Weed Hosts for Onion Thrips (Thysanoptera: Thripidae) and Their
Potential Role in the Epidemiology of Iris Yellow Spot Virus in an
Onion Ecosystem
ERIK A. SMITH,1,2A. DITOMMASO,3M. FUCHS,4A. M. SHELTON,1ANDB. A. NAULT1
Environ. Entomol. 40(2): 194Ð203 (2011); DOI: 10.1603/EN10246
ABSTRACT Onion thrips, Thrips tabaci Lindeman, is a key foliage-feeding pest of onion worldwide and the principal vector of a serious onion pathogen, Iris yellow spot virus (IYSV). Long-term management of T. tabaci and IYSV will require an understanding of T. tabaci ecology and IYSV epidemiology in onion ecosystems. This study focused on identifying winter-annual, biennial and perennial weed species that host both T. tabaci and IYSV. Unlike summer-annual weeds, weeds with these habits survive overwinter and could serve as a green bridge for IYSV to survive between onion-growing seasons. T. tabaci larvae and adults were sampled every two weeks from 69 weed species in Þve areas located adjacent to onion Þelds in western New York in 2008 and 2009. Twenty-Þve of the 69 weed species were identiÞed as hosts for T. tabaci larvae and populations were highest on the Brassicaceous weeds, Barbarea vulgaris Ait. f., Sinapis arvensis L., and Thalspi arvense L. None of these species are hosts for IYSV. Four of the 25 weed species were hosts for both T. tabaci larval populations and IYSV: common burdock, Arctium minus Bernh., dandelion, Taraxacum officinale G.H. Weber ex Wiggers, curly dock, Rumex crispus L., and chicory, Cichorium intybus L. Of these four weed species, T. officinaleand A. minus may play an important role in the epidemiology of IYSV in New York onion Þelds because they may survive between onion-growing seasons, they are relatively abundant in the landscape, and they support relatively high densities of T. tabaci.
KEY WORDS Thrips tabaci, population ecology, Tospovirus, Allium cepa
Onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae), is a major pest of onion, Allium cepa L., and other Allium spp. worldwide (Lewis 1997). Thrips damage onion plants by feeding on leaf tissue, often leading to signiÞcant reductions in bulb yield (Fournier et al. 1995, Childers 1997). T. tabaci is the only thrips pest of onion in New York. Populations of T. tabacioverwinter in crops and weedy vegetation (North and Shelton 1986) as well as in the soil within and/or near onion Þelds (Larentzaki et al. 2007). Adult T. tabaci are observed from late March through May on volunteer onion plants (Larentzaki et al. 2007), which sprout from bulbs remaining in the Þeld from the previous season. During this time, onion seed is planted and onion plants are transplanted in New York. Colonization of onion crops begins in June and populations are found on the crop until harvest from late-July through September. After harvest, T. tabaci
adults have been observed on several weed species including common lambsquarters, Chenopodium al-bumL., evening primrose, Oenothera biennis L., yellow nutsedge, Cyperus esculentus L., and smooth pigweed, Amaranthus hybridus L. (Larentzaki et al. 2007). Whether T. tabaci reproduce on these weeds or on other weeds in this cropping system has not been determined.
T. tabaciis the only reported vector of Iris yellow spot virus(IYSV) (Bunyaviridae: Tospovirus) (Kritz-man et al. 2001), a serious yield-reducing pathogen of onion and other Allium crops worldwide (Gent et al. 2006). IYSV is transmitted by T. tabaci in a persistent, circulative manner (Ullman et al. 1992, WhitÞeld et al. 2005). Thrips are known to acquire tospoviruses as Þrst instars and transmit them as second instars or adults (Ullman et al. 1992, Wijkamp et al. 1993). IYSV is not known to be seed-transmitted (Gent et al. 2004), nor is it easily transmitted by mechanical means (Gent et al. 2006).
IYSV was Þrst identiÞed in Brazil in 1981 (de A´ vila et al. 1981) and has since spread to regions of onion production across the globe (Gent et al. 2006). IYSV was Þrst detected in New York in 2006 (Hoepting et al. 2007) and has since been found throughout onion-producing regions in the state (Nault et al. 2008). In 2007 and 2008, IYSV was not detected in onion Þelds 1Department of Entomology, Cornell University, New York State
Agricultural Experiment Station, 630 W. North Street, Geneva, NY 14456.
2Corresponding author, e-mail: [email protected].
3Department of Crop and Soil Sciences, Cornell University, 903
BradÞeld Hall, Ithaca, NY 14853.
4Department of Plant Pathology and Plant-Microbe Biology,
Cor-nell University, New York State Agricultural Experiment Station, 630 W. North Street, Geneva, NY 14456.
until mid-June to mid-July in New York and levels in these Þelds were very low until early to mid August; ⬍12% of samples within a Þeld tested positive at this time (Hsu et al. 2010). The incidence of IYSV in many onion Þelds increased dramatically during the second half of August with some Þelds reaching levels⬎90% by harvest (Hsu et al. 2010).
Potential sources of IYSV in New York recently have become better understood. IYSV may be rein-troduced annually via infected onion transplants orig-inating in states like Arizona where the virus is estab-lished (Pappu and Matheron 2008). Onion transplants imported into New York from Arizona have tested positive for IYSV (Hsu et al. 2011). Transplants ac-count for⬇15% of New YorkÕs onion crop and only 0.04% of transplants tested positive for IYSV in 2007 and 2008 (Hsu et al. 2011).
