Phosphorylation/Dephosphorylation of the Repressor MDBP-2-H1 Selectively Affects the Level of Transcription from a Methylated Promoter in Vitro
6
0
0
Texte intégral
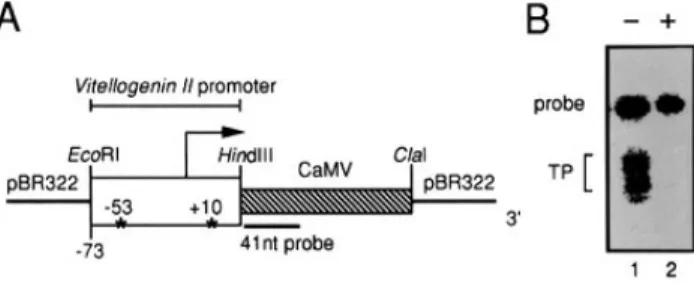
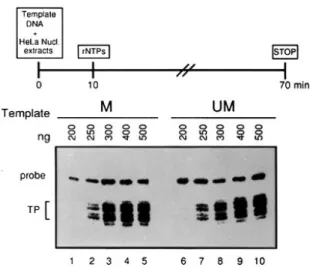
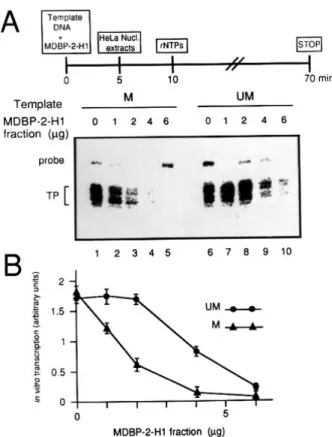
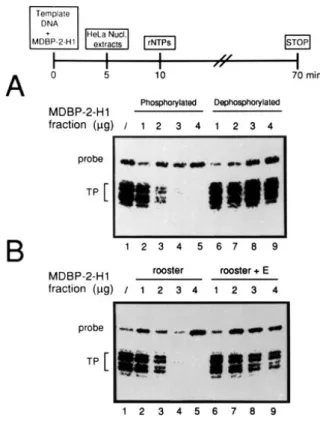
Figure
Documents relatifs