Smith et al. 1988; Williams et al. 1988; Niwa et al. 1998). Also, Smad-dependent induction of the Id (for inhibitor of differentiation) genes by BMP4 is critical for maintenance of ES cell pluripotency (Ying et al. 2003). Together, the LIF and BMP4 signaling molecules are sufficient for prolonged cell culture maintenance of ES cell pluripotency in the absence of serum (Ying et al. 2003).
The transcription factors Oct4, Sox2, and Nanog are additional requirements for the maintenance of ES cell pluripotency (Nichols et al. 1998; Chambers et al. 2003; Mitsui et al. 2003; Masui et al. 2007). These three transcription factors frequently co-localize at the promoters of their target genes (Boyer et al. 2005). Oct4/Sox2/Nanog bound genes can be broadly grouped into two classes: genes that are transcriptionally active and likely contribute to ES cell identity, and genes that are transcriptionally silent. The silent class of Oct4/Sox2/Nanog-bound genes are also bound by the Polycomb complex, and are highly enriched in developmental regulator genes whose ES cell expression would likely lead to differentiation (Boyer et al. 2006; Lee et al. 2006b).
RNAi also has a role in the maintenance of ES cell identity. Unlike many differentiated cell types, ES cells can survive deletion of Dicer; however, despite appearing morphologically normal and expressing wild-type levels of the pluripotency markers Oct4 and Nanog, Dicer null ES cells are no longer pluripotent (Kanellopoulou et
al. 2005). Given the demonstrated role of miRNAs as essential regulators of cell-fate
specification, this loss of pluripotency is likely not completely due to a change in ES cell state. Rather, it may partly be due to the inability of putative differentiated precursors to fully differentiate without the presence of additional non-ES cell miRNAs. Nevertheless, ES cells express a set of miRNAs specific to early developmental lineages that are likely important for ES cell identity (Houbaviy et al. 2003; Houbaviy et al. 2005; Tang et al. 2007). Dicer null ES cells also display proliferation defects (Murchison et al. 2005) (see also Chapter 5 of this thesis), again consistent with a cell autonomous role for RNAi in the maintenance of ES cell identity.
References
Ambros, V., Lee, R.C., Lavanway, A., Williams, P.T., and Jewell, D. 2003. MicroRNAs and other tiny endogenous RNAs in C. elegans. Curr Biol 13(10): 807-818. Andl, T., Murchison, E.P., Liu, F., Zhang, Y., Yunta-Gonzalez, M., Tobias, J.W., Andl,
C.D., Seykora, J.T., Hannon, G.J., and Millar, S.E. 2006. The miRNA-processing enzyme dicer is essential for the morphogenesis and maintenance of hair follicles.
Curr Biol 16(10): 1041-1049.
Aravin, A., Gaidatzis, D., Pfeffer, S., Lagos-Quintana, M., Landgraf, P., lovino, N., Morris, P., Brownstein, M.J., Kuramochi-Miyagawa, S., Nakano, T. et al. 2006. A novel class of small RNAs bind to MILI protein in mouse testes. Nature
442(7099): 203-207.
Aravin, A.A., Naumova, N.M., Tulin, A.V., Vagin, V.V., Rozovsky, Y.M., and Gvozdev, V.A. 2001. Double-stranded RNA-mediated silencing of genomic tandem repeats and transposable elements in the D. melanogaster germline. Curr Biol 11(13):
1017-1027.
Aravin, A.A., Sachidanandam, R., Girard, A., Fejes-Toth, K., and Hannon, G.J. 2007. Developmentally regulated piRNA clusters implicate MILI in transposon control.
Science 316(5825): 744-747.
Bagga, S., Bracht, J., Hunter, S., Massirer, K., Holtz, J., Eachus, R., and Pasquinelli, A.E. 2005. Regulation by let-7 and lin-4 miRNAs results in target mRNA degradation.
Cell 122(4): 553-563.
Bartel, D.P. 2004. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116(2): 281-297.
Baskerville, S. and Bartel, D.P. 2005. Microarray profiling of microRNAs reveals
frequent coexpression with neighboring miRNAs and host genes. Rna 11(3): 241-247.
Beddington, R.S. and Robertson, E.J. 1989. An assessment of the developmental potential of embryonic stem cells in the midgestation mouse embryo.
Development 105(4): 733-737.
Bernstein, E., Kim, S.Y., Carmell, M.A., Murchison, E.P., Alcorn, H., Li, M.Z., Mills, A.A., Elledge, S.J., Anderson, K.V., and Hannon, G.J. 2003. Dicer is essential for mouse development. Nat Genet 35(3): 215-217.