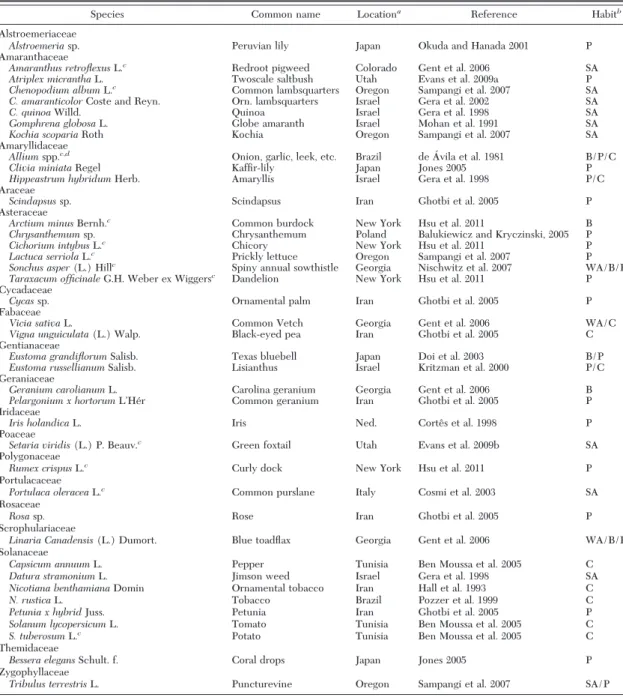
There also are sources of IYSV that may permit this virus to bridge seasons in New York. For example, volunteer onion plants in nonrotated Þelds and in onion cull piles may be infected with IYSV (Hsu et al. 2011). These sources could support an early genera-tion of onion thrips whose viruliferous adults could disperse to and infect the main onion crop. Another possible source that could enable IYSV to bridge sea-sons in New York includes winter-annual, biennial, or perennial weeds. In addition to onion, there are at least 46 plant species in 19 families known to host IYSV (Table 1), many of which are weed species not in the genus Allium (Gent et al. 2006). In 2007 and 2008, Hsu et al. (2011) tested a number of commonly occurring winter-annual, biennial, and perennial weed species for IYSV in New York. Four species tested positive including common burdock, Arctium minus Bernh., chicory, Cichorium intybus L., curly dock, Rumex crispusL., and dandelion, Taraxacum officinale G.H. Weber ex Wiggers (Table 1). The frequency of in-fection in each weed species was not reported.
Winter-annual, biennial, and perennial weed spe-cies could be available as hosts for T. tabaci early in the spring and late in the fall when the onion crop is not available as a host. These weed species would have the potential to be epidemiological bridges between crop-ping seasons for IYSV because they survive through the winter in New York. Potential candidates include the four species identiÞed by Hsu et al. (2011) plus two other species: the perennial, prickly lettuce, Lac-tuca serriolaL. (Sampangi and Mohan 2007), and the winter-annual, spiny sowthistle, Sonchus asper (L.) Hill (Nischwitz et al. 2007). Others may exist as well, but have yet to be identiÞed.
In contrast to the aforementioned weed species, summer-annual weeds are unlikely to be signiÞcant hosts in the epidemiology of IYSV. These weeds would have to be infected by viruliferous T. tabaci in the summer and subsequent generations of T. tabaci adults would then need to disperse and transmit the virus into the onion crop. Such a scenario is conceivable if viruliferous T. tabaci preferentially colonize summer-annual weeds rather than onion; however, it would likely not occur until late in the season, perhaps when the onion crop is near harvest or already harvested.
The objectives of this study were to: (1) identify weed species that host T. tabaci larvae, especially win-ter-annual, biennial, and perennial species; (2) deter-mine the overall abundance and temporal patterns of T. tabacidensities on weed species known to be re-productive hosts for T. tabaci and hosts for IYSV; and (3) identify weed species that may be the best can-didates for impacting the epidemiology of IYSV in the onion ecosystem.
Materials and Methods
Sampling Location and Period. In 2008 and 2009,
thrips were collected and weeds sampled from Þve sites located in the Elba muck region of western New York (43.1N, 78.1W), the second largest onion pro-ducing region in the state. Sites were at least 1.5 km apart. Data collection sites were selected for their proximity to onion Þelds and were located in areas of fallow land 10 to 50 m parallel and adjacent to onion Þelds. Each site was sampled in both years of this study starting several weeks before onions were colonized by thrips and ending several weeks after onions were harvested. In addition to sampling thrips on weeds, 15 onion plants in the Þeld nearest to the weed sample site were visually inspected for T. tabaci larvae and the number per plant was recorded. In 2008, data were collected every 2 wk from 9 May to 30 August and again on 26 September (10 sampling dates). In 2009, data were collected every 2 wk from 18 May through 5 October (11 sampling dates). This schedule was chosen to ensure that data were collected before, during, and after the onion growing season.
Sampling Weeds and Thrips. At each of the Þve
collection sites in both years, entire plants or leaf subsamples were collected and the numbers of T. tabaciadults and larvae were recorded. No more than Þve plants per weed species were collected per col-lection site. Plants within a colcol-lection site were com-bined to produce one composite weed species-sample per site per sampling date (n⫽ 5 per sampling date). Data were represented as the number of thrips per plant per sampling date. Not all weed species were present at each site, and because of the seasonal life-cycle of some species, not all species were collected on each sample date. For example, winter-annual weeds were not usually present in July and early-August. The composite samples were placed in a 5.68 liters poly-propylene container (Sterilite Corporation, Townsend, MA) and maintained in an environmental chamber at 29⬚C, 78% RH, and 16:8 L:D until analysis. Lids of containers were Þtted with thrips-proof screen for ventilation (150⫻ 150 m). Entire plants were collected when possible, but larger plants required the collection of a representative sub-sample of leaves. In such cases, leaf numbers were recorded for each sam-pled plant so thrips populations could be extrapolated to a per-plant basis for further comparisons.
