Dynamique holocène de la végétation et des feux d’un
paysage contemporain dominé par des espèces de
début de succession au sein de la Pessière à mousses
de l’Ouest du Québec
Mémoire
Maxime Asselin
Maîtrise en sciences géographiques
Maître en sciences géographiques (M.Sc.Géogr.)
Québec, Canada
Dynamique holocène de la végétation et des feux d’un
paysage contemporain dominé par des espèces de
début de succession au sein de la Pessière à mousses
de l’Ouest du Québec
Mémoire
Maxime Asselin
Sous la direction de :
iii
Résumé
Dans le contexte d’études visant à établir des liens entre les paysages forestiers naturels actuels et la façon dont ils se sont transformés au cours de l’Holocène (variabilité naturelle), il est important de définir des stratégies sylvicoles qui répondent aux critères de la mise en place de l’aménagement écosystémique. Au sein du domaine bioclimatique de la pessière à mousses de l’Ouest du Québec, certains paysages forestiers sont aujourd’hui caractérisés par de vastes peuplements de début de succession dominés par le peuplier faux-tremble et le pin gris. Les objectifs de cette étude étaient 1) de reconstituer l’histoire postglaciaire de la végétation et des feux d’un secteur dominé par de tels peuplements, et 2) de préciser leur variabilité naturelle plurimillénaire. À cette fin, les paysages forestiers d’un territoire couvrant une superficie de 4000 km2 dans la région de la plaine du lac Matagami ont d’abord été caractérisés. Dans un deuxième temps, l’histoire de la végétation et des feux a été reconstituée à l’aide de l’analyse pollinique et anthracologique de deux tourbières. Les lieux de prélèvement des sédiments ont été choisis en fonction d’obtenir un enregistrement local et extra-local de l’histoire des forêts jouxtant chacune des tourbières. Les résultats montrent que la majeure partie de l’Holocène (7000 à 1000 ans A.A.) a été caractérisée des feux peu fréquents qui ont permis le maintien de paysages dominés par la pessière noire à sapin baumier. Au cours des 1000 dernières années, des feux plus fréquents et sans doute plus sévères et de plus grandes étendues ont provoqué une diminution de l’abondance du sapin et une augmentation des espèces de début de succession. Les feux du dernier millénaire seraient ainsi à l’origine des actuels peuplements de début de succession qui se maintiendraient selon une dynamique de récurrence.
Table des Matières
Résumé ... iii
Table des Matières ... iv
Liste des tableaux ... v
Liste des figures ... vi
Liste des abréviations ... vii
Abréviations utilisées dans le texte ... vii
Abréviations des espèces végétales ... vii
Remerciements ... viii
Introduction ... 1
Le territoire d’étude ... 4
Méthodes ... 6
Les paysages forestiers contemporains ... 6
Les tourbières échantillonnées ... 6
La forêt contemporaine ceinturant les deux tourbières à l’étude ... 8
Stratigraphie ... 8
La reconstitution de l’historique des feux ... 9
La reconstitution de l’histoire de la végétation ... 10
Datations radiochronologiques ... 11
Résultats ... 12
Les paysages forestiers contemporains de la région d’étude ... 12
La forêt contemporaine ceinturant les deux tourbières à l’étude ... 15
Reconstitutions paléoenvironnementales ... 17
Stratigraphie et données chronologiques ... 17
Reconstitution de l’historique des feux de forêt ... 20
Reconstitution de l’histoire de la végétation ... 20
Discussion et conclusion ... 26
La dynamique plurimillénaire des paysages ... 26
La dynamique contemporaine des paysages ... 31
v
Liste des tableaux
Tableau 1 : Données radiochronologiques des tourbières Aspen et Shadow. Les dates exclues dans des modèles âge-profondeur sont en caractère italique. ... 16 Tableau 2 : Représentation pollinique (espèces selectionnés) de la végétation actuelle de deux
Liste des figures
Figure 1 : Localisation du territoire d’étude dans la région écologique de la plaine du lac Matagami. Les domaines bioclimatiques sont aussi illustrés ... 4 Figure 2 : Photos terrestres (haut) et photographies aériennes (bas; échelle 1: 20 000) des forêts
entourant les tourbières Aspen et Shadow. Le site d’échantillonnage de la carotte sédimentaire est représenté par le carré noir. Crédit photo: Pierre Grondin ... 7 Figure 3 : Délimitation des paysages forestiers du territoire d’étude (P1-P4). A) Altidude (m). B)
Répartition des dépôts de surface. C) Répartition des peuplements de drainage mésique. D) Répartition des pessières noires sur sol minéral et des pessières noires sur sol organique. E) Répartition des peuplements forestiers dominés par des tremblaies et des pinèdes grises. F) Répartition des peuplements dont la période d’origine est 1820 et 1910. Les deux tourbières à l’étude se situent dans le paysage P4b (A) qui est le mieux pourvu en espèces de début de succession. ... 14 Figure 4 : Caractéristiques des forêts ceinturant les deux tourbières à l’étude, en regard du
nombre de tiges par espèce, de la surface terrière et de la régénération coefficient de distribution. ... 16 Figure 5 : Stratigraphie et modèles âge-profondeur des carottes sédimentaires récoltées aux
tourbières Aspen et Shadow. ... 19 Figure 6 : Diagramme pollinique (pourcentages; taxons polliniques choisis) et de l’anthracomasse
de la tourbière Aspen. Les courbes polliniques non remplies représentent une exagération de 10 ×. ... 23 Figure 7 : Diagramme pollinique (pourcentages; taxons polliniques choisis) et de l’anthracomasse
de la tourbière Shadow. Les courbes polliniques non remplies représentent une exagération de 10 ×. ... 24 Figure 8 : Résultat de l’analyse en composantes principales (ACP) des assemblages polliniques
combinés des sites Aspen et Shadow. a) Ordination stratigraphique de la note des échantillons sur le premier axe. Les résultats sont présentés selon une échelle de temps. La première valeur propre (λ1) et le pourcentage de la variance sont donnés. b) Saturations des principaux taxons sur le premier axe ACP. ... 25 Figure 9 : Territoire auquel les résultats de la présente étude pourraientêtre potentiellement
extrapolés. Ce territoire correspond à la portion sud de la région écologique de la plaine argileuse de l'Abitibi (domaine bioclimatique de la pessière à mousses ; région 6a) et la plaine de l'Abitibi (domaine de la sapinière à bouleau blanc ; région écologique 5a). Les peuplements forestiers dominés par Populus tremuloides sur argile mésique-subhydrique sont illustrés en vert. La zone d'étude où furent menées les analyses paléoécologiques est illustrée par le rectangle gris et les deux tourbières étudiées par les deux points orange. ... 30
vii
Liste des abréviations
Abréviations utilisées dans le texte
ACP : analyse en composantes principales
Années A.A. : années avant aujourd’hui (années étalonnées) CDR : coefficient de distribution de la régénération
ha : hectare mg : milligramme
RDA : analyse de redondance
SMA : spectrométrie de masse par accélérateur ZAP : zone d’assemblage pollinique
Abréviations des espèces végétales
PMA : Picea mariana PBA : Pinus banksiana ABA : Abies balsamea BPA : Betula papyrifera
AUC : Alnus viridis subsp. crispa AUR : Alnus incana subsp. rugosa AUX : Alnus
SAL : Salix
TUB : Tubuliflorae
Remerciements
Je voudrais d’abord remercier ma famille, ainsi que ma conjointe qui, chacun à leur manière, m’ont supporté tout au long de mes études. Mes remerciements s’adressent ensuite à mon directeur de recherche Martin Lavoie. Il a toujours été présent afin de me faire cheminer dans la bonne direction durant mes trois années. Une mention spéciale à Pierre Grondin, chercheur au ministère des Forêts, de la faune et des Parcs du Québec, qui fut pour moi un mentor exceptionnel durant la totalité du projet. Il a été présent dans chaque facette de cette recherche afin de m’aider à cheminer et faire évoluer mes idées. Du terrain à la rédaction, Pierre a continuellement repoussé mes limites personnelles pour m’amener à proposer un projet de recherche complet et original. J’ai aussi bénéficié de l’expertise indispensable de Ann Delwaide en dendrochronologie, de Bianca Fréchette avec mes analyses numériques et de Myosotis Bourgon Desroches avec mes analyses polliniques. La réalisation de ce projet de recherche n’aurait pas été possible sans l’aide financière du FRQNT obtenu par Yves Bergeron (2013-2016) et d’une bourse Mitacs. Pour terminer, j’aimerais aussi remercier Véronique Poirier et Jean Noël de l’équipe de géomatique de la direction de la recherche forestière du MFFP pour leur collaboration et leur aide.
