HAL Id: hal-03127291
https://hal.sorbonne-universite.fr/hal-03127291
Submitted on 1 Feb 2021
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Exploring the potential impact of nutritionally
actionable genetic polymorphisms on idiopathic male
infertility: a review of current evidence
Sinda Mahbouli, Charlotte Dupont, Yaelle Elfassy, Eric Lameignère, Rachel
Lévy
To cite this version:
Sinda Mahbouli, Charlotte Dupont, Yaelle Elfassy, Eric Lameignère, Rachel Lévy.
Exploring
the potential impact of nutritionally actionable genetic polymorphisms on idiopathic male
infertil-ity: a review of current evidence. Asian Journal of Andrology, Medknow Publications, In press,
�10.4103/aja.aja_87_20�. �hal-03127291�
considered, from hormonal balance, nutritional requirements, and optimal gut microbiome to exposure to environmental toxins.
Role of obesity and fatty acids as modulators of sperm function
The prevalence of male obesity in reproductive age has nearly tripled in the last 30 years.7 There is increasing awareness that male obesity reduces sperm quality, in particular by altering the physical and molecular structure of germ cells in the testes and mature sperm, and raises the risk of sperm DNA damage linked to excess production of reactive oxygen species (ROS).8,9 Recently, growing evidence has linked abdominal obesity, together with insulin resistance and dyslipidemia, to male fertility.10 Obesity adversely affects male fertility through changes at the hormonal level, as well as by direct changes to sperm function and gamete molecular composition. Adipose tissue depots containing adipocytes and infiltrated immune cells generate inflammatory molecules that may also play an important role in the chronic pro-inflammatory state in the testicular microenvironment and/or excurrent ductal system. Increased adipose tissue is associated with overproduction of adipocytokines, such as leptin, resistin, adiponectin, ghrelin, tumor necrosis factor-α (TNF-α), and interleukin-6 (IL-6), which have a negative influence on spermatogenesis.11
INTRODUCTION
In recent years, several studies have provided evidence that semen quality in humans is decreasing, which may lead to a significant decline in male fertility.1 Moreover, some studies have investigated the possible association between the infertile male phenotype and specific gene variants.2,3 Recent research on impaired sperm quality has demonstrated genetic variation in sperm DNA.4 In fact, the growing technology of genetic tools that use genomic information has also provided an avenue for experimental approaches to support the genetic causes of male infertility. Personal genetic testing can provide information that may be used to recommend dietary choices that are more effective at the individual level than the current dietary advice. A recent systematic review found that individuals are more likely to change health behaviors, including their dietary choices, when their genetic information include actionable advice.5 A large number of micronutrients are required as cofactors for enzymes, or as part of the structure of proteins involved in DNA synthesis and repair, prevention of oxidative damage to DNA, and maintenance methylation of DNA.6 Therefore, genetic variants with poor nutritional and environmental factors may have an impact on fertility, fetal growth, and birth outcomes. To preserve male fertility, the whole person should be
REVIEW
Exploring the potential impact of nutritionally
actionable genetic polymorphisms on idiopathic
male infertility: a review of current evidence
Sinda Mahbouli
1, Charlotte Dupont
2,3, Yaelle Elfassy
3, Eric Lameignère
1, Rachel Levy
2,3Infertility affects about 15% of the world’s population. In 40%–50% of infertile couples, a male factor underlies the problem, but in about 50% of these cases, the etiology of male infertility remains unexplained. Some clinical data show that lifestyle interventions may contribute to male reproductive health. Cessation of unhealthy habits is suggested for preserving male fertility; there is growing evidence that most preexisting comorbidities, such as obesity and metabolic syndrome, are highly likely to have an impact on male fertility. The analysis of genetic polymorphisms implicated in metabolic activity represents one of the most exciting areas in the study of genetic causes of male infertility. Although these polymorphisms are not directly connected with male infertility, they may have a role in specific conditions associated with it, that is, metabolic disorders and oxidative stress pathway genes that are potentially associated with an increased risk of male infertility due to DNA and cell membrane damage. Some studies have examined the impact of individual genetic differences and gene-diet interactions on male infertility, but their results have not been synthesized. We review the current research to identify genetic variants that could be tested to improve the chances of conceiving spontaneously through personalized diet and/or oral vitamin and mineral supplementation, by examining the science of genetic modifiers of dietary factors that affect nutritional status and male fertility.