Boyer, L.A., Lee, T.I., Cole, M.F., Johnstone, S.E., Levine, S.S., Zucker, J.P., Guenther, M.G., Kumar, R.M., Murray, H.L., Jenner, R.G. et al. 2005. Core transcriptional regulatory circuitry in human embryonic stem cells. Cell 122(6): 947-956.
Boyer, L.A., Plath, K., Zeitlinger, J., Brambrink, T., Medeiros, L.A., Lee, T.I., Levine, S.S., Wernig, M., Tajonar, A., Ray, M.K. et al. 2006. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature
441(7091): 349-353.
Brennecke, J., Aravin, A.A., Stark, A., Dus, M., Kellis, M., Sachidanandam, R., and Hannon, G.J. 2007. Discrete Small RNA-Generating Loci as Master Regulators of Transposon Activity in Drosophila. Cell.
Brennecke, J., Stark, A., Russell, R.B., and Cohen, S.M. 2005. Principles of microRNA-target recognition. PLoS Biol 3(3): e85.
Buhler, M., Verdel, A., and Moazed, D. 2006. Tethering RITS to a nascent transcript initiates RNAi- and heterochromatin-dependent gene silencing. Cell 125(5): 873-886.
Cai, X., Hagedorn, C.H., and Cullen, B.R. 2004. Human microRNAs are processed from capped, polyadenylated transcripts that can also function as mRNAs. Rna 10(12):
1957-1966.
Carmell, M.A., Girard, A., van de Kant, H.J., Bourc'his, D., Bestor, T.H., de Rooij, D.G., and Hannon, G.J. 2007. MIWI2 is essential for spermatogenesis and repression of transposons in the mouse male germline. Dev Cell 12(4): 503-514.
Carmell, M.A., Xuan, Z., Zhang, M.Q., and Hannon, G.J. 2002. The Argonaute family: tentacles that reach into RNAi, developmental control, stem cell maintenance, and tumorigenesis. Gene Dev 16(21): 2733-2742.
Chalfie, M., Horvitz, H.R., and Sulston, J.E. 1981. Mutations that lead to reiterations in the cell lineages of C. elegans. Cell 24(1): 59-69.
Chambers, I., Colby, D., Robertson, M., Nichols, J., Lee, S., Tweedie, S., and Smith, A. 2003. Functional expression cloning of Nanog, a pluripotency sustaining factor in
embryonic stem cells. Cell 113(5): 643-655.
Chan, S.W., Zilberman, D., Xie, Z., Johansen, L.K., Carrington, J.C., and Jacobsen, S.E. 2004. RNA silencing genes control de novo DNA methylation. Science
303(5662): 1336.
Chendrimada, T.P., Finn, K.J., Ji, X., Baillat, D., Gregory, R.I., Liebhaber, S.A., Pasquinelli, A.E., and Shiekhattar, R. 2007. MicroRNA silencing through RISC recruitment of eIF6. Nature 447(7146): 823-828.
Chendrimada, T.P., Gregory, R.I., Kumaraswamy, E., Norman, J., Cooch, N., Nishikura, K., and Shiekhattar, R. 2005. TRBP recruits the Dicer complex to Ago2 for microRNA processing and gene silencing. Nature 436(7051): 740-744.
Cheng, H.Y., Papp, J.W., Varlamova, O., Dziema, H., Russell, B., Curfman, J.P., Nakazawa, T., Shimizu, K., Okamura, H., Impey, S. et al. 2007. microRNA modulation of circadian-clock period and entrainment. Neuron 54(5): 813-829. Colmenares, S.U., Buker, S.M., Buhler, M., Dlakic, M., and Moazed, D. 2007. Coupling
of double-stranded RNA synthesis and siRNA generation in fission yeast RNAi.
Mol Cell 27(3): 449-461.
Dalmay, T., Hamilton, A., Rudd, S., Angell, S., and Baulcombe, D.C. 2000. An RNA-dependent RNA polymerase gene in Arabidopsis is required for
posttranscriptional gene silencing mediated by a transgene but not by a virus. Cell
101(5): 543-553.
Deleris, A., Gallego-Bartolome, J., Bao, J., Kasschau, K.D., Carrington, J.C., and Voinnet, 0. 2006. Hierarchical action and inhibition of plant Dicer-like proteins in antiviral defense. Science 313(5783): 68-71.
Deng, W. and Lin, H. 2002. miwi, a murine homolog of piwi, encodes a cytoplasmic protein essential for spermatogenesis. Dev Cell 2(6): 819-830.