1
Introduction
Les paysages forestiers boréaux se développent sous l’influence combinée du climat, des variables du milieu physique (dépôts de surface, conditions de drainage, topographie, etc.) et des perturbations écologiques naturelles et anthropiques (Girardin et al. 2008 ; Ali et al. 2009 ; Mansuy et. 2010 ; Oris et al. 2013). Ainsi, les paysages naturels contemporains consistent en des peuplements d’âge, de composition et de structure variés issus de l’effet des feux qui sont de superficies et de sévérités variables et qui affectent des milieux physiques aux caractéristiques diverses (Heinselman 1973 ; White 1979 ; Pickett et White 1985 ; Bergeron et al. 2001). Cette hétérogénéité s’exprime à différentes échelles spatiales. Les paysages de la zone boréale sont considérés comme distincts de ceux de la zone tempérée. Ces zones sont ensuite subdivisées en unités territoriales plus petites, celles-ci étant mises en évidence dans les systèmes de classification écologique (Saucier et al. 2009). Au final, les paysages forment des portions de territoire de superficie variable qui expriment des liens entre la végétation, les variables du milieu physique et les perturbations écologiques (Grondin et al. 2014).
À l’intérieur des paysages forestiers boréaux, les espèces arborescentes se répartissent le long d’une toposéquence de milieux physiques, depuis les sols minces et bien drainés vers les sols organiques épais aux conditions hydriques. Ainsi, dans le nord-est de l’Amérique du Nord, le pin gris (Pinus banksiana) et l’épinette noire (Picea mariana) sont deux espèces bien adaptées aux stations caractérisées par des sols minces et sableux où les feux surviennent de façon récurrente. L’épinette noire et le mélèze (Larix laricina) caractérisent les stations hydriques où les feux surviennent beaucoup plus rarement. Pour leur part, les sols épais et de drainage modéré supportent une végétation relativement plus exigeante en ce qui a trait au régime nutritif comme le peuplier faux tremble (Populus tremuloides), le bouleau à papier (Betula papyrifera) et le sapin baumier (Abies balsamea). Sous l’effet des feux, une dynamique forestière propre à chacun de ces milieux physiques se réalise. Par exemple, le pin gris occupera d’abord les sites rocheux ou sableux alors que ce sont les feuillus de lumière (P. tremuloides, B. papyrifera) qui coloniseront les milieux plus riches. Avec le temps, les espèces de fin de succession (P. mariana, A. balsamea) gagneront en importance pour ainsi définir une dynamique successionelle (Frelich 2002). Toutefois, il arrive que les feux soient si rapprochés dans le temps que des cohortes de début de succession se succèdent, définissant ainsi une dynamique cyclique. Dans de telles situations, la forêt de début de succession peut être considérée comme à l’équilibre avec le climat et les régimes de perturbation.
Chacun des paysages spatialement distincts que l’on observe aujourd’hui résulte d’une dynamique temporelle qui fut dictée, entre autres, par les changements survenus au sein du climat et du régime des perturbations. Cette dynamique temporelle peut, dans certains cas, être de l’ordre de plusieurs millénaires. Dans le domaine bioclimatique de la pessière à mousses de l’Ouest du Québec (Canada), le climat plus chaud de l’optimum climatique de l’Holocène (~8000 à ~4000 ans avant aujourd’hui [A.A.]) et des feux sans doute moins fréquents qu’actuellement auraient notamment favorisé le développement du sapin baumier et du thuya (Thuya occidentalis) (Liu 1990 ; Richard 1995 ; Carcaillet et al. 2001). Des feux plus fréquents et un climat plus frais au cours des derniers millénaires (Néoglaciaire) auraient ensuite eu pour conséquence de fragmenter l’aire de répartition de ces espèces. Aujourd’hui, le sapin est principalement cantonné sur les versants de collines ainsi qu’à des bordures de cours d’eau qui semblent agir comme des refuges contre le feu. Pour sa part, le thuya est restreint à des populations marginales jouxtant quelques lacs. En contrepartie, on observe une augmentation de la représentation pollinique du pin gris dans plusieurs diagrammes polliniques (Richard 1979 ; Carcaillet et al. 2001 ; Fréchette et al. 2016).
Dans le contexte d’études visant à établir des liens entre les paysages naturels actuels et la façon dont ils se sont transformés au cours des millénaires (variabilité naturelle), il est important de définir des stratégies sylvicoles qui répondent aux critères de la mise en place de l’aménagement écosystémique, c’est-à-dire d’une réduction des écarts entre les paysages naturels et ceux aménagés (Kuuluvainen 2002 ; Gauthier et al. 2008). Actuellement, au Québec, les états de référence pour l’aménagement écosystémique sont définis en fonction des proportions de vieux peuplements, des types de composition forestière (feuillus, mixtes, conifériens) et les cycles de feu contemporains (˂300 ans) (Boucher et al. 2012). Le régime forestier en vigueur appuie cette approche et soutient l’idée du maintien à long terme des multiples fonctions de l’écosystème et de leur biodiversité (Gauthier et al. 2008) dans un contexte de changements climatiques (Logan et al. 2012). Notre compréhension de la dynamique naturelle des paysages repose principalement sur ces états de référence contemporains. Il s’agit maintenant d’étendre ces connaissances sur une plus longue échelle temporelle par l’entremise d’études paléoécologiques.
3
La présente étude vise à 1) reconstituer l’histoire postglaciaire de végétation et des feux et 2) de bonifier nos connaissances de la dynamique forestière contemporaine par un éclairage de la paléoécologie pour un territoire du domaine de la pessière à mousses de l’Ouest du Québec où abondent de vastes peuplements de début de succession caractérisés par le peuplier faux-tremble et le pin gris. Nous posons l’hypothèse que la configuration actuelle de la végétation s’est développée en lien avec une fréquence relativement élevée de feux survenus lors des derniers millénaires, et que cette configuration se serait maintenue jusqu’à nos par une dynamique de récurrence. Afin de répondre aux objectifs et de tester cette hypothèse, nous avons caractérisé les paysages forestiers contemporains d’un territoire couvrant une superficie de 4000 km2 et reconstitué l’histoire à long terme de la végétation et des feux à l’aide d’une analyse paléoécologique (grains de pollen, charbons de bois macroscopiques) des sédiments organiques de deux tourbières situées à quelques kilomètres l’une de l’autre. Les lieux de prélèvement des carottes sédimentaires ont été choisis en fonction d’obtenir un enregistrement local et extra-local de l’histoire de la végétation et des feux des forêts jouxtant chacune des tourbières, tout en essayant d’obtenir la plus longue séquence temporelle possible. Au final, nous cherchons à préciser la variabilité naturelle plurimillénaire d’un paysage contemporain dominé par les espèces de début de succession et à situer les connaissances acquises dans le contexte de l’aménagement écosystémique.
Le territoire d’étude
Le territoire à l’étude couvre une superficie de 4000 km2 au sein du domaine bioclimatique de la pessière à mousses de l’Ouest du Québec, au nord du village de Matagami (Fig. 1). Il appartient à la région écologique de la plaine argileuse du lac Matagami (Saucier et al. 2009). Cette région se caractérise par la présence d’épais dépôts glaciolacustres argileux mis en place par le lac proglaciaire Ojibway suite au retrait de l’inlandsis Laurentidien il y a environ 8000 ans (Vincent et Hardy 1977; Veillette 1994). La température moyenne annuelle se situe entre -1 et -2°C. Le mois le plus chaud est juillet (moyenne : 16,8°C) et janvier le plus froid (moyenne : -20°C). Les précipitations totales annuelles varient entre 800 et 1000 mm, dont 25-30 % tombent sous forme de neige (station météorologique de Matagami; Environnement Canada 2013).
Figure 1 : Localisation du territoire d’étude dans la région écologique de la plaine du lac Matagami. Les domaines
5
Le couvert forestier se compose principalement d’espèces boréales, notamment Picea mariana,
Pinus banksiana, Populus tremuloides, Abies balsamea et Betula papyrifera. L’épinette noire
domine les paysages et forme des peuplements possédant une diversité de densité, de hauteur et de classes d’âge (Robitaille et Saucier 1998). Le feu est la perturbation écologique la plus importante. Le cycle de feu était de 100 ans avant l’année 1850 et de 130 ans entre les années 1850 et 1920; il est présentement estimé à 400 ans (Bergeron et al. 2001). Deux grandes voies successionnelles caractérisent la dynamique forestière après feu. La première est typique des sites les plus bas en altitude, mal drainés et où les tourbières abondent. Dans ces endroits, bon nombre de pessières noires sur sol minéral évoluent, en fonction du temps écoulé depuis le dernier feu, vers des pessières noires paludifiées (Lecomte et al. 2005 ; Simard et al. 2007 ; Belleau et al. 2011). La seconde voie successionnelle caractérise les sites plus hauts en altitude, mieux drainés et où les tourbières sont plus rares. Dans ces secteurs, les pessières noires ont moins tendance à se paludifier et les tremblaies à pin gris et épinette noire évoluent vers des pessières noires à sapin. En raison de la fréquence et de la superficie des feux contemporains, ces forêts de fin de succession sont actuellement rares dans la région.