Identifying T. tabaci from Weeds. If T. tabaci larvae
were collected from a weed species, the weed was considered as a reproductive host. Because identiÞ-cation keys do not exist for T. tabaci larvae, thrips
larvae were removed from plant samples and reared to the adult stage. Using a Þne paintbrush, up to 30 larvae per composite weed sample were transferred to ven-tilated petri dishes containing sterilized leaf disks of cabbage, Brassica oleracea L. (Capitata group), and reared to adulthood following the procedure
de-scribed in Nault et al. (2006). Based on experience rearing T. tabaci larvae, those larvae that were clearly not T. tabaci were not collected nor counted. Upon reaching the adult stage, thrips were identiÞed and counted according to species (Moritz et al. 2001). The percentage of thrips larvae determined to be T. tabaci Table 1. Plant species reported to be hosts of IYSV
Species Common name Locationa
Reference Habitb
Alstroemeriaceae
Alstroemeriasp. Peruvian lily Japan Okuda and Hanada 2001 P
Amaranthaceae
Amaranthus retroflexusL.c Redroot pigweed Colorado Gent et al. 2006 SA
Atriplex micranthaL. Twoscale saltbush Utah Evans et al. 2009a P
Chenopodium albumL.c
Common lambsquarters Oregon Sampangi et al. 2007 SA
C. amaranticolorCoste and Reyn. Orn. lambsquarters Israel Gera et al. 2002 SA
C. quinoaWilld. Quinoa Israel Gera et al. 1998 SA
Gomphrena globosaL. Globe amaranth Israel Mohan et al. 1991 SA
Kochia scopariaRoth Kochia Oregon Sampangi et al. 2007 SA
Amaryllidaceae
Alliumspp.c,d Onion, garlic, leek, etc. Brazil de A´ vila et al. 1981 B/P/C
Clivia miniataRegel KafÞr-lily Japan Jones 2005 P
Hippeastrum hybridumHerb. Amaryllis Israel Gera et al. 1998 P/C
Araceae
Scindapsussp. Scindapsus Iran Ghotbi et al. 2005 P
Asteraceae
Arctium minusBernh.c Common burdock New York Hsu et al. 2011 B
Chrysanthemumsp. Chrysanthemum Poland Balukiewicz and Kryczinski, 2005 P
Cichorium intybusL.c Chicory New York Hsu et al. 2011 P
Lactuca serriolaL.c Prickly lettuce Oregon Sampangi et al. 2007 P
Sonchus asper(L.) Hillc Spiny annual sowthistle Georgia Nischwitz et al. 2007 WA/B/P Taraxacum officinaleG.H. Weber ex Wiggersc Dandelion New York Hsu et al. 2011 P
Cycadaceae
Cycassp. Ornamental palm Iran Ghotbi et al. 2005 P
Fabaceae
Vicia sativaL. Common Vetch Georgia Gent et al. 2006 WA/C
Vigna unguiculata(L.) Walp. Black-eyed pea Iran Ghotbi et al. 2005 C
Gentianaceae
Eustoma grandiflorumSalisb. Texas bluebell Japan Doi et al. 2003 B/P
Eustoma russellianumSalisb. Lisianthus Israel Kritzman et al. 2000 P/C
Geraniaceae
Geranium carolianumL. Carolina geranium Georgia Gent et al. 2006 B
Pelargonium x hortorumLÕHe´r Common geranium Iran Ghotbi et al. 2005 P
Iridaceae
Iris holandicaL. Iris Ned. Corteˆs et al. 1998 P
Poaceae
Setaria viridis(L.) P. Beauv.c Green foxtail Utah Evans et al. 2009b SA
Polygonaceae
Rumex crispusL.c Curly dock New York Hsu et al. 2011 P
Portulacaceae
Portulaca oleraceaL.c Common purslane Italy Cosmi et al. 2003 SA
Rosaceae
Rosasp. Rose Iran Ghotbi et al. 2005 P
Scrophulariaceae
Linaria Canadensis(L.) Dumort. Blue toadßax Georgia Gent et al. 2006 WA/B/P
Solanaceae
Capsicum annuumL. Pepper Tunisia Ben Moussa et al. 2005 C
Datura stramoniumL. Jimson weed Israel Gera et al. 1998 SA
Nicotiana benthamianaDomin Ornamental tobacco Iran Hall et al. 1993 C
N. rusticaL. Tobacco Brazil Pozzer et al. 1999 C
Petunia x hybridJuss. Petunia Iran Ghotbi et al. 2005 P
Solanum lycopersicumL. Tomato Tunisia Ben Moussa et al. 2005 C
S. tuberosumL.c Potato Tunisia Ben Moussa et al. 2005 C
Themidaceae
Bessera elegansSchult. f. Coral drops Japan Jones 2005 P
Zygophyllaceae
Tribulus terrestrisL. Puncturevine Oregon Sampangi et al. 2007 SA/P
aLocation of Þrst detection.
bMost common life history: B⫽ biennial, P ⫽ perennial, WA ⫽ winter-annual, SA ⫽ summer-annual, C ⫽ crop. Combinations indicate that
multiple life histories are common.
cSpecies found in the Elba Muck ecosystem, Elba, NY (43.1N, 78.1W) (E.A. Smith, unpublished data).
dIncludes seven species of Allium: cultivated onion, A. cepa (de A´ vila et al. 1981; Hall et al. 1993), shallot, A. cepa var. ascalonicum
(Robene-Soustrade et al. 2006), leek, A. porrum (Coutts et al. 2003), garlic, A. sativum (Robene-Soustrade et al. 2006), and wild onions, A. altaicum, A. pskemense,and A. vavilovii (Pappu et al. 2006).
was multiplied by the total initial number of larvae in the petri dish to estimate the percentage of thrips larvae that were T. tabaci. These numbers were di-vided by the number of plants in the sample to esti-mate the number of T. tabaci larvae per plant. For example, let us say that there were 100 thrips larvae (excluding thrips that were clearly not T. tabaci) de-tected in the original container that had Þve plants of the same weed species. Thirty larvae were removed and placed into petri dishes until they reached adult-hood. Only 24 of the 30 larvae survived to the adult stage and three of the survivors were T. tabaci (⫽3/24 or 12.5% of surviving thrips were T. tabaci). The Þnal estimate of the number of T. tabaci in the original container was 12.5 thrips or 2.5 per plant (⫽12.5 thrips/5 plants). Voucher specimens are maintained in the Department of Entomology, Cornell University, NYSAES (Geneva, NY).
Weed Population Density Estimates. On each
sam-ple date, weed densities at each of the Þve sites were estimated by recording the number of individuals of each weed species in 0.5⫻ 0.5 m quadrats placed every 10 m along a 90 m linear transect parallel to the re-spective onion Þelds. Transects at each of the Þve collection sites occurred in the same locations both years and were adjacent to areas where weeds were collected for thrips population surveys. Numbers of plants per species in all 10 quadrats were totaled at each site and then across all sites on each sampling date. These totals were divided by the total area sam-pled per date (12.5 m2) to estimate the number of
plants per m2. Weed densities are presented on a
plants per hectare basis by extrapolating the mean number of plants per species on each sampling date. Values for all sampling dates were averaged to obtain an estimated season mean number of plants per hect-are for each weed species.
Identifying Weed Species That May be Important in IYSV Epidemiology. A weed species that may be an
important epidemiological source for IYSV must be a reproductive host for T. tabaci, but also may need to be relatively abundant in the landscape. The potential importance of a weed species being a source of IYSV was estimated by calculating numbers of T. tabaci larvae per hectare for each weed species. T. tabaci larval populations (larvae per plant per sampling date) on IYSV-susceptible winter-annual, biennial and pe-rennial weed species were multiplied by each speciesÕ respective population density (plants per hectare) to determine the number of larvae per hectare per sam-pling date.
Results
Larval T. tabaci Population Survey on Weeds.