Asian Journal of Andrology (2021) 23, 1–9; doi: 10.4103/aja.aja_87_20; published online: 26 January 2021
Keywords: energy metabolism; genetic polymorphisms; idiopathic male infertility; lifestyle factors; nutrition; oxidative stress;
semen quality
1FabLife, 104 Avenue Albert 1er, Rueil-Malmaison 92500, France; 2Sorbonne Université, Saint Antoine Research Centre, INSERM Genetic and Acquired Lipodystrophies
Team, Reproductive Biology and CECOS, AP-HP, Tenon Hospital, Paris F-75020, France; 3Reproductive Biology and CECOS, AP-HP, Tenon Hospital, Paris F-75020,
France.
Correspondence: Dr. R Levy (rachel.levy@aphp.fr) Received: 10 May 2020; Accepted: 09 November 2020
Open Access
Male
Inf
er
til
ity
Asian Journal of Andrology
Impact of nutritional SNPs on male infertility
S Mahbouli et al 2
Although some data are currently available on dietary modulation of lipid metabolism, little is known about the nutritional regulation of energy metabolism in sperm. In this regard, lipid profile alterations have been correlated with male infertility.12 There are three types of natural fatty acids, namely, saturated, monounsaturated, and polyunsaturated. Polyunsaturated fatty acids (PUFAs) are essential because they cannot be synthesized by the body. Docosahexaenoic acid (DHA), eicosapentaenoic acid (EPA), and α-linolenic acid are the main omega-3 PUFAs. Linoleic acid, γ-linolenic acid, and arachidonic acid are the main omega-6 PUFAs. The first mechanism by which PUFAs affect spermatogenesis is their incorporation into the spermatozoon cell membrane. PUFAs are structural components of spermatozoon membranes.13,14 Conquer et al.15 reported that DHA levels were lower and oleic acid levels were higher in spermatozoa of patients suffering from asthenozoospermia, compared to that of a control group. Another case-control study of idiopathic infertile men and a healthy control group showed that blood and spermatozoa levels of omega-3 were significantly higher in fertile men compared to that of their infertile counterparts. Moreover, the serum omega-6 to omega-3 proportion was considerably lower in fertile individuals.16 Some studies have reported a detrimental impact of body mass index (BMI) on sperm parameters, notably a decrease in sperm concentration.17,18 Consequently, the possibility of improving semen quality through weight reduction has also been considered. Improvement of hormonal status was mainly observed after weight loss;19 furthermore, a positive impact of weight loss on semen parameters has been observed.20
Association between nutritional factors and the risk of male infertility
Nutritional factors are known to be critical determinants of normal reproductive function.21 Indeed, worldwide, environmental conditions have changed dramatically, especially with respect to diet and exercise, and the pronounced changes in these factors suggest that they may be involved in the etiology of declining male fertility and impairment of sperm production.22 In fact, diet and obesity are two important lifestyle factors that can influence spermatogenesis; in terms of both macro- and micro-nutrient intake, they have major effects on normal reproductive function.23 Within the last few decades, reproductive-age people have started eating more highly refined carbohydrate-rich food, food high in saturated fat and transfatty acids, and sodium and ultra-processed food, while simultaneously consuming less fresh fruit and vegetables.16 There is increasing evidence indicating a potential relationship between incorrect nutritional attitudes and lower sperm quality.12 Essential nutrients, especially vitamins such as folate, are involved in DNA and RNA synthesis, and thus play an important role in spermatogenesis by protecting the sperm’s DNA from free radical damage.24 A recent meta-analysis of randomized clinical trials suggests that some dietary supplements could beneficially modulate sperm quality parameters and affect male fertility.25 However, no consensus has been reached on systematic recommendation of oral supplementation.
Impacts of oxidative stress on male reproduction
Evidence has been increasing in recent years for oxidative stress playing a vital role in the pathogenesis of idiopathic male factor infertility. One of the factors that may contribute to the onset of male infertility is the overproduction of ROS.26 Aitken27 (2016) reviewed twenty studies of infertile male patients treated with antioxidants. The review showed a significant decrease in oxidative stress and improved motility in asthenozoospermic patients, but only 50% of the studies reported a pregnancy rate.28 However, there is weak evidence
from a few randomized controlled trials suggesting that antioxidant supplementation in subfertile males may improve live birth rates in couples attending fertility clinics.25 Further studies are required to formulate an optimal dosage and ideal combination of nutrients, both necessary to provide the appropriate response to each patient.