Méthodes
Les paysages forestiers contemporains
La première étape fut de délimiter des entités de végétation afin d’orienter le choix des tourbières à échantillonner pour les reconstitutions paléoenvironnementales. Ces entités homogènes, que nous avons dénommées « paysages », ont été identifiées à partir de cartes écoforestières (échelle de 1 : 20 000) compilées par le Ministère des Forêts, de la Faune et des Parcs du Québec (MFFP) et qui sont divisées en 49 244 peuplements définis sur la base de diverses variables (importance relative des espèces, la hauteur, la densité, l'âge) et les caractéristiques physiques du site. Afin de bonifier nos connaissances concernant l’âge des peuplements, nous avons élaboré une carte d’origine des peuplements sur laquelle chacun des polygones a été caractérisé selon son année d’origine, ce qui correspond au temps écoulé depuis le dernier feu. Les travaux de terrain (2013 et 2014) ont consisté en l’échantillonnage de 140 sites dans lesquels 313 arbres furent échantillonnés via la récolte d’une rondelle au niveau du collet. En laboratoire, les rondelles ont été poncées et le nombre de cernes dénombrés. Par la suite, les données de végétation et des variables explicatives ont fait l’objet d’une analyse de redondance (RDA) qui permet de positionner sur des axes d’ordination chacun des points associés aux polygones composant les cartes écoforestières. Les polygones ont été regroupés à l’aide d’une analyse de groupement K-means (Legendre et Legendre 2012).
Les tourbières échantillonnées
Deux tourbières ombrotrophes (Aspen et Shadow), distantes de 10,5 km l’une de l’autre (Fig. 2), ont été choisies en raison de leurs caractéristiques similaires : elles sont de petite taille et ceinturées de forêts de début de succession (Populus tremuloides, Pinus banksiana). Les deux tourbières sont colonisées par des éricacées (Rhododendron groenlandicum, Chamaedaphne calyculata, Kalmia
angustifolia) sur un tapis de sphaignes. Des épinettes noires arbustives (1-4 m de hauteur) forment
une strate continue, mais de faible densité (20-40 %). La tourbière Aspen (50º00.46' N; 77º00.14' O) se situe à 283 m d’altitude et est d’une superficie de 30 ha. La tourbière Shadow (49º56.57' N; 77º06.43' O), d’une superficie de 15 ha, se trouve à une altitude de 280 m. À la tourbière Aspen, la carotte sédimentaire a été prélevée à 100 m de part et d’autre de la forêt (Fig. 2). À la tourbière Shadow, l’échantillonnage a été fait à une distance de 25 m de la forêt. Les échantillonnages se sont effectués par découpage de monolithes de tourbe (30 × 30 × 30 cm) au sein de tranchées excavées à
7
la pelle jusqu’au contact de l’argile sous-jacente. Les monolithes ont été enveloppés dans une pellicule plastique et du papier d’aluminium. Ils furent ensuite entreposés au laboratoire au frais à 5°C jusqu’au moment des analyses.
Shadow Aspen
Figure 2 : Photos terrestres (haut) et photographies aériennes (bas; échelle 1: 20 000) des forêts entourant les tourbières
Aspen et Shadow. Le site d’échantillonnage de la carotte sédimentaire est représenté par le carré noir. Crédit photo: Pierre Grondin
La forêt contemporaine ceinturant les deux tourbières à l’étude
Pour caractériser la forêt qui ceinture chacune des tourbières, trois transects parallèles (50 × 4 m ; 200 m2) distants de 10 m l’un de l’autre ont été échantillonnées. Pour chacun, les tiges vivantes de plus de 2 cm de diamètre à hauteur de poitrine ont été identifiées à l’espèce. Celles constituant la régénération (moins de 2 cm de diamètre) ont été identifiées puis dénombrées au sein de placettes de 1 m2 réparties à tous les 5 m au sein des transects (10 placettes par transect). Enfin, cinq tiges par site d’étude ont été tronçonnées afin de récolter la rondelle basale pour connaitre l’année d’origine des peuplements. Les dénombrements de tiges ont été classés par diamètre et transformés en surface terrière totale par espèce. Les tiges de régénération ont été analysées afin de définir une fréquence des espèces par site d’échantillonnage (coefficient de distribution). Enfin, deux échantillons de mousses de surface ont été récoltés et analysés pour leur contenu pollinique actuel. Un échantillon provient de la forêt bordant la tourbière Aspen et, l’autre, d’une pessière noire à sapin à une dizaine de kilomètres plus au nord.
Stratigraphie
Préalablement aux analyses paléoécologiques, les deux séquences sédimentaires ont été découpées en tranches contigües de 1 cm d’épaisseur. La composition générale de la matrice de la tourbe (sphaignes, herbacées, mousses brunes, matière ligneuse) fut déterminée de façon qualitative à partir de l’analyse de sous-échantillons à la loupe binoculaire.
9
La reconstitution de l’historique des feux
Dans le but d’établir la chronologie de l’historique des feux ayant eu lieu au sein des forêts bordant les deux tourbières, des sous-échantillons (2 cm3) ont été prélevés à tous les 2 cm d’intervalle pour une analyse de charbon de bois de taille macroscopique. Ils ont été défloculés dans une solution d’hydroxyde de potassium (KOH 5 %), puis tamisés dans un tamis à mailles 0.450 mm. Les charbons supérieurs à cette de taille proviennent surtout de feux locaux (Hornberg et al. 1995; Ohlson et Tryterud 2000; Lynch et al. 2004). Les particules ont été dénombrées dans une boîte de pétri en utilisant un stéréomicroscope (20×) selon les procédures standards de Hornberg et al. (1995). Chaque particule noire opaque, aux côtés anguleux, fragile et avec une surface argentée brillante et souvent striée a été identifiée comme étant un charbon de bois (Scott 2010). Les charbons ont été séchés, comptés et pesés à l'aide d'une balance électronique. Les données sont exprimées en anthracomasse (mg/2 cm3).
La reconstitution de l’histoire de la végétation
Afin de reconstituer l’histoire postglaciaire de la végétation ceinturant les tourbières Aspen et Shadow, des sous-échantillons (1-3 cm3, en fonction du contenu en pollen) ont été prélevés à tous les 2 cm durant les périodes de forte fréquence de feu (détectées préalablement par l’analyse des charbons de bois) et à tous les 4 cm durant les périodes de faible fréquence de feu. Les premiers centimètres à la base des monolithes correspondant à de l’argile n'ont pas été analysés car ils étaient très pauvres en pollen. Les échantillons ont été traités chimiquement suivant la méthode de Faegri et Iversen (1989). Afin de permettre l'estimation des concentrations polliniques (grains/cm-3), des comprimés de concentration connue de spores de Lycopodium ont été ajoutés à chaque sous- échantillon avant la préparation (Stockmarr 1971). Au moins 300 grains de pollen de plantes vasculaires terrestres (à l'exclusion des Ericaceae et Cyperaceae) ont été comptés (somme pollinique) pour chaque niveau en utilisant un microscope optique (grossissement 400×). Les grains de pollen et les spores ont été identifiés à l’aide d’atlas (Richard 1970 ; McAndrews et al. 1973) et de la collection de référence du Centre d'études nordiques de l’Université Laval. Les résultats sont exprimés en pourcentages. Seules les courbes polliniques des principaux taxons sont présentées sur les diagrammes polliniques. Les diagrammes ont été subdivisés en zones d’assemblage pollinique (ZAP) à l’aide d’un dendrogramme produit par le programme CONISS du logiciel Tilia (Grimm 1987). Pour faciliter la comparaison des deux sites d’étude, les limites des ZAP ont été déplacées afin de tenir compte de la chronologie et des résultats d’une analyse en composantes principales (ACP; voir plus bas). Les ZAP finales illustrées sont communes pour les deux diagrammes. Pour la zonation, les pourcentages polliniques ont été transformés par une racine carrée. Cette transformation permet d’augmenter l’importance des espèces faiblement représentées par leur pollen (p. ex. : Populus) et de réduire l’importance de celles fortement représentées (p. ex. : Betula).