Sixty-nine winter-annual, biennial, and perennial weed spe-cies and four crop spespe-cies were sampled over 2 yr. No wild Allium spp. were observed in this study. Twenty-Þve weed species and three crop species were iden-tiÞed as hosts for T. tabaci larvae (Table 2). No T. tabacilarvae were observed on 44 weed species and one crop species (Appendix 1). Of the 25 weed species
that were hosts for T. tabaci larvae, only common burdock, chicory, curly dock, and dandelion are known hosts for IYSV (Hsu et al. 2011). T. tabaci larvae were identiÞed on common burdock and dandelion in both 2008 and 2009, but were found on curly dock only in 2008 and on chicory only in 2009 (Fig. 1). Mean densities of T. tabaci larvae were highest on common burdock in 2008 and highest on both common burdock and dandelion in 2009.
Though they are not likely to be important epide-miological sources for IYSV, T. tabaci larvae were identiÞed on Þve of 20 summer-annual weeds: redroot pigweed, Amaranthus retroflexus L. (Amaranthaceae), common lambsquarters (Amaranthaceae), ladyÕs-thumb, Persicaria maculosaL. (Polygonaceae), wild buckwheat, Polygonum convolvulus L. (Polygonaceae), and green smartweed, P. scabrum Moench (Polygonaceae) (data available in Smith 2010).
Seasonal Dynamics of T. tabaci on Weeds and On-ions. In 2008 and 2009, populations of T. tabaci were
much greater on onion plants than on weeds (Figs. 2BÐD). Peaks in adult T. tabaci populations on weeds were recorded on common burdock, chicory, curly dock, and dandelion early in the season (27 May through 15 June), and again late in the season when onions were harvested (18 August through 26 Sep-tember) (Fig. 2A,B). On common burdock, peaks in adult populations occurred on 27 May, 18 August and 26 September 2008, and on 15 June and 8 September 2009. On chicory, adults were only observed on 9 June and 26 September 2008 and on 15 June and 10 August 2009. On curly dock, no observable peaks were de-tected in 2008 and no T. tabaci adults were observed in 2009. On dandelion, a peak occurred on 30 August 2008 and 21 September 2009. Adult onion thrips were present but not recorded on onion crops in 2008, but in 2009 adult populations on onion peaked to a level of 2.5 T. tabaci per plant on 13 July 2009 and peaked again to a level of 42.5 per plant on 8 September.
Peaks in larval populations on these weed species followed peaks in adult populations both early in the growing season (June) and during onion harvest (late August through September in 2008 and 2009, and into October in 2009) (Fig. 2C,D). On common burdock, peaks in larval populations occurred on 9 June and 18 August 2008, and on 15 June, 8 September, and 5 October 2009. On chicory, larvae were not observed in 2008, and were observed only on 10 August 2009. On curly dock, T. tabaci larvae were only observed on 9 June, 2008, and were not observed in 2009. On dan-delion, peaks in larval populations occurred on 9 June and 30 August 2008, and on only 29 June 2009. Larval T. tabaci populations were Þrst recorded on onion crops in early June of both years of this study. Popu-lations peaked around 22 July and again on 18 August 2008, and around 27 July and 8 September 2009.
Estimated Weed Population Densities of Potential IYSV Sources. Dandelion plants were 21 and 44 times
more abundant than common burdock and six and eight times more abundant than chicory in 2008 and 2009, respectively (Fig. 3). Dandelion plants were 341 times more abundant than curly dock in 2008.
Curly dock was quite rare in survey transects in 2008, and thus it is not particularly surprising that it was absent in 2009 despite its presence in the region.
Estimated Population Densities of T. tabaci Larvae on IYSV-Positive Weed Hosts on a Per-Area Basis. In
2008 and 2009, the estimated populations of T. tabaci larvae per hectare on common burdock and dandelion
were considerably higher than those on chicory and curly dock (Fig. 4).
Discussion
T. tabacilarvae were detected on 25 winter-annual, biennial, and perennial weed species representing 14 families. These results indicate that despite a prefer-Table 2. Winter-annual, biennial, and perennial weed species found to be reproductive hosts of T. tabaci in onion, Allium cepa, in western New York
Family
Common name Habita Periodb
Mean no. of T. tabaci larvae per plantc
Species 2008 SEd 2009 SE
d
Weed species Apiaceae
Conium maculatumL. Poison hemlock B All 0.54 0.36 0.52 0.23
Asclepiadaceae
Asclepias syriacaL. Common milkweed P All 0.36 0.24 0 0
Asteraceae
Arctium minusBernh.e Common burdock B All 0.36 0.22 0.46 0.25
Cichorium intybusL.e Chicory P All 0 0 0.02 0.02
Cirsium arvense(L.) Scop. Canada thistle P All 0 0 0.05 0.05
Solidago CanadensisL. Goldenrod P All 0.06 0.03 0.09 0.09
Taraxacum officinaleG.H. Weber ex Wiggerse
Dandelion P All 0.08 0.05 0.44 0.44
Brassicaceae
Barbarea vulgarisAit. f. Yellow rocket WA/B/P All 0.23 0.14 2.45 1.64
Capsella bursa-pastoris(L.) Medik. ShepherdÕs purse WA Sp 0.39 0.34 0 0
Lepidium virginicumL. Virginia pepperweed WA Sp, F 0.51 0.33 0.34 0.34
Raphanus raphanistrumL. Wild radish WA Sp 1.80 N/A N/A N/A
Sinapis arvensisL. Wild mustard WA/B/P All 1.07 0.52 3.42 1.73
Thlaspi arvenseL. Field pennycress WA Sp 0.09 0.09 2.27 2.27
Convolvulaceae
Calystegia sepium(L.) R. Br. Hedge bindweed P Sp, Su 0.04 0.04 0 0
Fabaceae
Medicago lupulinaL. Black medic SA/WA All 0 0 0.13 0.12
Lamiaceae
Lamium purpureumL. Purple deadnettle WA Sp 0.04 0.04 0 0
Nepeta catariaL. Catnip P All 0 0 1.14 0.64
Malvaceae
Malva neglectaWallr. Common mallow WA/B/P All 0.54 0.50 0 0
Onagraceae
Oenothera biennisL. Evening primrose B All 0.03 0.03 0.24 0.24
Oxalidaceae
Oxalis strictaL. Yellow woodsorrel P Su, F 0.27 0.18 N/A N/A
Plantaginaceae
Plantago lanceolataL. Buckhorn plantain P All 0 0 0.04 0.04
Poaceae
Loliumsp. Ryegrass WA/B/P Su N/A N/A 0.60 N/A
Polygonaceae
Rumex crispusL.e Curly dock P All 0.02 0.02 0 0
Scrophulariaceae
Verbascum Thapsus L. Common mullein B All 0.04 0.04 0 0
Urticaceae
Urtica dioica L. Stinging nettle P All 0.01 0.01 0 0
Crop species Amaryllidaceae
Allium cepa L.e Onion B All 27.74 18.29 14.56 4.64
Fabaceae
Medicago sativa L. Alfalfa P Sp, Su 0.10 0.10 0 0
Poaceae
Avenasp. Oat, winter cover crop WA/C F N/A N/A 1.40 N/A
aMost common life history: SA⫽ summer-annual, WA ⫽ winter-annual, B ⫽ biennial, P ⫽ perennial, C ⫽ crop. Combinations indicate that
all three life-history variations are possible and are known to occur.