Doench, J.G., Petersen, C.P., and Sharp, P.A. 2003. siRNAs can function as miRNAs.
Genes Dev 17(4): 438-442.
Doench, J.G. and Sharp, P.A. 2004. Specificity ofmicroRNA target selection in translational repression. Genes Dev 18(5): 504-511.
Duchaine, T.F., Wohlschlegel, J.A., Kennedy, S., Bei, Y., Conte, D., Jr., Pang, K., Brownell, D.R., Harding, S., Mitani, S., Ruvkun, G. et al. 2006. Functional proteomics reveals the biochemical niche of C. elegans DCR-1 in multiple small-RNA-mediated pathways. Cell 124(2): 343-354.
Fagard, M., Boutet, S., Morel, J.B., Bellini, C., and Vaucheret, H. 2000. AGO , QDE-2, and RDE-1 are related proteins required for post-transcriptional gene silencing in plants, quelling in fungi, and RNA interference in animals. Proc Natl Acad Sci U
SA 97(21): 11650-11654.
Farh, K.K., Grimson, A., Jan, C., Lewis, B.P., Johnston, W.K., Lim, L.P., Burge, C.B., and Bartel, D.P. 2005. The widespread impact of mammalian MicroRNAs on mRNA repression and evolution. Science 310(5755): 1817-1821.
Forstemann, K., Horwich, M.D., Wee, L., Tomari, Y., and Zamore, P.D. 2007.
Drosophila microRNAs are sorted into functionally distinct argonaute complexes after production by dicer-1. Cell 130(2): 287-297.
Galiana-Arnoux, D., Dostert, C., Schneemann, A., Hoffmann, J.A., and Imler, J.L. 2006. Essential function in vivo for Dicer-2 in host defense against RNA viruses in
Giraldez, A.J., Mishima, Y., Rihel, J., Grocock, R.J., Van Dongen, S., Inoue, K., Enright, A.J., and Schier, A.F. 2006. Zebrafish MiR-430 promotes deadenylation and clearance of maternal mRNAs. Science 312(5770): 75-79.
Girard, A., Sachidanandam, R., Hannon, G.J., and Carmell, M.A. 2006. A germline-specific class of small RNAs binds mammalian Piwi proteins. Nature 442(7099):
199-202.
Griffiths-Jones, S. 2004. The microRNA Registry. Nucleic Acids Res 32(Database issue): D109-111.
Grimaud, C., Bantignies, F., Pal-Bhadra, M., Ghana, P., Bhadra, U., and Cavalli, G. 2006. RNAi components are required for nuclear clustering of Polycomb group response elements. Cell 124(5): 957-971.
Grimson, A., Farh, K.K., Johnston, W.K., Garrett-Engele, P., Lim, L.P., and Bartel, D.P. 2007. MicroRNA targeting specificity in mammals: determinants beyond seed pairing. Mol Cell 27(1): 91-105.
Grishok, A., Pasquinelli, A.E., Conte, D., Li, N., Parrish, S., Ha, I., Baillie, D.L., Fire, A., Ruvkun, G., and Mello, C.C. 2001. Genes and mechanisms related to RNA
interference regulate expression of the small temporal RNAs that control C. elegans developmental timing. Cell 106(1): 23-34.
Grishok, A., Sinskey, J.L., and Sharp, P.A. 2005. Transcriptional silencing of a transgene by RNAi in the soma of C. elegans. Genes Dev 19(6): 683-696.
Gunawardane, L.S., Saito, K., Nishida, K.M., Miyoshi, K., Kawamura, Y., Nagami, T., Siomi, H., and Siomi, M.C. 2007. A slicer-mediated mechanism for repeat-associated siRNA 5' end fobrmation in Drosophila. Science 315(5818): 1587-1590. Hall, I.M., Noma, K., and Grewal, S.I.S. 2003. RNA interference machinery regulates
chromosome dynamics during mitosis and meiosis in fission yeast. P Natl Acad Sci USA 100(1): 193-198.
Hall, I.M., Shankaranarayana, G.D., Noma, K., Ayoub, N., Cohen, A., and Grewal, S.I. 2002. Establishment and maintenance of a heterochromatin domain. Science 297(5590): 2232-2237.
Hamilton, A., Voinnet, O., Chappell, L., and Baulcombe, D. 2002. Two classes of short interfering RNA in RNA silencing. Embo J21(17): 4671-4679.
Han, J., Lee, Y., Yeom, K.H., Nam, J.W., Heo, I., Rhee, J.K., Sohn, S.Y., Cho, Y., Zhang, B.T., and Kim, V.N. 2006. Molecular basis for the recognition of primary microRNAs by the Drosha-DGCR8 complex. Cell 125(5): 887-901.