Une analyse en composantes principales (ACP) a été réalisée pour chacun des spectres polliniques afin de synthétiser les grands changements survenus au sein du couvert végétal au cours de l’Holocène et de les comparer avec l’historique des feux. Comme les variables ont une dimension homogène, le calcul des composantes principales a été fait sur une matrice de dispersion (variance/covariance) en utilisant le logiciel R (http://cran.r-project.org/). La matrice soumise à l’ACP contient les pourcentages polliniques transformés par une racine carrée de tous les taxons polliniques recensés, à l’exception des Cyperaceae et des Ericaceae. Cette transformation a été
11
réalisée afin d'optimiser le rapport signal-bruit et de stabiliser la variance. Le nombre de taxons polliniques a été réduit en ne sélectionnant que ceux ayant une valeur supérieure ou égale à 1 % dans au moins un spectre pollinique et en créant un taxon pollinique regroupant l’ensemble des herbacées. La taille de matrice, c’est-à-dire, le nombre de niveaux analysés (spectres polliniques) et nombre de taxons pollinique retenus est de 54×21 pour la tourbière Aspen et de 41×17 pour la tourbière Shadow.
Datations radiochronologiques
Au total, 20 échantillons ont été soumis à la datation au radiocarbone par spectrométrie de masse par accélérateur (SMA). Ils ont été préalablement préparés au laboratoire de radiochronologie du Centre d’études nordiques pour ensuite être datés au Laboratoire Kerk (Université de Californie, Irvine). Les échantillons consistent en des particules de charbon de bois, à l’exception d’un échantillon à la base d’une des carottes (pièces macrofossiles). Les niveaux contenant les anthracomasses les plus élevées ont été sélectionnés pour les datations. Le logiciel CALIB 7.0.4 et la base de données IntCal13 (Reimer et al. 2013) ont été utilisés pour déterminer les distributions de probabilité en années étalonnées associées à chacune des dates 14C (± 2 sigmas). La médiane de la distribution de probabilité a été retenue comme date étalonnée. Les charbons dont les dates se chevauchent dans la distribution de probabilité ont été considérés comme provenant du même feu. Les datations ont été utilisées pour établir un modèle âge-profondeur pour chacun des sites et pour calculer les taux d’accumulation sédimentaire nets (après décomposition) de la tourbe. Les modèles ont été développés en utilisant le logiciel CLAM 2.2 (Blaauw 2010) en appliquant une interpolation linéaire entre chaque niveau daté (1000 itérations). Les résultats sont partout exprimés en années étalonnées A.A.
Résultats
Les paysages forestiers contemporains de la région d’étude
Le territoire d’étude (4000 km2) a été segmenté en huit paysages (Fig. 3). Les paysages similaires entre-eux ont été regroupés en quatre entités (P1 à P4). Chacune des entités montre des caractéristiques particulières en regard de l’altitude, de la proportion des types de dépôts de surface, de la superficie occupée par les peuplements sur sol bien drainé (drainage mésique), de l’année d’origine (relativement au dernier feu) et de la composition des peuplements.
Les paysages P1 (P1a, P1b) sont relativement bas en altitude (Fig. 3a) et fort bien pourvus en dépôts organiques (˃30 % de la superficie totale; Fig. 3b). Les peuplements de drainage mésique sont peu abondants (Fig. 3c). Les pessières noires dominent, tout particulièrement les pessières noires à sphaignes (Fig. 3d). Pour leur part, les espèces de début de succession (Pinus banksiana,
Populus tremuloides) sont très rares (Fig. 3e). Les forêts originent des incendies des périodes 1820
et 1910 (Fig. 3f). Les paysages P2 (P2a, P2b) s’inscrivent le long d’un gradient altitudinal qui va en s’accentuant. La quantité de dépôts organiques est moindre que dans les précédents (15-25 %). Les peuplements de drainage mésique sont faiblement représentés (moins de 30 %). Ceux d’altitude supérieure à 275 m sont rares dans P2a alors qu’ils sont bien représentés dans P2b. Les pessières noires sur sol minéral dominent et proviennent surtout des feux des périodes 1820 (P2a) et 1910 (P2b). Les peuplements de début de succession sont relativement bien représentés dans P2b.
Les paysages P3 (P3a, P3b) sont caractérisés par peu de dépôts organiques (10-15 %). Les peuplements de drainage mésique sont bien représentés (˃30 %). Les peuplements d’altitude supérieure à 275 m augmentent en abondance depuis P3a vers P3b. Les pessières noires sur sol minéral dominent alors que les pessières noires à sapin s’observent ici et là. Les peuplements originant de la période 1820 abondent. Les paysages P4 (P4a, P4b) ne comportent que très peu de dépôts organiques (<10 %). Les peuplements d’altitude supérieure à 275 m abondent. Ceux de drainage mésique dominent et le relief est ondulé; plus de la moitié des pentes associées aux peuplements forestiers sont supérieures à 4 %. Les pessières noires sur sol minéral sont largement distribuées. Le peuplier faux-tremble et le pin gris sont bien représentés : le peuplier faux-tremble est clairsemé dans P4a alors qu’il est abondant dans P4b. Les peuplements issus de la période 1910
13
(Aspen et Shadow) sont situées dans le paysage P4b parce que les peuplements dominés par les espèces de début de succession y sont bien représentés. De plus, ce type de configuration est assez bien répandu hors du présent territoire d’étude, autant dans le reste de la région écologique de la plaine argileuse du lac Matagami (domaine bioclimatique de la pessière noire) que plus au sud dans la région écologique de la plaine de l’Abitibi au sein du domaine bioclimatique de la sapinière (Fig.1).
Figure 3 : Délimitation des paysages forestiers du territoire d’étude (P1-P4). A) Altidude (m). B) Répartition des dépôts
de surface. C) Répartition des peuplements de drainage mésique. D) Répartition des pessières noires sur sol minéral et des pessières noires sur sol organique. E) Répartition des peuplements forestiers dominés par des tremblaies et des pinèdes grises. F) Répartition des peuplements dont la période d’origine est 1820 et 1910. Les deux tourbières à l’étude se situent
15
La forêt contemporaine ceinturant les deux tourbières à l’étude
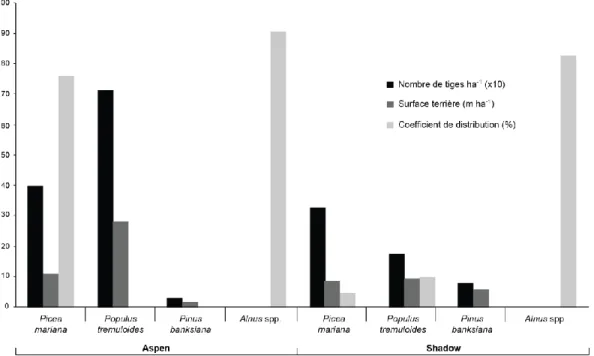
La forêt entourant la tourbière Aspen est de type feuillu (Fig. 4). La surface terrière est dominée par Populus tremuloides (70 %) et sous-dominée par Picea mariana (40 %), tandis que Pinus
banksiana (3 %) est faiblement représenté. Le coefficient de distribution de la régénération (CDR),
mesurant le taux d’occupation d’une superficie, est formée d’épinette noire (76 %) et d’une combinaison (90 %) d’aulne crispé (Alnus viridis subsp. crispa) et d’aulne rugueux (Alnus incana subsp. rugosa). Le feu le plus récent est survenu en 1917. La forêt ceinturant la tourbière Shadow est de type mélangé. La surface terrière est composée de P. mariana (37 %), P. tremuloides (21 %) et P. banksiana (10 %), alors que la régénération est dominée par les aulnes (83 %) avec un peu de
P. tremuloides (10 %). La forêt provient aussi du feu de 1917.
L'assemblage pollinique de l'échantillon de surface provenant de la forêt jouxtant la tourbière Aspen est essentiellement caractérisé par Picea (43 %), Pinus banksiana (36 %), Alnus (10 %) et
Betula (5 %) (Tableau 1). Malgré que le peuplier soit aujourd’hui dominant au sein de la forêt, il
est absent de cet assemblage. L’assemblage de l'échantillon provenant de la pessière à sapin (paysage P3b ; Fig. 3a) est pour sa part principalement composée par Picea (49 %), P. banksiana (23 %), Alnus (12 %), Betula (10 %) et Abies balsamea (2 %).
Figure 3 : Caractéristiques des forêts ceinturant les deux tourbières à l’étude, en regard du nombre de tiges par espèce, de
la surface terrière et de la régénération (coefficient de distribution).
Tableau 1 : Représentation pollinique en pourcentage (espèces sélectionnées) de la végétation actuelle de
deux peuplements distincts dans le territoire d’étude.