bPeriod when plants were sampled, indicating prevalence in the sampling location. Sp⫽ spring sampling (May, June), Su ⫽ summer sampling
(July, Aug.), F⫽ autumn sampling (Sept., Oct.), All ⫽ sampling through the duration of the onion growing season (May through Sept. and Oct.).
cPopulations were estimated to no. of larvae per plant per sampling date except on hedge bindweed, black medic, purple deadnettle, ryegrass,
common mallow, yellow woodsorrel, alfalfa, and the winter cover crop, where populations were estimated to no. of larvae per 0.0125 m2per
sampling date. 0⫽ no T. tabaci larvae were identiÞed (n ⱖ1 collected plant samples). N/A ⫽ no plants were sampled.
dStandard error of larvae per plant per sampling date or larvae per 0.0125 m2per sampling date. N/A⫽ insufÞcient no. of samples were
collected for calculation (0 or 1).
ence for onion (Doederlein and Sites 1993), T. tabaci larvae can exploit a wide variety of taxonomically diverse hosts. Still, 40% (11 of 25 species) of the re-productive hosts are members of Asteraceae (Þve spe-cies) and Brassicaceae (six spespe-cies), and 84% of T. tabaci larvae observed on weeds during this study were observed on members of these two taxa. Popu-lations of T. tabaci larvae were most abundant on four cruciferous species, yellow rocket, Barbarea vulgaris Ait. f., wild radish, Raphanus raphanistrum L., wild
mustard Sinapis arvensis L., and Þeld pennycress, Thlaspi arvenseL. Populations of T. tabaci larvae were greater than one larva per plant per sampling date on all four of these species, and populations on wild mustard were greater than two larvae per plant per sampling date during this study.
A number of factors are likely responsible for T. tabacito use a weed species as a host. Some of these factors include attraction to plant volatiles, nutrition, a relative absence of predators, or plant architecture
Fig. 1. Mean populations of T. tabaci larvae⫾SE on weed species known as hosts for IYSV in 2008 and 2009 in the Elba Muck onion growing region near Elba, NY.
Fig. 2. Temporal population densities of T. tabaci adults and larvae on onion and on four weed species known to be hosts for IYSV in 2008 and 2009 in the Elba Muck onion growing region near Elba, NY. Densities of adult T. tabaci on onion were not recorded in 2008. The left Y axis corresponds to populations on chicory, common burdock, curly dock, and dandelion, whereas the right Y axis corresponds to populations on onion.
Fig. 3. Estimated population densities⫾SE of weed spe-cies known as hosts for T. tabaci and IYSV in 2008 and 2009 in the Elba Muck onion growing region near Elba, NY.
that is favorable to thripsÕ cryptophilic and thigmo-tropic behavior (Lewis 1973). One of the more likely causes may be the abundance of Asteraceae and Bras-sicaceae species in the Elba Muck region, allowing T. tabacito exploit preferable species of these two highly varied taxa. Species in the Asteraceae and Brassi-caceae accounted for 22 and 9% of species in this region, respectively, and populations of species in these two families are among the highest in the region (Smith 2010).
In addition to onion, T. tabaci is also a major pest of cabbage crops worldwide (North and Shelton 1986, Shelton et al. 1998, Zezlina and Blazic 2003). The habit and architecture of cabbage and other cruciferous crops provide favorable habitats for T. tabaci as thig-mophilic animals; however, this is less true for weedy crucifers. This suggests that the relative preference for weedy Brassicaceae species over other surveyed weedy species may have less to do with physical char-acteristics than other factors such as attraction to plant volatiles, nutritional value, or a lack of predators.
Knowledge of when T. tabaci use plant species dur-ing the season can provide insight into which hosts may be important in the epidemiology of IYSV. In both years of this study, populations of T. tabaci larvae peaked on weeds from late-May to mid-June. Because onions are seeded and transplanted in April and May, they are too small to be colonized in May and early June in most years (E.A. Smith, personal observation). Although data were not collected in March or April in either year of this study, Larentzaki et al. (2007) found T. tabacion volunteer onions as early as March. This indicates that T. tabaci are active well before coloniz-ing onion crops, but are concentrated on other hosts (Fig. 2). During July and early August, densities of T. tabaci larvae decreased in weeds, but increased in onion crops. Late in the onion growing seasons of both years, larval densities on weeds experienced a second major peak from August through October. Larentzaki et al. (2007) observed adult T. tabaci on weeds as late as November. Onion crops are harvested from July through September, and T. tabaci adults originating in
onion may migrate from these Þelds into nearby weeds to feed and reproduce.
Weeds are largely ubiquitous in all onion growing regions. T. tabaci adults have been found on weeds bordering onion Þelds (Larentzaki et al. 2007), and at least 47 plant species in 19 families are hosts of IYSV (Table 1). Eleven of these species are non-Allium weed species typically found in New York onion eco-systems (E.A. Smith, personal observation), and four have tested positive for IYSV in New York (Hsu et al. 2011). Five of these 11 susceptible weed species are summer annual species in New York, but as stated previously, it is unlikely that summer-annuals contrib-ute signiÞcantly to the epidemiology of IYSV. IYSV and tospoviruses in general are not known to be seed-transmissible, and T. tabaci are not likely to preferentially colonize summer-annuals and pro-duce a viruliferous generation of T. tabaci until onions are harvested.
Winter-annuals, biennials, and perennials have the potential to be overwintering reservoirs for IYSV be-cause they could be infected in the fall and remain infected in the spring. Such weed species would be considered a green bridge for the virus to survive between onion-growing seasons and could be impor-tant sources of IYSV. In this study, the four weed species, common burdock, chicory, curly dock and dandelion, were candidates as potentially important IYSV sources because they were (1) hosts of IYSV, (2) hosts for T. tabaci larvae, and (3) had winter-annual, biennial, or perennial life histories.