GENETIC VARIANTS ASSOCIATED WITH NUTRITION AND MALE REPRODUCTIVE POTENTIAL
Single-nucleotide polymorphisms (SNPs) represent genetic variation among individuals in a population. These variations in the DNA sequence may significantly affect an individual’s response to certain drugs or influence the risk of developing certain diseases. In the field of reproductive medicine, considerable research effort has been devoted to identifying polymorphisms which may influence steroidogenesis and fertility. Genetic risk involved in spermatogenesis is considered one of the main factors in male infertility. In recent years, various studies have reported possible associations between infertile phenotypes and specific genetic variants. The group of variants related to energy metabolism, folate metabolism, and antioxidant defense probably influences male fertility, with the impact of the variants potentially modulated by nutritional interventions. Genetic variation affecting responses to various micro- and macronutrients, as well as bioactives such as folate, and male infertility risk will be reviewed in this paper. Genes were selected based on their potential contribution to infertile male phenotypes and a presumed involvement in various parts of the pathogenic processes of male infertility.
Genetic variants involved in energy balance and lipid metabolism, and their influence on male fertility
The physiological mechanisms that control energy balance are reciprocally linked to those that control reproduction; these mechanisms optimize reproductive success under fluctuating metabolic conditions. Mechanisms regulating energy balance involve complex interactions between genetic, environmental, and behavioral factors. The major driving force behind obesity in modern society is overeating, which is largely coded in genes that are responsible for appetite and satiety regulation. About fifty genes that regulate satiety in humans have been reported; fat mass and obesity-associated gene (FTO) and melanocortin 4 receptor (MC4R) are the two best-known examples (Table 1). MC4R codes for a protein that is mainly found in the hypothalamus, an area responsible for controlling appetite and satiety.29 The FTO gene has been reported to demethylate mRNA N6-methyladenosine (m6A) in mammalian cells.30 FTO-depleted cells exhibit higher levels of m6A than control cells; the demethylase activity of FTO protein is required for differentiation of preadipocytes.31 Indeed, the discovery of two missense mutations with potentially detrimental effects on the functionality of the methylation protein FTO, as well as a genetic variant of the same protein that is associated with altered semen quality, suggests that aberrant demethylation of mRNA is a factor involved in reduced male fertility.32 In addition to low satiety, FTO risk variants can also affect food preferences; FTO carriers tend to overeat and prefer high-sugar and high-fat foods. For these reasons, it is essential to adapt the dietary intervention in order to modulate the impact of genetic variations, which can also induce alteration in sperm parameters due to disturbance of the energy balance. The crucial role of lipid homoeostasis and energy balance in endocrine regulation of spermatogenesis is well known. Thus, genes of the spermatozoon membrane structure represent a logical target for mutational analysis in infertile males. Cholesterol and lipid homoeostasis are important for male fecundity.33 It is reported that 65% of infertile men show hypercholesterolemia [Downloaded free from http://www.ajandrology.com on Tuesday, January 26, 2021, IP: 134.157.254.96]
and/or triglyceridemia.34 Moreover, Ergün et al.35 (2007) reported that increased very-low-density lipoprotein (VLDL) impaired seminal parameters, and that increased triglycerides may have deleterious effects on spermatogenesis. In addition, Schisterman et al.36 reported that lipid concentrations may affect semen parameters, specifically sperm head morphology, highlighting the importance of cholesterol and lipid homoeostasis for male fecundity. Lipid components of the spermatozoon have an important role in the functional activity of this cell.