Site et espèces dominantes du peuplement Picea mariana Pinus banksiana Abies balsamea Betula Populus tremuloides Alnus Tourbière Aspen P. tremuloides, P. banksiana, P. mariana 43.3 36.2 0.9 5.4 0 10.6 10 km au nord de la tourbière Aspen P. mariana, A. balsamea 49.4 23.1 2.1 9.9 0 12
17
Reconstitutions paléoenvironnementales
Stratigraphie et données chronologiques
Aux points d’échantillonnage des tourbières Aspen et Shadow, le début de l’entourbement a été daté respectivement à 7990 et 4110 ans A.A. (Tableau 2). Aux deux endroits, la matière organique repose sur de l’argile (Fig. 5). L’épaisseur de matière organique à Aspen est de 134 cm. Le taux d’accumulation sédimentaire fut en moyenne de 0,18 mm/an de 7990 à 5200 ans A.A.. Une diminution est ensuite survenue de 5200 à 840 ans A.A. (0,03 à 0,08 mm/an). Elle fut suivie d’une légère augmentation (0,14 mm/an) pendant quelques siècles (900 à 100 ans A.A.). L’épaisseur de matière organique de la tourbière Shadow est de 96 cm (Fig. 5). La présence de nombreux fragments de bois de bonne dimension au contact organo-minéral laisse croire l’entourbement à cet endroit a débuté par un processus de paludification forestière suite au développement spatial de la tourbière. Le taux d’accumulation sédimentaire fut de 0,07 mm/an entre 4110 et 2900 ans A.A. Il a ensuite augmenté de façon importante (0,37 mm/an) de 2900 à 1900 ans A.A. pour ensuite diminuer à nouveau de 1900 à 100 ans A.A. (0,16 mm/an). Pour les deux tourbières, le taux est très élevé dans la portion supérieure en raison d’une moindre compaction et décomposition de la matière organique comparativement aux niveaux inférieurs.
Tableau 2 : Données radiochronologiques des tourbières Aspen et Shadow. Les dates exclues des modèles âge-
profondeur sont en caractère italique.
Site et profondeur
(cm)
Numéro de
laboratoirea Matériel daté Âge 14C
(années A.A.) (années A.A.) Intervalle 2σ (années A.A.) Médiane Aspen 37-38 145391 - 4797 Charbon 180 ± 30 136-224 180 38-39 150957 - 5063 Charbon 75 ±15 33-73 50 44-45 145611 - 4828 Charbon 235 ± 20 281-307 290 49-50 52-53 145608 - 4825 150956 - 5062 Charbon Charbon 955 ± 20 915 ± 15 796-874 840-910 840 880 64-65 150951 - 5057 Charbon 2275 ± 15 2306-2346 2330 77-78 150952 - 5058 Charbon 3590 ± 20 3839-3929 3880 81-82 150955 - 5061 Charbon 4345 ± 20 4856-4963 4910 84-85 145607 - 4824 Charbon 4705 ± 20 5325-5409 5370 132-133 145602 - 4819 Graines 7185 ± 25 7956-8026 7990 Shadow 21-22 145390 - 4796 Charbon 120 ± 30 51-149 100 24-25 145609 - 4826 Charbon 240 ± 20 281-308 300 32-33 145603 - 4820 Charbon 945 ± 20 796-874 835 41-42 145612 - 4829 Charbon 1760 ± 20 1608-1724 1670 44-45 145604 - 4821 Charbon 1680 ± 20 1539-1619 1580 45-46 150954 - 5060 Charbon 1655 ± 15 1524-1573 1550 50-51 145605 - 4822 Charbon 2010 ± 20 1920-1998 1960 85-86 145606 - 4823 Charbon 2790 ± 25 2842-2956 2900 93-94 145610 - 4827 Charbon 3425 ± 20 3613-3720 3680 94-95 145582 - 4783 Charbon 3745 ± 25 4068-4157 4110
19
Reconstitution de l’historique des feux de forêt
Sur la base des valeurs d’anthracomasse et des datations 14C des charbons de bois, huit feux furent identifiées, autant à la tourbière Aspen qu’à la tourbière Shadow (Fig. 6 et 7; Tableau 2). Pour chacune des tourbières, un contenu élevé en charbons de bois était présent sur quelques centimètres d’épaisseur à quelques endroits dans la carotte sédimentaire. À la tourbière Aspen (Fig. 6), les anthracomasses sont très faibles entre la base du dépôt et le premier feu daté à 5370 ans A.A. Quatre des huit incendies ont eu lieu il y a plus de 2000 ans (5370, 4910, 3880 et 2330 ans A.A.), et quatre au cours des 1000 dernières années (880-840, 290, 180 et 50 ans A.A.). Des charbons ont été trouvés à une profondeur d'environ 60 cm, mais ils étaient trop petits et trop peu nombreux pour en faire la datation. À la tourbière Shadow (Fig. 7), trois des huit feux datés ont eu lieu il y a plus de 2000 ans (4110, 3680 et 2900 ans A.A.). Les cinq autres ont été datées à 1960, 1670, 1550-1580, 835, 300 et 100 ans AA. Seul le dernier millénaire est marqué par des feux relativement synchrones entre les deux sites : 880-840 (Aspen) et 835 ans A.A. (Shadow), 290 (Aspen) et 300 ans A.A. (Shadow), 50 (Aspen) et 50 ans A.A. (Shadow). Enfin, les dates obtenues les plus récentes sont en lien avec les données dendrochronologiques : la date de 180 ans A.A. est associée aux feux de la décennie 1820 et celles de 100 et 50 ans A.A. aux feux de la période 1910.
Reconstitution de l’histoire de la végétation
L’histoire postglaciaire de la végétation ceinturant les tourbières Aspen et Shadow est reconstituée sur la base des deux diagrammes polliniques. Au total, quatre zones d’assemblages polliniques (ZAP 1 à 4) ont été délimitées. Puisque l’accumulation de la matière organique à la tourbière Aspen a débuté près de 4000 ans avant celle de la tourbière Shadow, son diagramme pollinique montre la présence de deux zones (ZAP 1 et 2) associées à la phase d’afforestation et au début de la phase forestière qui sont absentes du diagramme pollinique de la tourbière Shadow.
La ZAP 1 (ca. 8000 à ca. 7600 ans A.A.), à la base du diagramme de la tourbière Aspen, montre une représentation pollinique relativement élevée des herbacées (Artemisia, Tubulifloreae, Chenopodiaceae, Poaceae) et de certains arbustes (Salix, Alnus). Les taxons arboréens sont représentés par Picea (fort probablement Picea mariana), Pinus banksiana et Betula (fort probablement Betula papyrifera), bien qu’il soit possible qu’une part du pollen de bouleau soit aussi attribuée au bouleau glanduleux (Betula glandulosa). Les très faibles concentrations
21
polliniques suggèrent que cette première étape de la colonisation végétale suite au retrait des eaux du lac proglaciaire Ojibway était caractérisée par des forêts très ouvertes correspondant à la phase d’afforestation, et qu’une certaine proportion du pollen arboréen était de source lointaine. La ZAP 2 (ca. 7600 à ca. 6800 ans A.A.) montre une augmentation graduelle des concentrations polliniques et une diminution de la représentation des taxons herbacés et arbustifs, ce qui suggère une densification du couvert forestier. Les assemblages polliniques sont essentiellement dominés par Picea (40-50%) et P. banksiana (20-30%). Le bouleau atteint des valeurs maximales (50-60%) au sommet de la zone.
La ZAP 3 (ca. 6800 à ca.780-735 ans A.A.) couvre une longue période relativement homogène au cours de laquelle Abies balsamea est bien représenté (maximum : 10 %), tout comme Picea (45- 65 %). Trois sous-zones (3a, 3b et 3c) ont été distinguées. La sous-zone 3a couvre une période de plus de 3000 ans à la tourbière Aspen (ca. 6800 à ca. 3100 ans A.A. à Aspen ; ca. 6800 ca. 2600 ans A.A. à Shadow). Elle montre des pourcentages relativement élevés de Pinus banksiana, Betula et Alnus. Dans la sous-zone 3b (ca. 3100 à ca. 2200 ans A.A. à Aspen ; ca. 2600 à ca. 2000 ans A.A. à Shadow), une baisse de la représentation pollinique de P. banksiana et d’Alnus est notée. Enfin, la sous-zone 3c (ca. 2200-2000 à ca. 780-735 ans A.A.) se caractérise par une légère augmentation de P. banksiana dans le diagramme de la tourbière Aspen et de P. banksiana et
Alnus dans celui de Shadow.
Pour les deux sites, la ZAP 4 correspond aux 780-735 dernières années (Fig. 6 et 7). Elle est subdivisée en deux sous-zones : 4a (ca. 780-735 à ca. 160-115 ans A.A.) et 4b (ca. ˂160- 115 ans A.A.). Les faibles concentrations polliniques résultent du caractère peu décomposé et moins compacte de la matière organique qui a pour effet une dilution du pollen dans la tourbe. La représentation du pollen d’Abies balsamea diminue de la base au sommet de la sous-zone 4a puis disparaît pratiquement dans la sous-zone 4b, en particulier dans le diagramme de la tourbière Aspen. Dans les deux diagrammes, le début de sous-zone 4b est marqué par une augmentation notable de Pinus banksiana (> 40% à Aspen; > 30% à Shadow). L'espèce y atteint sa représentation pollinique maximale dans cette sous-zone.