In both years of this study, common burdock and dandelion supported the most T. tabaci larvae com-pared with chicory and curly dock. Prickly lettuce is a known host of IYSV (Sampangi et al. 2007) and T. tabaci larvae (Chatzivassiliou et al. 2007) and is a perennial weed known to occur in agro-ecosystems in New York. However, none of the 73 prickly lettuce plants sampled in this study were hosts of T. tabaci larvae and 0 of 60 plants previously sampled tested positive for IYSV (Hsu et al. 2011).
Populations of T. tabaci larvae per hectare on com-mon burdock and dandelion were estimated to be greater than on chicory and curly dock. These obser-vations suggest that the potential impacts of chicory and curly dock on IYSV epidemiology in New York may be relatively inconsequential when compared with the likely impacts that common burdock and dandelion may have on IYSV epidemiology. Common burdock has been observed with high populations of T. tabaci larvae, but its relative infrequency in the landscape suggests that its impact on the spread of IYSV may be more localized than that of dandelion, which is ubiquitous in the onion-producing landscape in the region.
The biennial life habit of common burdock may also affect IYSV persistence. Tomato spotted wilt virus (TSWV) (Bunyaviridae: Tospovirus) is known to per-sist in perennial plants for multiple years (Groves et al. 2002). While similar studies have not been conducted with IYSV, this suggests that IYSV may persist in dan-delion plants for multiple seasons. Conversely,
com-Fig. 4. Estimated abundance⫾SE of T. tabaci larvae per hectare on IYSV-positive weed species known as hosts for IYSV in 2008 and 2009 in the Elba Muck onion growing region near Elba, NY.
mon burdock survives only one winter before com-pleting its life cycle, and an infected burdock plant is likely to be a source of inoculum for only one onion growing season. This may be especially important for IYSV persistence after cool, wet growing seasons as thrips populations are known to be negatively im-pacted by such conditions (Liu 2004). In addition to these reasons, the compact rosette growth habit of dandelion plants offer T. tabaci a suitable habitat. Moreover, dandelion may have the greatest impact on IYSV epidemiology among the candidate weed species that are likely sources for IYSV.
Knowledge of rates of acquisition and transmission of IYSV by T. tabaci among the most abundant weed species is important. While transmission efÞciencies are unknown in weed species associated with T. tabaci and IYSV, Okazaki et al. (2009) evaluated acquisition and transmission rates of TSWV by Frankliniella oc-cidentals (Pergande) (Thysanoptera: Thripidae) in sticky chickweed, Cerastium glomeratum Thuill., black nightshade, Solanum nigrum L., common chickweed, Stellaria media(L.) Vill., and Galinsoga quadriradiata Cav. TSWV acquisition rates by F. occidentalis for each species were 85.4, 73.6, 72.6, and 35.6%, respectively, and transmission rates were 76.4, 60.9, 61.3, and 29.9%, respectively. Their results indicated that weed species vary in their abilities to be sources of TSWV. IYSV and TSWV are closely related, so this degree of variability in acquisition and transmission may be likely for T. tabaci and weed hosts of IYSV. Analogous studies involving T. tabaci transmission efÞciencies of IYSV from weeds to onions and vice versa will be important steps in identifying the most important weed sources of IYSV in the onion ecosystem.
More research on T. tabaci ecology and IYSV epi-demiology is needed to develop effective manage-ment strategies. Such research should include inves-tigation of the dispersal capabilities of T. tabaci and the effect of distance of weeds to onion crops on T. tabaci populations and IYSV incidence. Overwintering ca-pabilities of IYSV in speciÞc weeds as well as acqui-sition and transmission efÞciencies should also be in-vestigated. In particular, these Þndings will provide the information needed to determine if control of weeds in adjacent areas to onion Þelds is a worthwhile IYSV management strategy.
Acknowledgments
We thank the onion growers in Orleans and Genesee Counties, NY, who permitted sampling of weeds and onion thrips on their farms. This study was funded by grants from several agencies including the New York State Onion Re-search and Development Program; NY Farm Viability Insti-tute; USDA CSREES Hatch; and the Department of Ento-mology, Cornell University.
References Cited
de A´ vila, A. C., M.I.C.S. Gama, E. W. Kitajima, and W. Pereira. 1981. Um võ´rus do grupo vira-cabec¸a do
to-mateiro isolado de cebola (Allium cepa L.). (abstr.) Fi-topatol. Bras. 6: 525.
Balukiewicz, A., and S. Kryczynski. 2005. Tospoviruses in
chrysanthemum mother stock plants in Poland. Phyto-pathol. Pol. 37: 59 Ð 67.
Ben Moussa, A., M. Marrakchi, and M. Makni. 2005.
Char-acterisation of Tospovirus in vegetable crops in Tunisia. Infect. Genet. Evol. 5: 312Ð322.
Chatzivassiliou, E. K., D. Peters, and N. I. Katis. 2007. The
role of weeds in the spread of tomato spotted wilt virus by
Thrips tabaci (Thysanoptera: Thripidae) in tobacco crops. J. Phytopathol. 155: 699 Ð705.
Childers, C. C. 1997. Feeding and oviposition injuries to
plants, pp. 505Ð537. In T. Lewis (ed.), Thrips as Crops Pests. CAB International, Oxon, NY.
Corteˆs, I., I. C. Livieratos, A. Derks, D. Peters, and R. Kormelink. 1998. Molecular and serological
character-ization of iris yellow spot virus, a new and distinct to-spovirus species. Phytopathology 88: 1276 Ð1282.
Cosmi, T., E. Marchesini, and G. Martini. 2003. Presence
and spread of tospovirus and thrip vectors in Veneto. Inf. Agrar. 59: 69 Ð72.
Coutts, B.A., L. A. McMichael, L. Tesoriero, B. C. Rodoni, C. R. Wilson, A. J. Wilson, D. M. Persley, and R.A.C. Jones. 2003. Iris yellow spot virus found infecting onions
in three Australian states. Australas Plant Pathol. 32: 555Ð 557.
Doederlein, T. A., and R. W. Sites. 1993. Host plant
prefer-ences of Frankliniella occidentalis and Thrips tabaci (Thy-sanoptera: Thripidae) for onions and associated weeds on the Southern High Plains. J. Econ. Entomol. 86: 1706 Ð 1713.
Doi, M., S. Zen, M. Okuda, H. Nakamura, K. Kato, and K. Hanada. 2003. Leaf necrosis disease of lisianthus
(Eus-toma grandiflorum) caused by Iris yellow spot virus. Jpn. J. Phytopathol. 69: 181Ð188.
Evans, C. K., S. Bag, E. Frank, J. R. Reeve, D. Drost, and H. R. Pappu. 2009a. Natural infection of Iris yellow spot virus
in twoscale saltbush (Atriplex micrantha) growing in Utah. Plant Dis. 93: 430.