Apolipoprotein E (APOE) has a central role in lipid transport by mediating the binding of lipoprotein particles to their receptors.37 APOE is a constituent apoprotein of VLDL, intermediate-density lipoprotein, high-density lipoprotein (HDL), and chylomicron particles.38 Three important polymorphisms have been discovered in the human APOE gene. These co-dominant alleles, designated ε2, ε3, and ε4, give rise to functionally distinct proteins, namely, APOE2, APOE3, and APOE4.39 APOE polymorphisms have also been found to affect male fertility and cause marked differences in reproductive efficiency.40 Setarehbadi et al.37 found significant differences in the distribution of APOE allelic combinations between fertile men and infertile men, with a higher percentage of fertile males possessing the ε3/ε3 genotype. APOE genotypes may be linked to differences in the efficacy of the expressed APOE isoforms in promoting sperm maturation during epididymal transit.37 Diet is the major reason for variation in lipid metabolism in human populations. Several recent studies have demonstrated that a specific dietary intervention may elicit extraordinary effects in certain genetic subgroups of patients.41 The ability of dietary intervention to improve plasma lipoprotein– lipid profiles varies greatly among individuals with different APOE genotypes. Ordovas and Galluzzi42 focused on the APOE genotype and dietary response in a comprehensive review of 27 studies. In general, the ε4 allele appears to be the most responsive to a low-fat and low-cholesterol dietary intervention; however, it may not be the most responsive to changes in other aspects of the diet. For example, subjects carrying the ε2 allele had the greatest change in total and LDL cholesterol in response to wheat- or oat-bran supplementation. Plasma lipid levels in subjects carrying the APOE2 allele show a more favorable response to tea drinking, and possibly to a fruit-and-vegetable
diet. A long-term increase in dietary soluble fiber does not affect fat metabolism after meals in subjects with the APOE4 allele; however, it does enhance fat absorption in subjects with the APOE3/3 phenotype.
Hyperinsulinemia and hyperglycemia are common occurrences in obese individuals, and are constant confounding factors in many rodent studies of male obesity.43 Both hyperinsulinemia and hyperglycemia have been shown to have an inhibitory effect on sperm quantity and quality, and, therefore, may contribute to the reduced fertility seen in obese men.44 One of the gene variants that plays an important role in the development of type 2 diabetes mellitus in individuals with metabolic syndrome is transcription factor 7-like 2 (TCF7L2) gene (Table 1). Variants of this gene, such as rs12255372 and rs7903146, have been consistently shown to raise the genetic risk of β-cell dysfunction and development of type 2 diabetes.45 Increased ROS and sperm DNA damage are also seen in diabetic patients with commonly altered markers of sperm function. High circulating levels of insulin are suggested as one possible mechanism for the above effects, with increased insulin reducing the production of sex hormone-binding globulin (SHBG).44,46 The decreased levels of SHBG to sustain homoeostatic levels of testosterone could contribute to the decreased levels of testosterone and reduced sperm counts seen in these patients.
Genetic variants that affect the one-carbon metabolic pathway and their influence on male fertility
Male fertility depends on the normal process of spermatogenesis which is itself a complex process regulated by a number of genes involved in growth, differentiation, apoptosis, and DNA damage. This complex pathway is associated with another important pathway, one-carbon metabolism. Folate and the normal activity of one-carbon metabolic enzymes are central to nucleotide synthesis, methylation, and maintenance of genomic integrity, as well as protection from DNA damage. The key enzymes implicated in these metabolic pathways are methylene-tetrahydrofolate reductase (MTHFR), methionine synthase (MTR), and methionine synthase reductase (MTRR) (Table 2).47 Polymorphisms of important genes in the one-carbon metabolic pathway that affect several physiological processes also have an impact on spermatogenesis, and may directly or indirectly influence the quality of sperm parameters and male infertility.48 Homocysteine
Table 1: Summary of genetic variants involved in energy balance and homoeostatic lipid metabolism, and their influence on male fertility Gene
symbol Gene name SNPs Gene consequence Phenotype impact Study Protein function Dietary recommendation based on FTO FTO, alpha-
ketoglutarate- dependent dioxygenase
rs9939609 Intron variant Obesity risk Type 2 diabetes risk Low fat oxidation rates Higher weight and abdominal
circumference
32,104 RNA demethylase that mediates oxidative demethylation of different RNA species Acts as a regulator of fat