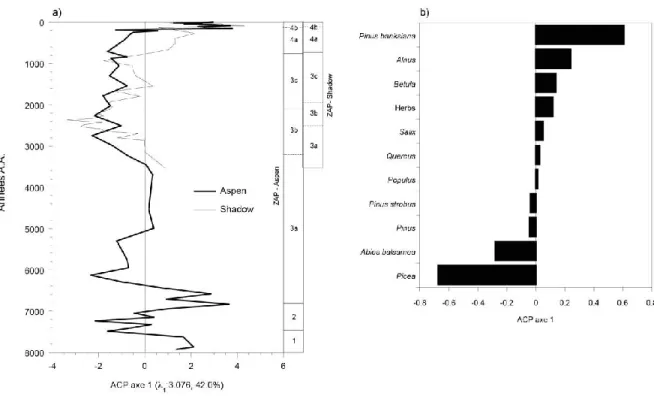
L’histoire postglaciaire de la végétation ceinturant les deux tourbières a été synthétisée à l’aide d’une ACP. La tendance principale des ZAP des deux sites a été comparée objectivement en appliquant l’ACP à l’ensemble combiné des données polliniques. Le premier axe de l’ACP
représente 42,0% (λ1 = 3,076) de la variance des données et montre le gradient majeur de végétation (Fig. 8a). Les fortes notes (valeurs positives) de l’axe 1 sont causées principalement par
Pinus banksiana, Alnus et Betula, trois espèces de début de succession. Les faibles notes (valeurs
négatives) sont quant à elles, causées par Picea et Abies balsamea, deux espèces associées aux paysages de fin de succession (Fig. 8b). La tendance des notes dans l'axe 1 de l’analyse est similaire pour les deux sites. Les quatre ZAP sont également mis en évidence par l'axe 1. À Shadow et Aspen, il y a un changement prononcé vers 100-150 ans A.A. (ZAP 4b / 4a). Le changement est toutefois plus graduel à Shadow comparativement à Aspen où il est plus abrupt.
23
Figure 5 : Diagramme pollinique (pourcentages; taxons polliniques choisis) et de l’anthracomasse de la tourbière Aspen.
Figure 6 : Diagramme pollinique (pourcentages; taxons polliniques choisis) et de l’anthracomasse de la tourbière
25
Figure 7 : Résultat de l’analyse en composantes principales (ACP) des assemblages polliniques combinés des sites
Aspen et Shadow. a) Ordination stratigraphique de la note des échantillons sur le premier axe. Les résultats sont présentés selon une échelle de temps. La première valeur propre (λ1) et le pourcentage de la variance sont donnés. b)
Discussion et conclusion
Cette étude présente une reconstitution de l’histoire de la végétation et des feux d'une partie du domaine bioclimatique de la pessière à mousses de l’Ouest. Nous avons analysé les sédiments de deux tourbières qui se trouvent à 10,5 km de distance l’une de l’autre, dans un paysage où les espèces de début succession sont bien représentées. Ces paysages proviennent pour la plupart de feux de la décennie 1910 (plus précisément de l’année 1916). La carotte de sédimentaire de la tourbière Aspen, recueillie à une distance de 80-100 m de la bordure de la forêt, offre une histoire continue de près de 8000 ans, c’est-à-dire, depuis le retrait des eaux du lac proglaciaire Ojibway. Pour sa part, l’accumulation sédimentaire de la tourbière Shadow, échantillonnée au contact de la forêt, couvre une période d'un peu plus de 4000 ans.
La dynamique plurimillénaire des paysages
L'impact du feu sur la végétation varie de court terme (< 200 ans) à plus long terme. Après un incendie, les espèces au début de succession (Pinus banksiana, Populus tremuloides, Betula
papyrifera, Alnus spp.) sont les premières à s’installer. Avec le temps, elles sont progressivement
remplacées par des espèces de fin de succession, notamment Picea mariana et Abies balsamea (Gauthier et al. 2008), à moins qu’un feu surviennent à nouveau dans un court laps de temps. On peut supposer qu'une modification de la fréquence des incendies peut mener à des changements au sein de la composition des forêts et de la structure des peuplements (Bergeron et al. 2004 ; Shorohova et al. 2009 ; Kuosmanen et al. 2014). En outre, l'activité des feux varie considérablement en fonction des conditions physiques stationnelles comme l'altitude, les obstacles à la propagation du feu et le type de dépôt de surface.
À la tourbière Aspen, seulement quelques petits charbons de bois isolés (profondeurs 116 cm et 109 cm ~ ca. 7000 ans A.A.) ont été trouvés dans les sédiments qui représentent la période comprise entre le drainage des eaux du lac proglaciaire Ojibway (environ 8000 ans A.A.) et le premier feu daté à 5370 ans A.A. (Fig. 6). Cette période correspond aux ZAP 1 et 2 (phase d’afforestation), ainsi qu’au début de la ZAP 3 (sous-zone 3a ; début de la phase forestière). Il est possible que la rareté des charbons de bois à cette époque soit liée à la faible densité du couvert forestier au cours de la période d’afforestation qui ne permettait pas encore de soutenir une activité récurrente des feux. Cependant, l'analyse des sédiments de quelques lacs situés à l'ouest et au nord
27
de la présente zone d'étude dans le domaine bioclimatique de la pessière à mousses a montré que des incendies avaient eu lieu au cours de cette période (Carcaillet et al. 2001 ; Oris et al. 2014). La rareté des charbons de bois et des macrorestes carbonisés pourrait aussi être de nature taphonomique : les conditions locales très humides présentes initialement à la tourbière Aspen auraient pu empêcher le feu de se propager vers le centre de la tourbière. Cette possibilité est soutenue par la présence de mousses brunes et de graines de Menyanthes trifoliata qui indique qu’à l’époque, la tourbière Aspen était minérotrophe et saturée en eau.
Le début de la phase forestière à la tourbière Aspen, vers 6800 ans A.A. (sous-zone 3a), est marqué par une augmentation de la représentation pollinique d’Abies balsamea (Fig. 6), une espèce de fin de succession qui est généralement sous-représentée dans les assemblages polliniques (Richard 1993). Le sapin a été relativement abondant jusqu’à tout récemment, soit au milieu de la sous-zone 4a. Cette plus grande abondance du sapin semble avoir été favorisée par une récurrence des feux relativement faible entre ca. 6800 et ca. 1000 ans A.A., période pendant laquelle seul quatre épisodes ont été recensés (5370, 4910, 3880 et 2330 ans A.A.). La présence de fragments de charbon bois en petites quantités à d'autres niveaux de la carotte sédimentaire suggère toutefois qu’il s’agit d’un nombre minimal, et qu’il est possible que d’autres feux aient eu lieu.
À la tourbière Shadow, les feux documentés durant la ZAP 3 (4110, 3680, 2900, 1960, 1670 et 1550-1580 ans A.A.) se sont produits à des dates différentes des feux survenus à la tourbière Aspen. Comme les deux sites ne sont distants que d’une dizaine de kilomètres, cet asynchronisme suggère que les incendies étaient de faible intensité et/ou ne couvraient que de petites superficies. Un tel régime de feu aurait permis aux populations de sapin de se maintenir à long terme et que les paysages forestiers devaient correspondre à des pessières noires à sapin. On note toutefois que les espèces pionnières (Pinus banksiana, Betula papyrifera, Alnus spp.) étaient aussi bien représentées dans les diagrammes polliniques, en particulier dans celui de la tourbière Shadow.
Les anthracomasses enregistrées pour la période correspondant à la sous-zone 3b (ca. 2500 à ca. 2000 ans A.A.) sont faibles, en particulier pour la tourbière Shadow où aucun charbon n'a été trouvé. La représentation pollinique d’Abies balsamea est relativement élevée au cours de ces quelques siècles, alors que celle de Pinus banksiana, Betula et Alnus spp. diminue. Comme précédemment, les paysages forestiers étaient caractérisés par des pessières noires à sapin. De
faibles fréquences de feu entre ca. 3000 et ca. 2000 ans A.A. ont également été identifiées ailleurs dans l'ouest du Québec (Ali et al. 2009 ; 2012 ; Oris et al. 2014 ; El-Guellab et al. 2015). Les valeurs des anthracomasses de la sous-zone 3c à Aspen (ca.2100 à ca. 800 ans A.A.) sont faibles et comparables à celles de la sous-zone 3b. La représentation pollinique de A. balsamea reste constante, tandis qu'une augmentation de P. banksiana, Betula et Alnus est observée, en particulier à la tourbière Shadow. Cette augmentation des espèces pionnières est bien illustrée par les résultats de l'ACP pour les deux sites (Fig. 8). Les résultats des analyses anthracologiques et polliniques suggèrent donc que les feux ont permis, durant la majeure partie de l’Holocène, le maintien d’une mosaïque forestière dans laquelle l’association sapin baumier-épinette était dominante.
