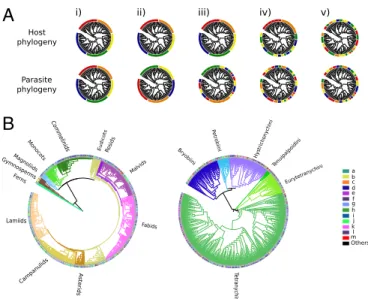
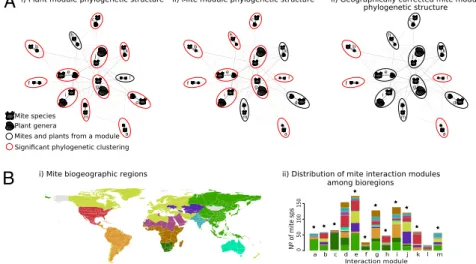
Geography and major host evolutionary transitions shape the resource use of plant parasites
Texte intégral
Figure

Documents relatifs
3) What leadership skills and behavior relate to employee satisfaction, motivation for performance, group performance, and organizational performance? 4) Does the leader's
Assessment of an iconic-geometric nonlinear registration method for deep brain stimulation (DBS) planning..
Pour cela il faut que l'air injecté soit soufflé à une vitesse suffisante pour entraîner l'air de la pièce tout en produisant une circulation secondaire, des quantités
Les limites de détection des biosenseurs bactériens couplés à des transducteurs électrochimiques peuvent être extrêmement basses et
The P-CuReD is a method with linear complexity for wavefront reconstruction from pyramid sensor data which employs the CuReD algorithm and a data preprocessing step converting
axonopodis inferred from the analysis of seven housekeeping genes from 131 strains estimated on overall data set (a), each of the 6 groups (b), and pathovars (c).. Values for
• Ralstonia solanacearum : responsible for heavy losses on cash and subsistence crops throughout tropical and temperate areas (bacterial wilt, potato brown rot, Moko disease). •
Le Conseil scientifique, par l’intermédiaire de son président, collabore à l’installation d’une serre à Welwitschia mirabilis à Porrentruy, en partenariat étroit avec le
![[PDF] Formation sur les Notions de Base d’OPenOffice Calc | Cours informatique](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)