Evans, C. K., S. Bag, E. Frank, J. R. Reeve, C. Ransom, D. Drost, and H. R. Pappu. 2009b. Green foxtail (Setaria
viridis), a naturally infected grass host of Iris yellow spot virus in Utah. Plant Dis. 93: 670.
Fournier, F., G. Boivin, and R. K. Stewart. 1995. Effect of
Thrips tabaci(Thysanoptera: Thripidae) on yellow onion yields and economic thresholds for its management. J. Econ. Entomol. 88: 1401Ð1407.
Gent, D. H., H. F. Schwartz, and R. Khosla. 2004.
Distribu-tion and incidence of Iris yellow spot virus in Colorado and its relation to onion plant population and yield. Plant Dis. 88: 446 Ð 452.
Gent, D. H., L. J. du Toit, S. F. Fichtner, S. K. Mohan, H. R. Pappu, and H. F. Schwartz. 2006. Iris yellow spot virus:
an emerging threat to onion bulb and seed production. Plant Dis. 90: 1468 Ð1480.
Gera, A., A. Kritzman, H. Beckelman, J. Cohen, and B. Rac-cah. 2002. Detection of Iris yellow spot virus in lisianthus.
Acta Hortic. 568: 43Ð 49.
Gera, A., A. Kritzman, J. Cohen, and B. Raccah. 1998.
To-spoviruses infecting bulb crops in Israel. In D. Peters and R. Goldbach (eds.), Recent Progress in Tospovirus and Thrips Research, pp. 86 Ð 87. Fourth International Sym-posium on Tospoviruses and Thrips in Floral and Vege-table Crops, Wagengingen, The Netherlands.
Ghotbi, T., N. Shahraeen, and S. Winter. 2005. Occurrence
of tospoviruses in ornamental and weed species in Markazi and Tehran provinces in Iran. Plant Dis. 89: 425Ð 429.
Groves, R. L., J. F. Walgenbach, J. W. Moyer, and G. G. Kennedy. 2002. The role of weed hosts and tobacco
thrips, Frankliniella fusca, in the epidemiology of Tomato
spotted wilt virus.Plant Dis. 86: 573Ð582.
Hall, J. M., K. Mohan, E. A. Knott, and J. W. Moyer. 1993.
Tospoviruses associated with scape blight of onion
(Al-lium cepa) seed crops in Idaho. Plant Dis. 77: 952.
Hoepting, C. A., H. F. Schwartz, and H. R. Pappu. 2007. First
report of Iris yellow spot virus on onion in New York. Plant Dis. 91: 327.
Hsu, C. L., C. A. Hoepting, M. Fuchs, A. M. Shelton, and B. A. Nault. 2010. Temporal dynamics of Iris yellow spot virus
and its vector, Thrips tabaci(Thysanoptera: Thripidae), in seeded and transplanted onion Þelds. Environ. Entomol. 39: 266 Ð277.
Hsu, C. L., C. A. Hoepting, M. Fuchs, E. A. Smith, and B. A. Nault. 2011. Sources of Iris yellow spot virus in New
York. Plant Dis. (in press).
Jones, D. R. 2005. Plant viruses transmitted by thrips. Eur. J.
Plant Pathol. 113: 119 Ð157.
Kritzman, A., H. Beckelman, S. Alexandrov, J. Cohen, M. Lampel, M. Zeidan, B. Raccah, and A. Gera. 2000.
Li-sianthus leaf necrosis: a new disease of liLi-sianthus caused by Iris yellow spot virus. Plant Dis. 84: 1185Ð1189.
Kritzman, A., M. Lampel, B. Raccah, and A. Gera. 2001.
Distribution and transmission of Iris yellow spot virus. Plant Dis. 85: 838 Ð 842.
Larentzaki, E., A. M. Shelton, F. R. Musser, B. A. Nault, and J. Plate. 2007. Overwintering locations and hosts for
on-ion thrips (Thysanoptera: Thripidae) in the onon-ion crop-ping ecosystem in New York. J. Econ. Entomol. 100: 1194 Ð1200.
Lewis, T. 1973. Thrips: their biology, ecology and economic
importance. Academic, London, United Kingdom.
Lewis, T. (ed). 1997. Thrips as crop pests. CAB
Interna-tional, New York.
Liu, T. X. 2004. Seasonal population dynamics, life stage
composition of Thrips tabaci (Thysanoptera: Thripidae), and predacious natural enemies on onions in south Texas. Southwest Entomol. 29: 127Ð135.
Mohan, S. K., E. A. Knott, T. C. Allen Jr., and D. O. Wilson Jr. 1991. Evidence for association of Tomato spotted wilt
viruswith a scape blight disease of onion seed crops in Idaho and Oregon, pp. 26 Ð31. In Proceedings of the Na-tional Onion Research Conference, Savannah, GA.
Moritz, G., D. Morris, and L. Mound. 2001. ThripsID: pest
thrips of the world: an introduction to identiÞcation and classiÞcation of pest thrips of the world. CSIRO Publish-ing, Collingwood, VIC, Australia. [CD-ROM].
Nault, B. A., M. Fuchs, C. Hsu, E. A. Smith, and A. M. Shelton. 2008. Potential sources of IYSV, relationship between
IYSV and onion thrips and thrips control in New York, pp. 9 Ð12. In Proceedings of the Wisconsin Muck Crops Re-search Update 2008 Annual Meeting, 4 March, 2008, Uni-versity of WisconsinÐMadison, Madison, WI.
Nault, B. A., A. M. Shelton, J. L. Gangloff-Kaufmann, M. E. Clark, J. L. Werren, J. C. Cabrera-La Rosa, and G. G. Kennedy. 2006. Reproductive modes in onion thrips
(Thysanoptera: Thripidae) populations from New York onion Þelds. Environ. Entomol. 35: 1264 Ð1271.
Nischwitz, C., R. D. Gitaitis, S. W. Mullis, A. S. Csinos, D. B. Langston, and A. N. Sparks. 2007. First report of Iris
yellow spot virusin spiny sowthistle (Sonchus asper) in the United States. Plant Dis. 91: 1518.
North, R. C., and A. M. Shelton. 1986. Overwintering of the
onion thrips, Thrips tabaci (Thysanoptera: Thripidae), in New York. Environ. Entomol. 15: 695Ð 699.