Au cours des 1000 dernières années (ZAP 4), trois des quatre incendies recensés semblent avoir affecté les deux tourbières simultanément (835-880/840, 290-300, 50-100 ans A.A.). Un feu daté à 180 ans A.A. n’a pour sa part, été enregistré qu’à la tourbière Aspen. Les assemblages polliniques de la ZAP 4 sont principalement dominés par Pinus banksiana. C’est d’ailleurs au cours de lapériode représentée par la ZAP 4 que les pourcentages de cette espèce sont les plus élevés pour les 8000 dernières années. Bien que faible, une légère augmentation de la représentation des
Populus est aussi notée dans le diagramme de la tourbière Shadow (sous-zone 4b). Il est bien
établi que le peuplier est fortement sous-représenté dans les assemblages polliniques (Mott 1978 ; Comtois et Payette 1984).
Le fait que les forêts qui bordent les deux tourbières furent affectées par les mêmes feux à trois reprises suggère fortement que les incendies ont été plus sévères et ont couvert de plus grandes superficies que ceuxayant eu lieu auparavant. Dans cette région, deux incendies, à la fois sévères et étendus, ont eu lieu au 19e et 20e siècles, plus précisément en 1820 et 1917 (Bergeron et al. 2001 ; 2004 ; Cyr et al. 2009). Le feu daté à 180 ans A.A. à la tourbière Aspen correspondrait à celui de 1820, et le feu daté à 50-100 ans A.A. aux deux sites à celui de 1917. Il est ainsi possible d’émettre l’hypothèse que le régime de feu ayant caractérisé le dernier millénaire (feux sévères couvrant de vastes étendues) aurait favorisé le maintien des espèces pionnières jusqu'à nos joursselon une dynamique cyclique. Ces mêmes feux ont conduit à la fragmentation graduelle de la répartition de A. balsamea. Aujourd’hui, cette espèce est principalement cantonnée aux rives descours d’eau et aux versants de collines qui sont moins susceptibles d’être affectés par le feu. Ce ne serait toutefois qu’au cours des 200-300 dernières années que le sapin se serait vraiment raréfié. Cette interprétation est en désaccord avec celle de Ali et al. (2008) qui, pour leur part, ont proposé
29
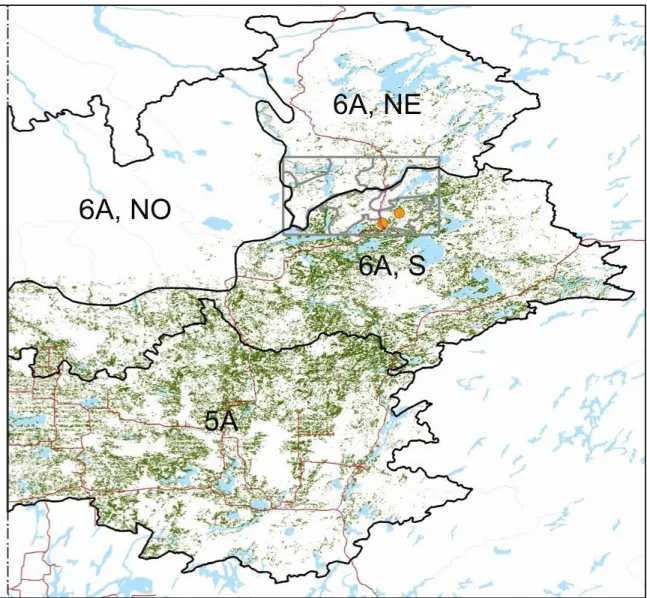
une fragmentation dès 3500 ans avant nos jours au sud-ouest du domaine de la pessière à mousses de l’Ouest (lac aux Cèdres). L'augmentation simultanée de la représentation polliniques d’espèces pionnières (notamment P. banksiana) au cours du dernier millénaire est un trait commun à plusieurs sites de la forêt boréale de l'Ouest du Québec et de l'est de l'Ontario, bien qu’elle ne soit pas nécessairement synchrone d’un endroit à l’autre (Liu 1990 ; Garralla et Gajewski 1992 ; Carcaillet et al. 2001 ; Genries et al. 2012 ; Fréchette et al. 2016). En outre, la dynamique forestière observée au cours du dernier millénaire dans la zone d'étude (paysage P4b) pourrait potentiellements’appliquer à l’ensembles des régions écologiques 5A et 6A (NE, S) caractérisées par la présence de vastes peuplements dominés par Populus tremuloides sur argile mésique- subhydrique (Figure 9). Les caractéristiques y sont similaires en termes d'altitude, de proportion des types de dépôt de surface, de composition des forêts et d'année d'origine des peuplements.
Figure 9 : Territoire auquel les résultats de la présente étude pourraientêtre potentiellement extrapolés. Ce territoire
correspond à la portion sud de la région écologique de la plaine argileuse de l'Abitibi (domaine bioclimatique de la pessière à mousses ; région 6a) et la plaine de l'Abitibi (domaine de la sapinière à bouleau blanc ; région écologique 5a). Les peuplements forestiers dominés par Populus tremuloides sur argile mésique-subhydrique sont illustrés en vert. La zone d'étude où furent menées les analyses paléoécologiques est illustrée par le rectangle gris et les deux tourbières étudiées par les deux points orange.
31
La dynamique contemporaine des paysages
Les forêts contemporaines entourant les deux tourbières sont composées de peuplements de feuillus dominés par Populus tremuloides (tourbière Aspen) et de peuplements mixtes formés par
Picea mariana, Pinus banksiana et P. tremuloides (tourbière Shadow). Cela suggère que la
composition des peuplements forestiers résultant des incendies de la décennie 1910 a été variable sur des dépôts de surface constituée d’argiles mésiques à subhydriques. La régénération actuelle est constituée essentiellement de P. mariana accompagné de quelques P. tremuloides.. Sur la base de cette régénération, les forêts actuelles devraient, en principe, évoluer vers des peuplements de
P. mariana-P. tremuloides (Blouin et al. 2005). Abies balsamea est présentement rare au sein de la
régénération. Si le temps écoulé avant le prochain feu est suffisamment long (de l'ordre de 200-250 ans), il se pourrait que cette espèce atteigne la maturité et se régénère en abondance. Les forêts de fin de succession, telles que celles qui ont caractérisé le paysage durant la majeure partie de l'Holocène jusqu’à tout récemment, pourraient ainsi se former à nouveau. Cette possibilité reste cependant peu probable dans le contexte des changements climatiques en cours, car une augmentation de la fréquence et de la sévéritédes feux est prévue pour les prochaines décennies dans la partie ouest du domaine de la pessière à mousses du Québec (Bergeron et al. 2010 ; Girardin et al. 2013).
RÉFÉRENCES
Ali A.A., C. Carcaillet et Y. Bergeron. 2009. Long-term fire frequency variability in the eastern Canadian boreal forest: the influences of climate vs. local factors. Global Change Biology 15:1230–1241. Ali A.A., C. Carcaillet et Y. Bergeron. 2009. Long-term fire frequency variability in the eastern Canadian boreal forest: the influences of climate vs. local factors. Global Change Biology 15:1230-1241.
Ali, A.A., O. Blarquez, M.P. Girardin, C. Hely, F. Tinquaut, A. El Guellab, C. Carcaillet, V. Valsecchi, A. Terrier, L. Bremond, A. Genries, S. Gauthier et Y. Bergeron. 2012. Control of the multimillennial wildfire size in boreal North America by spring climatic conditions. Proceedings of the National Academy of Science, 109: 20966-20970.
Asselin, M. 2013. Feux contemporains et paysages forestiers d’une portion de la région écologique de la plaine du lac Matagami (6a) dans le sous-domaine bioclimatique de la pessière à mousses de l’Ouest, Québec. Mémoire de fin d’études, Université Laval, xx p.
Belleau, A., A. Leduc, N. Lecomte et Y. Bergeron. 2011. Forest succession rate and pathways on different surface deposit types in the boreal forest of northwestern Quebec. Écoscience 18: 329-340.
Bergeron Y., S. Gauthier, V. Kafka, P. Lefort et D. Lesieur. 2001. Natural fire frequency for the Eastern Canadian boreal forest: consequences for sustainable forestry. Canadian Journal of Forest Research 31: 384-391.
Blaauw, M. 2010. Methods and code for ‘classical’age-modelling of radiocarbon sequences. Quaternary Geochronology, 5: 512-518.