mass, adipogenesis, and energy homoeostasis Genetic profile matched to low-fat, low-carbohydrate, Mediterranean or balanced diet, including genetic risks for metabolic health factors (e.g., blood sugar, lipids) rs1558902 Intron variant rs7193144 Intron variant MC4R Melanocortin 4 receptor rs17782313 Intergenic variant
Increased appetite and decreased satiety
Overweight
Autosomal dominant obesity Higher BMI
29,105 Plays a central role as a leptin-targeted neural circuit in energy homoeostasis and somatic growth
APOE Apolipoprotein E rs429358 Missense variant
Hypertriglyceridemia Hyperlipoproteinemia Cardiovascular risk Atherosclerosis
Increased risk factor for male infertility
37 Transports lipoproteins, vitamins, and cholesterol, particularly in the brain rs7412 Missense variant TCF7L2 Transcription
factor 7-like 2 rs12255372 Intron variant Increased type 2 diabetes riskReduction of insulin sensitivity Fasting proinsulin
106,107 Implicated in blood glucose homoeostasis rs7903146 Intron variant
Asian Journal of Andrology
Impact of nutritional SNPs on male infertility
S Mahbouli et al 4
is a sulfur-containing amino acid, which is an intermediate product in the metabolism of the amino acid methionine. The homocysteine, folate, and methyl group metabolic pathways are linked processes. The choline, methionine, and folate metabolic pathways interact at the point where homocysteine is converted to methionine (Figure 1). Homocysteine can be methylated to form methionine49 by two parallel pathways, both of which decrease homocysteine concentrations.50 The alternative pathway for methylation of homocysteine to form methionine is catalyzed by betaine-homocysteine S-methyltransferase (BHMT).51 Betaine, derived from dietary choline by the action of choline dehydrogenase (CHDH), is the methyl group donor in this reaction, and supplemental oral betaine can lower plasma homocysteine concentrations.52 Dysfunction of folate metabolism pathways due to insufficient dietary folate intake, vitamin B9 deficiency,53 and genetic variations that impair the activity of enzymes involved in these processes will lead to a reduction in the conversion of homocysteine to methionine.54,55 This may lead to hyperhomocysteinemia associated with an increased risk of cardiovascular disease and disturbed DNA synthesis and/or DNA methylation reactions, causing DNA mutations and altered gene expression.
A few studies have assessed an association between male infertility and variants in key genes, MTR, MTRR, and MTHFR, for enzymes involved in methylation and homocysteine metabolism, with mixed
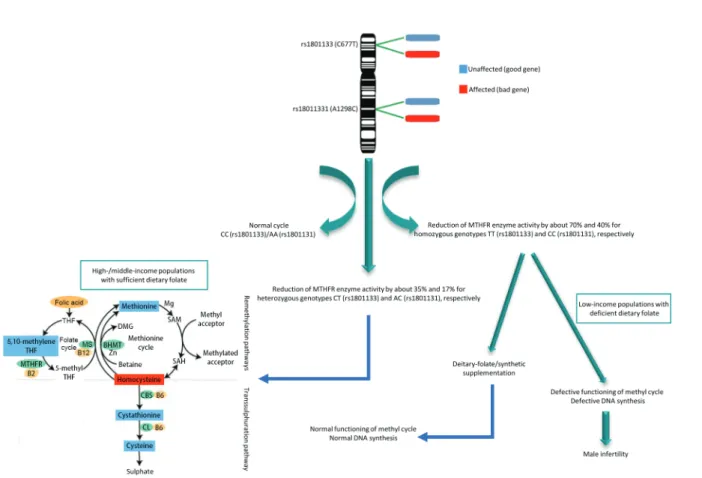
results.48,56 MTHFR is one of the main regulatory enzymes involved in folate metabolism, DNA synthesis, and remethylation reactions. Moreover, MTHFR, which performs a key function in the metabolism of folate and homocysteine, is potentially one of the candidates for genetic vulnerability to spermatogenic failure. Carriers of the MTHFR rs1801133 (C677T) variant have been shown to have decreased activity of MTHFR enzyme – by 35% in the presence of heterozygosis and 70% in homozygosis.57 Similarly, the MTHFR rs1801131 (A1298C) polymorphism has been shown to be associated with lower enzymatic activity in vitro, but to a lesser degree than MTHFR rs1801133,58 with a resultant increase in homocysteine levels.59 Some authors have described a statistically significant correlation between MTHFR polymorphisms and human male infertility.60–62 Possible negative effects of the MTHFR rs1801133 mutation on male fertility may be caused by alteration of the expression of genes involved in spermatogenesis induced by undermethylation,63 or spermatozoa may be damaged by higher production of ROS metabolites, causing DNA damage (Figure 2) and reduced sperm counts.64,65 Significant experimental data show that the chief enzymes of the folate metabolism cycle are vital to male spermatogenesis.60
Genetic variants involved in vitamin D metabolism and their influence on male fertility
The male reproductive tract is one of the sites where vitamin D is metabolized. Vitamin D receptor (VDR) expression in various reproductive tissues, such as the smooth muscles of the epididymis, spermatogonia, Sertoli cells, and spermatozoa (especially the midpiece and nucleus), shows that it plays a crucial role in reproduction, and, therefore, fertility (Table 3).66 The role of vitamin D in the modulation of testicular functions, including hormone production and spermatogenesis, has been investigated in animals and humans.67 Experimental studies support a beneficial effect of vitamin D on male fertility, by modulating hormone production through genomic and nongenomic actions, and, particularly, by improving semen quality, essentially through nongenomic actions.68 Indeed, vitamin D seems to contribute to the modulation of the bioavailable rather than total testosterone. Moreover, although an increased prevalence or risk of testosterone deficiency was reported in men with vitamin D deficiency in observational studies, most interventional studies demonstrated a lack of effect of vitamin D supplementation on circulating levels of testosterone. The most consistent effect of vitamin D was reported for
