B cells with regulatory properties in transplantation tolerance
Texte intégral
Figure
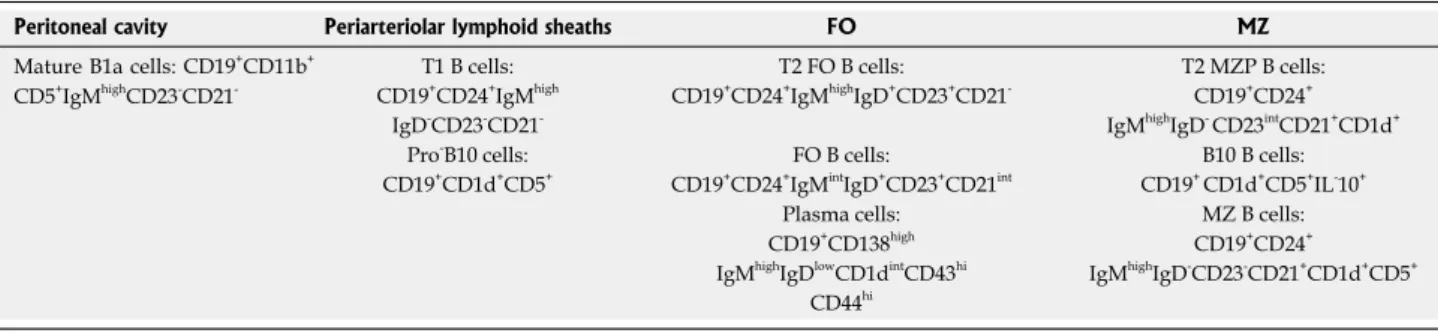

Documents relatifs
However, immunotherapies, such as the anti-CD20 B cell-depleting antibody, can generate deleteri- ous effects on the transplant, likely due to the deletion of beneficial
We have shown that highly confined surface magnon polaritons, such as those on anti- ferromagnetic materials, could speed up magnetic transitions by more than 10 orders of
f, z-projection (0.6 mm) magnifications of C1/C2 centrosomal centrioles (white arrows) and deuterosome (flower). g, h, Lengths and widths of procentrioles from cells at the halo,
(A) Proportions of total FOXP3 + Tregs (gray bars); B) na ï ve Tregs (blue bars); C) and eTregs (green bars) in the blood of study subjects, by clinical status (hepatic fibrosis
Binding of antigens to cell surface was evaluated using total or negatively selected B cells from splenocytes of naïve mice.. Cells were incubated 1 hr on ice alone or in the
Abbreviations: ADC, adenocarcinoma; Breg, regulatory B cell; CAF, cancer- associated fibroblast; CM, central-memory; DC, dendritic cell; EM, effector- memory; FDR, false discovery
Alors que le pourcentage de Treg était similaire dans le thymus des souris saines et des souris déficientes en WASp, le nombre de Treg périphérique était
For example, with 1 million international unit (MIU) IL-2 ‘‘virtual’’ injections every 3 days, Treg cells were expected to remain stable with a delta variation of %Treg cells