Boucher, Y., M. Bouchard, P. Grondin et P. tardif. 2011. Le registre des états de référence : intégration des connaissances sur la structure, la composition et la dynamique des paysages forestiers naturels du Québec méridional Mémoire de recherche forestière n° 161
Borcard D, F. Gillet F et P. Legendre. 2011. Numerical Ecology with R. Springer, New York. Carcaillet, C., Y. Bergeron, P.J.H. Richard, B. Fréchette, S. Gauthier et Y.T. Prairie. 2001.
Change of fire frequency in the eastern Canadian boreal forests during the Holocene: does vegetation composition or climate trigger the fire regime? Journal of Ecology, 89: 930-946.
Carcaillet, C., P.J.H. Richard, Y. Bergeron, B. Fréchette et A.A. Ali. 2010. Resilience of the boreal forest in response to holocene fire-frequency changes assessed by pollen diversity and population dynamics. International Journal of Wildland Fire, 19: 1026-1039.
33
Comtois, P. et S. Payette. 1984. Représentation pollinique actuelle et subactuelle des peupleraies boréales au Nouveau-Québec. Géographie physique et Quaternaire, 38 : 123-133. El-Guellab, A., H. Asselin, S. Gauthier, Y. Bergeron et A.A. Ali. 2015. Holocene variations of
wildfire occurrence as a guide for sustainable management of the northeastern Canadian boreal forest. Forest Ecosystems, 2: 1-7.
Environment Canada. 2013. National climate data and information archive. Canadian Climate Normals 1971-2000. www.climate.weatheroffice.ec.gc.ca
Faegri, K., P.E. Kaland et K. Krzywinski. 1989. Textbook of Pollen Analysis. 4th ed. John Wiley & Sons, New York.
Fréchette, B., P.J.H. Richard, P. Grondin, M.Lavoie, A.C. Larouche. 2016. Histoire postglaciaire de la végétation et du climat des pessières et des sapinières de l’ouest du Québec. Mémoire de recherche, Direction des inventaires forestiers, ministère des Forêts, de la Faune et des Parcs du Québec.
Frelich, L.E. 2002. Forest Dynamics and Disturbance Regimes: Studies from Temperate Evergreen Forests. Cambridge University Press, Cambridge.
Gauthier, S., A. Leduc, B. Harvey, Y. Bergeron et P. Drapeau. 2001. Les perturbations naturelles et la diversité écosystémique. Naturaliste canadien 125 : 10-17.
Gauthier, S., M-A. Vaillancourt, A. Leduc, L. de Grandpré, D. Kneeshaw, H. Morin, P. Drapeau et Y. Bergeron. 2008. Aménagement écosystémique en forêt boréale. Presses de l'Université du Québec, Montréal.
Garralla, S. et K. Gajewski. 1992. Holocene vegetation history of the boreal forest near Chibougamau, central Québec. Canadian Journal of Botany, 70: 1364-1368.
Grondin, P., J. Noël et D. Hotte. 2003. Envahissement des parterres de coupe par le sapin en forêt boréale. Dans P. Grondin et A. Cimonv(coordonnateurs). Les enjeux de biodiversité relatifs à la composition forestière. Ministère des Ressources naturelles, de la Faune et des Parcs du Québec, Direction de la recherche forestière et Direction de l’environnement forestier, Québec, pp. 15-44.
Grondin, P., S. Gauthier, D. Borcard, Y. Bergeron et J. Noël. 2014. A new approach to ecological land classification for the Canadian boreal forest that integrates disturbances. Landscape Ecology, 29: 1-16.
Girardin, M.P. et M. Mudelsee. 2008. Past and future changes in Canadian boreal wildfire activity. Ecological Applications, 18: 391-406.
Grimm, E.C., 1987. CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Computers and Geosciences, 13: 13-35.
Heinselman M.L. 1973. Fire in the virgin forest of the boundary waters canoe area, Minnesota. Quaternary Research, 3: 329-382.
Hély, C., M.P. Girardin, A.A. Ali, C. Carcaillet, S. Brewer et Y. Bergeron. 2010. Eastern boreal North American wildfire risk of the past 7000 years: A model-data comparison, Geophysical Research Letters, 37: L14709.
Hörnberg, G., M. Ohlson M. et O. Zackrisson. 1995. Stand dynamics, regeneration patterns and long- term continuity in boreal old-growth Picea abies swamp-forests. Journal of Vegetation Science, 6: 291-298.
Kuosmanen, N., K. Fang, R.H.W. Bradshaw, J.L. Clear et H. Seppa. 2014. Role of forest fires in Holocene stand-scale dynamics in the unmanaged taiga forest of northwestern Russia. The Holocene, 24: 503-514.
Kuuluvainen, T. 2002. Natural variability of forests as a reference for restoring and managing biological diversity in boreal Fennoscandia. Silva Fennica, 36: 97-125.
Lecomte, N. et Y. Bergeron. 2005. Successionnel pathways on different surficial deposits in the coniferous boreal forest of the Quebec Clay Belt. Canadian Journal of Forest Research, 35: 1984-1995.
Legendre, P, et L. Legendre. 2012. Numerical Ecology. 3e édition. Elsevier, The Netherlands. Liu, K.-B.1990. Holocene paleoecology of the boreal forest and Great Lakes - St. Lawrence
forest in northern Ontario. Ecological Monographs, 60: 179-212.
Logan T., I. Charron, D. Chaumont et D. Houle. 2011. Atlas de scénarios climatiques pour la
forêt québécoise. Consortium Ouranos. Montréal,
Québec.http://www.ouranos.ca/media/publication/162_AtlasForet2011-Sans-Annexes.pdf [consulté le 7 mars 2012].
35
Lynch, J. A., J.S. Clark et B.J. Stocks. 2004. Charcoal production, dispersal, and deposition from the Fort Providence experimental fire: interpreting fire regimes from charcoal records in boreal forests. Canadian Journal of Forest Research, 34: 1642-1656.
Mansuy N., S. Gauthier, A. Robitaille et Y. Bergeron. 2010. The effects of surficial deposit– drainage combinations on spatial variations of fire cycles in the boreal forest of eastern Canada. International Journal of Wildland Fire, 19: 1083-1098
McAndrews, J.H., A.A. Berti et G. Norris. 1973. Key to Quaternary Pollen and Spores of the Great Lakes Region. Life Science Miscellaneous Publication. Royal Ontario Museum.Toronto, Ontario.
Mott, R.J. 1978. Populus in late-Pleistocene pollen spectra. Canadian Journal of Botany, 56: 1021- 1031.
Ohlson, M. et E. Tryterud. 2000. Interpretation of the charcoal record in forest soils: forest fires and their production and deposition of macroscopic charcoal. The Holocene, 10: 519-525.
Oris, F., H. Asselin, A.A. Ali, W. Finsinger et Y. Bergeron.2013. Effect of increased fire activity on global warming in the boreal forest. Environmental Reviews, 22: 206-219.
Oris, F., H. Asselin, W. Finsinger, C. Hély, O. Blarquez, M-E. Ferland, Y. Bergeron et A.A. Ali. 2014. Long‐term fire history in northern Québec: implications for the northern limit of commercial forests. Journal of Applied Ecology, 51: 675‐683.
Payette, S. 1992. Fire as a controlling process in the North American boreal forest. In H. Shugart, R. Leemans et G.B. Bonan (éditeurs). A System Analysis of the Boreal Forest. Cambridge University Press, Cambridge, pp. 144-169.
Pickett, S.T.A. et P.S. White. 1985. The Ecology of Natural Disturbances and Patch Dynamics. Academic Press, New York.
Reimer, P.J., Bard, E., Bayliss, A., Beck, J. W., Blackwell, P. G., Ramsey, Buck, C. E., Cheng, H., Edwards, R. L., Friedrich, M., Grootes, P. M., Guilderson, T. P., Haflidason, H., Hajdas, I., Hatté, C., Heaton, T. J., Hoffmann, D. L., Hogg, A. G., Hughen, K., A., Kaiser, K. F.,
Kromer, B., Manning, S. W., Niu, M., Reimer, R. W., Richards, D., A., Scott, E. M., Southon, J. R., Staff, R., A., Turney, C. S. M., van der Plicht, J. (2013). INTCAL 13 and MARINE13 radiocarbon age calibration curves 0-50,000 years cal BP. Radiocarbon, 55 (4), 2013, 1869-1887
Richard, P.J.H., 1970. Atlas pollinique des arbres et de quelques arbustes indigènes du Québec. Le Naturaliste canadien, 97 : 1-34 ; 97-161 ; 241-306.
Richard, P.J.H., 1979. Contribution à l’histoire postglaciaire de la végétation au nord-est de la Jamésie, Nouveau-Québec. Géographie physique et Quaternaire, 33 : 93-112.
Richard, P.J.H., 1995. Le couvert végétal du Québec-Labrador il y a 6000 ans BP: un essai. Géographie physique et Quaternaire, 49 : 117-140.