Table 2: Summary of genetic variants involved in folate metabolism, and their influence on male fertility Gene
symbol
Gene name SNPs Gene consequence
Phenotype impact Study Protein function Dietary recommendation based on
MTHFR Methylene-tetrahydrofolate
reductase rs1801133 Missense variant Lower folate statusIdiopathic male infertility Homocystinuria
Reduced activity of MTHFR
57,108 Converts MeTHF to
MTHF Genetic profile matched to folic acid-fortified foods and vitamin B-supplemented diet, including genetic risks of higher blood homocysteine rs1801131 Missense variant MTRR 5-methyl-tetrahydrofolate- homocysteine methyltransferase reductase
rs1801394 Missense variant Homocystinuria
Reduced activity of MTRR enzyme Hyperhomocysteinemia 109 Regenerates functional methionine synthase via reductive methylation BHMT Betaine–homocysteine
S-methyltransferase rs7356530 Intron variant Increased homocysteine levels in the blood 110 Converts betaine and homocysteine to dimethylglycine and methionine
CHDH Choline dehydrogenase rs12676 Missense variant Choline deficiency Changes in human sperm
cell function
111 Involved in step 1 of the subpathway that synthesizes betaine aldehyde from choline SNPs: single-nucleotide polymorphisms; MeTHF: 5,10-methylenetetrahydrofolate; MTHF: 5-methlytetrahydrofolate
Figure 1: Choline metabolism and its links to methionine and folate
metabolism. The pathways described are all present in the liver, with other tissues having one or more of these pathways. B12: cobalamin; BHMT: betaine-homocysteine S-methyltransferase; CHDH: choline dehydrogenase; DMG: dimethylglycine; MTHFD: methylene-tetrahydrofolate dehydrogenase; MTHFR: methylene-tetrahydrofolate reductase; PEMT: phosphatidylethanolamine N-methyltransferase; THF: tetrahydrofolate.
semen quality.69 Vitamin D has been shown to increase sperm motility by increasing intracellular calcium concentrations in spermatozoa through the VDR. We conclude that VDR polymorphism may play a major role in male factor infertility, either directly or indirectly, by reducing the effects of vitamin D.70 Four SNPs are associated with changes in VDR activity, one of which is the rs2228570 variant (Table 3). Furthermore, one SNP within the GC gene is associated with reduced levels of vitamin D. Studies have shown that men with higher dietary and supplemental intake of vitamin D may produce sperm with less DNA damage.71
Genetic variants that affect antioxidant defense and their influence on male fertility
Extensive research suggests that oxidative stress may be an important cause of male infertility, and that the pathology of infertility in
30%–80% of infertile men may be oxidative damage to spermatozoa.72 This is primarily due to DNA and cell membrane damage; however, little is known about the genetic causes underlying suboptimal functioning of the seminal enzymatic antioxidant system. Oxidative stress induces sperm DNA damage,73 a reduction in sperm motility,74 a decline in sperm’s fertilizing ability,75 a reduction in membrane fluidity, apoptosis,27,76 and defective sperm membrane integrity by the way of lipid peroxidation.77 The presence of ROS in seminal plasma is normally balanced by homoeostatic antioxidant systems that facilitate an appropriate level of ROS required for normal physiological processes, such as sperm capacitation, hyperactivation, acrosome reaction, and sperm–oocyte fusion.78 Semen is shown to possess large amounts of antioxidants to counterbalance the effects of ROS, thereby protecting mature spermatozoa from oxidative damage. The antioxidants in
Table 3: Summary of genetic variants involved in vitamin D metabolism, and their influence on male fertility Gene
symbol
Gene name SNPs Gene consequence
Phenotype impact
Study Protein function Dietary recommendation based on
VDR Vitamin D
receptor rs2228570 Start-lost Deficient in vitamin D 112 Plays a key role in the absorption of calcium from the gut, which is required for healthy bone formation, muscle, and heart activity, as well as numerous other cell functions
Genetic predisposition to food and nutrient needs profile and sensitivity to vitamin deficiency GC GC Vitamin D binding protein rs2282679 Intron variant Associated with reduced levels of vitamin D
113 Responsible for binding with the bioactive form of vitamin D, calcitriol, and shuttling it through the circulatory system into tissue and then presenting it to the VDR to allow its binding SNPs: single-nucleotide polymorphisms; VDR: vitamin D receptor
Figure 2: Effects of MTHFR polymorphisms on male fertility. The MTHFR gene, located on the short arm of chromosome 1 (1p36.3), is composed of 11 exons.