Harfe, B.D., McManus, M.T., Mansfield, J.H., Hornstein, E., and Tabin, C.J. 2005. The RNaseIII enzyme Dicer is required for morphogenesis but not patterning of the vertebrate limb. Proc Natl Acad Sci USA 102(31): 10898-10903.
Harris, K.S., Zhang, Z., McManus, M.T., Harfe, B.D., and Sun, X. 2006. Dicer function is essential for lung epithelium morphogenesis. Proc Natl Acad Sci USA 103(7): 2208-2213.
Houbaviy, H.B., Dennis, L., Jaenisch, R., and Sharp, P.A. 2005. Characterization of a highly variable eutherian microRNA gene. Rna 11(8): 1245-1257.
Houbaviy, H.B., Murray, M.F., and Sharp, P.A. 2003. Embryonic stem cell-specific
MicroRNAs. Developmental Cell 5(2): 351-358.
Humphreys, D.T., Westman, B.J., Martin, D.I., and Preiss, T. 2005. MicroRNAs control translation initiation by inhibiting eukaryotic initiation factor 4E/cap and poly(A) tail function. Proc Natl Acad Sci USA 102(47): 16961-16966.
Hutvagner, G., McLachlan, J., Pasquinelli, A.E., Balint, E., Tuschl, T., and Zamore, P.D. 2001. A cellular function for the RNA-interference enzyme Dicer in the
maturation of the let-7 small temporal RNA. Science 293(5531): 834-838. Hutvagner, G. and Zamore, P.D. 2002. A microRNA in a multiple-turnover RNAi
enzyme complex. Science 297(5589): 2056-2060.
Irvine, D.V., Zaratiegui, M., Tolia, N.H., Goto, D.B., Chitwood, D.H., Vaughn, M.W., Joshua-Tor, L., and Martienssen, R.A. 2006. Argonaute slicing is required for heterochromatic silencing and spreading. Science 313(5790): 1134-1137. Jakymiw, A., Lian, S., Eystathioy, T., Li, S., Satoh, M., Hamel, J.C., Fritzler, M.J., and
Chan, E.K. 2005. Disruption of GW bodies impairs mammalian RNA interference. Nat Cell Biol 7(12): 1267-1274.
Jia, S., Noma, K., and Grewal, S.I. 2004. RNAi-independent heterochromatin nucleation by the stress-activated ATF/CREB family proteins. Science 304(5679):
1971-1976.
Kafri, T., Ariel, M., Brandeis, M., Shemer, R., Urven, L., McCarrey, J., Cedar, H., and Razin, A. 1992. Developmental pattern of gene-specific DNA methylation in the mouse embryo and germ line. Genes Dev 6(5): 705-714.
Kanellopoulou, C., Muljo, S.A., Kung, A.L., Ganesan, S., Drapkin, R., Jenuwein, T., Livingston, D.M., and Rajewsky, K. 2005. Dicer-deficient mouse embryonic stem cells are defective in differentiation and centromeric silencing. Genes Dev 19(4): 489-501.
Karres, J.S., Hilgers, V., Carrera, I., Treisman, J., and Cohen, S.M. 2007. The conserved microRNA miR-8 tunes atrophin levels to prevent neurodegeneration in
Drosophila. Cell 131(1): 136-145.
Keller, G. 2005. Embryonic stem cell differentiation: emergence of a new era in biology and medicine. Genes Dev 19(10): 1129-1155.
Kertesz, M., lovino, N., Unnerstall, U., Gaul, U., and Segal, E. 2007. The role of site accessibility in microRNA target recognition. Nat Genet 39(10): 1278-1284. Ketting, R.F., Fischer, S.E., Bernstein, E., Sijen, T., Hannon, G.J., and Plasterk, R.H.
2001. Dicer functions in RNA interference and in synthesis of small RNA involved in developmental timing in C. elegans. Genes Dev 15(20): 2654-2659. Ketting, R.F., Haverkamp, T.H., van Luenen, H.G., and Plasterk, R.H. 1999. Mut-7 of C.
elegans, required for transposon silencing and RNA interference, is a homolog of Werner syndrome helicase and RNaseD. Cell 99(2): 133-141.
Ketting, R.F. and Plasterk, R.H. 2000. A genetic link between co-suppression and RNA interference in C. elegans. Nature 404(6775): 296-298.
Khvorova, A., Reynolds, A., and Jayasena, S.D. 2003. Functional siRNAs and miRNAs exhibit strand bias. Cell 115(2): 209-216.