Okazaki, S., M. Okuda, and T. Sakurai. 2009. Evaluation of
weed species as Tomato spotted wilt virus (Bunyaviridae: Tospovirus) acquisition sources for Frankliniella
occiden-talis(Pergande) (Thysanoptera: Thripidae). Jpn. J. Appl. Entomol. Z. 53: 181Ð184.
Okuda, M., and K. Hanada. 2001. RT-PCR for detecting Þve
distinct Tospovirus species using degenerate primers and dsRNA template. J. Virol. Meth. 96: 149 Ð156.
Pappu, H. R., B. C. Hellier, and F. M. Dugan. 2006. Wild
Alliumspp. as natural hosts of Iris yellow spot virus. Plant Dis. 90: 378.
Pappu, H. R., and M. Matheron. 2008. Characterization of
Iris yellow spot virusfrom onion in Arizona. Plant Health Prog. (dol: 10.1094/PHP-2008 Ð 0711-01-BR).
Pozzer, L., I. C. Bezerra, R. Kormelink, M. Prins, D. Peters, R. de O. Resende, and A. C. de A´ vila. 1999.
Character-ization of a tospovirus isolate of Iris yellow spot virus associated with a disease in onion Þelds in Brazil. Plant Dis. 83: 345Ð350.
Robe`ne-Soustrade, I., B. Hostachy, M. Roux-Cuvelier, J. Mi-natchy, M. He´dont, R. Pallas, A. Couteau, N. Cassam, and G. Wuster. 2006. First report of Iris yellow spot virus in
onion bulb- and seed-production Þelds in Re´union Island. Plant Pathol. 55: 288.
Sampangi, R. K., and S. K. Mohan. 2007. IdentiÞcation of
New Alternative Weed Hosts for Iris yellow spot virus in the PaciÞc Northwest. Plant Dis. 91: 1683.
Shelton, A. M., W. T. Wilsey, and M. A. Schmaedick. 1998.
Management of onion thrips on cabbage using plant re-sistance and insecticides. J. Econ. Entomol. 81: 1062Ð1067.
Smith, E. A. 2010. Identifying weed species hosts for onion
thrips (Thrips tabaci Lindeman) and their potential as sources of Iris yellow spot virus (Bunyaviridae: Tospovi-rus) in New York onion Þelds. M.S. Thesis, Cornell Uni-versity, Ithaca, NY.
Ullman, D. E., J. J. Cho, R.F.L. Mau, D. M. Westcott, and D. M. Custer. 1992. A midgut barrier to tomato spotted
wilt virus acquisition by adult western ßower thrips. Phy-topathology 82: 1333Ð1342.
Whitfield, A. E., D. E. Ullman, and T. L. German. 2005.
Tospovirus-Thripsinteractions. Annu. Rev. Phytopathol. 43: 459 Ð 489.
Wijkamp, I., J. Van Lent, R. Kormelink, R. Goldbach, D. Peters. 1993. Multiplication of tomato spotted wilt virus
in its insect vector, Frankliniella occidentalis. J. Gen. Virol. 74: 341Ð349.
Zezlina, I., and M. Blazic. 2003. Testing the efÞcacy of
dif-ferent insecticides to control onion thrips (Thrips tabaci Lindeman, Thysanoptera, Thripidae) in onion crops. Commun. Agric. Appl. Biol. Sci. 68: 287Ð290.
Appendix 1. Winter-annual, biennial, and perennial weed species that were not determined as hosts of T. tabaci larvae. Weeds were sampled from the Elba Muck onion growing region near Elba, NY, in 2008 and 2009
Family
Common name Habita
Species Apiaceae
Daucus carotaL. Wild carrot B
Pastinaca sativaL. Wild parsnip B
Apocynaceae
Apocynum cannabinumL. Hemp dogbane P
Asteraceae
Achillea millefoliumL. Common yarrow P
Centaurea stoebeL. Spotted knapweed P
Cirsium vulgare(Savi) Ten. Bull thistle B
Conyza canadensis(L.) Cronq. Horseweed P
Erigeron philadelphicusL. Philadelphia ßeabane B/P
Hieracium canadenseMichx Canadian hawkweed P
Hieracium caespitosumDumort Yellow hawkweed P
Hypochaeris radicataL. Catsear P
Lactuca serriolaL. Prickly lettuce SA/WA/P
Senecio vulgarisL. Common groundsel SA/WA
Sonchus arvensisL. Perennial sowthistle P
Symphyotrichum novae-angliae(L.) Nesom New england aster P
Brassicaceae
Alliaria petiolata(Bieb.) Cavara & Grande Garlic mustard B
Descurainia sophia(L.) Webb ex Prantl Flixweed WA/SA
Hesperis matronalisL. DameÕs rocket B/P
Rorippa palustris(L.) Bess. Marsh yellow cress SA/WA/P
Caryophyllaceae
Cerastium fontanumBaumg. Mouse ear chickweed P
Saponaria officinalisL. Bouncing bet P
Silene latifoliaPoir. White campion P
Dipsacaceae
Dipsacus fullonumL. Common teasel B
Fabaceae
Lotus corniculatusL. Birdsfoot trefoil P
Trifolium pratenseL. Red clover P
Trifolium repensL. White clover P
Lamiaceae
Lamium amplexicauleL. Henbit WA
Oxalidaceae
Oxalis strictaL. Woodsorrel P
Phytolaccaceae
Phytolacca americanaL. Am. pokeweed P
Plantaginaceae
Plantago majorL. Broadleaf plantain P
Poaceae
Bromus inermisLeyss. Smooth brome P
Bromus secalinusL. Rye brome (cheat) WA
Bromus tectorumL. Downy brome SA/WA
Dactylis glomerataL. Orchardgrass P
Elytrigia repens(L.) Gould Quackgrass P
Holcus lanatusL. Common velvetgrass P
Lolium multiflorumLam. Italian ryegrass WA/B/P
Phleum pratenseL. Timothy P
Phragmites australis(Cav.) Trin. Ex Steud. Common reed P
Poa annuaL. Annual bluegrass WA
Poa compressaL. Canada bluegrass P
Ranunculaceae
Ranunculus bulbosusL. Bulbous buttercup P
Rosaceae
Potentilla norvegicaL. Rough cinquefoil SA/WA/B/P
Potentilla rectaL. Sulphur cinquefoil P
Rubusspp. (R. occidentalis, etc.) Wild raspberry B/P
Vitaceae
Parthenocissus quinquefolia(L.) Planch. Virginia creeper P
Vitis labruscaL. Fox grape P
aSA⫽ summer-annual, WA ⫽ winter-annual, B ⫽ biennial, P ⫽ perennial, and combinations indicate that multiple life histories are known