Kiriakidou, M., Tan, G.S., Lamprinaki, S., De Planell-Saguer, M., Nelson, P.T., and Mourelatos, Z. 2007. An mRNA m7G cap binding-like motif within human Ago2 represses translation. Cell 129(6): 1141-1151.
Kloosterman, W.P. and Plasterk, R.H. 2006. The diverse functions of microRNAs in animal development and disease. Dev Cell 11(4): 441-450.
Kumar, M.S., Lu, J., Mercer, K.L., Golub, T.R., and Jacks, T. 2007. Impaired microRNA processing enhances cellular transformation and tumorigenesis. Nat Genet 39(5): 673-677.
Kuramochi-Miyagawa, S., Kimura, T., Ijiri, T.W., Isobe, T., Asada, N., Fujita, Y., Ikawa, M., Iwai, N., Okabe, M., Deng, W. et al. 2004. Mili, a mammalian member of piwi family gene, is essential for spermatogenesis. Development 131(4): 839-849. Lakatos, L., Szittya, G., Silhavy, D., and Burgyan, J. 2004. Molecular mechanism of
RNA silencing suppression mediated by p19 protein of tombusviruses. Embo J 23(4): 876-884.
Lander, E.S. Linton, L.M. Birren, B. Nusbaum, C. Zody, M.C. Baldwin, J. Devon, K. Dewar, K. Doyle, M. FitzHugh, W. et al. 2001. Initial sequencing and analysis of the human genome. Nature 409(6822): 860-921.
Lau, N.C., Seto, A.G., Kim, J., Kuramochi-Miyagawa, S., Nakano, T., Bartel, D.P., and Kingston, R.E. 2006. Characterization of the piRNA complex from rat testes.
Science 313(5785): 363-367.
Lee, R.C., Feinbaum, R.L., and Ambros, V. 1993. The C. elegans heterochronic gene lin-4 encodes small RNAs with antisense complementarity to lin-1lin-4. Cell 75(5): 8lin-43-
843-854.
Lee, R.C., Hammell, C.M., and Ambros, V. 2006a. Interacting endogenous and exogenous RNAi pathways in Caenorhabditis elegans. Rna 12(4): 589-597. Lee, T.I., Jenner, R.G., Boyer, L.A., Guenther, M.G., Levine, S.S., Kumar, R.M.,
Chevalier, B., Johnstone, S.E., Cole, M.F., Isono, K. et al. 2006b. Control of developmental regulators by Polycomb in human embryonic stem cells. Cell 125(2): 301-313.
Lee, Y., Ahn, C., Han, J., Choi, H., Kim, J., Yim, J., Lee, J., Provost, P., Radmark, O., Kim, S. et al. 2003. The nuclear RNase III Drosha initiates microRNA
processing. Nature 425(6956): 415-419.
Lee, Y., Kim, M., Han, J., Yeom, K.H., Lee, S., Baek, S.H., and Kim, V.N. 2004. MicroRNA genes are transcribed by RNA polymerase II. Embo J23(20): 4051-4060.
Lei, H., Oh, S.P., Okano, M., Juttermann, R., Goss, K.A., Jaenisch, R., and Li, E. 1996. De novo DNA cytosine methyltransferase activities in mouse embryonic stem cells. Development 122(10): 3195-3205.
Leung, A.K. and Sharp, P.A. 2007. microRNAs: a safeguard against turmoil? Cell
130(4): 581-585.
Lewis, B.P., Burge, C.B., and Bartel, D.P. 2005. Conserved seed pairing, often flanked by adenosines, indicates that thousands of human genes are microRNA targets.
Cell 120(1): 15-20.
Lewis, B.P., Shih, I.H., Jones-Rhoades, M.W., Bartel, D.P., and Burge, C.B. 2003. Prediction of mammalian microRNA targets. Cell 115(7): 787-798.
Li, C.F., Pontes, O., El-Shami, M., Henderson, I.R., Bernatavichute, Y.V., Chan, S.W., Lagrange, T., Pikaard, C.S., and Jacobsen, S.E. 2006. An ARGONAUTE4-containing nuclear processing center colocalized with Cajal bodies in Arabidopsis
thaliana. Cell 126(1): 93-106.
Li, H., Li, W.X., and Ding, S.W. 2002. Induction and suppression of RNA silencing by
an animal virus. Science 296(5571): 1319-1321.
Lim, L.P., Lau, N.C., Garrett-Engele, P., Grimson, A., Schelter, J.M., Castle, J., Bartel, D.P., Linsley, P.S., and Johnson, J.M. 2005. Microarray analysis shows that some