It possesses 14 common or rare single-nucleotide polymorphisms that are associated with enzymatic deficiency. Among them, rs1801133 (C677T) and rs1801131 (A1298C) are the most reported to possibly reduce MTHFR activity to various degrees. Reduced enzymatic activity due to MTHFR polymorphisms is considered a risk factor for many diseases, including infertility. B2: riboflavin; B6: pyridoxine; B12: cobalamin; BHMT: betaine-homocysteine methyltransferase; CBS: cystathionine-β-synthase; CL: cystathionine-γ-lyase; DMG: dimethylglycine; MS: methionine synthase; MTHFR: methylenetetrahydrofolate reductase; SAM: S-adenosyl methionine; SAH: A-adenosyl homocysteine; THF: tetrahydrofolate.
Asian Journal of Andrology
Impact of nutritional SNPs on male infertility
S Mahbouli et al 6
semen are present both in spermatozoa and the seminal plasma; however, they are most abundant in the latter because the amount of spermatozoan cytoplasm is small, thereby limiting antioxidant defense within the cells.79 Seminal plasma is enriched with both enzymatic antioxidants, such as superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GPx), and glutathione S-transferase (GST), and nonenzymatic antioxidants, such as glutathione, vitamin A, vitamin C, vitamin E, and coenzyme Q10.80 SOD2 is an enzyme that catalyzes the detoxification of superoxide radicals in the mitochondrion. CAT has the capacity to detoxify hydrogen peroxide (H2O2) by converting it to water (H2O)and oxygen (O2).GPx1 is related to the final electron transporter in mitochondria and neutralizes peroxide radicals into water, whereas GST conjugates toxic electrophiles and other intermediates, with glutathione neutralizing their toxicity. Dietary selenium intake is an important determinant of GPx1 activity, with selenium supplementation leading to an increase in blood GPx1 activity. Large interindividual differences have been observed in the response of GPx1 to selenium supplementation.81 These variations are mainly due to differences in the baseline selenium status; however, interactions between genetic polymorphisms in the GPX1 gene and dietary selenium intake may account for some of the interindividual variations.82
Deficiencies in enzymatic or nonenzymatic antioxidant systems in seminal plasma are widely associated with male infertility as the absence of any of these systems leads to the accumulation of excessive levels of ROS, resulting in impairment of both the structural and functional integrity of spermatozoa.83 Although these enzymes appear to be conserved phylogenetically, intraspecific polymorphisms may still cause changes in their activities, and, therefore, they may be useful in understanding the underlying origins of idiopathic infertility. Genetic variation in one or more enzymes involved in redox balance may induce alteration of antioxidant activity in hypofertile men (Table 3). Paraoxonase 1 (PON1) is one such gene that can have an impact on male infertility. PON proteins, localized in the seminiferous tubules and spermatozoa, have been implicated in the pathogenesis of male infertility.84,85 Impaired oxidative stress regulation in the seminal plasma of patients with abnormal sperm parameters has been proposed to be the result of decreased PON1 activity.86 PON possesses antioxidant properties and protects cells against oxidative stress.87 Lazaros et al.88 showed, for the first time, the association between PON gene variants and semen concentration and motility. Glutathione S-transferase mu 1 (GSTM1) and glutathione S-transferase theta 1 (GSTT1) have the highest frequency of polymorphisms among all glutathione S-transferase genes (Table 4). They are characterized by deletion polymorphisms manifested in the absence of enzymatic activity of relevant proteins, and with a frequency of 42%–60% for GSTM1 and 13%–26% for GSTT1 in Caucasian populations.89–91 Genetically determined imbalance in the detoxification system, due to reduced activity of glutathione system enzymes, can be the cause of various pathological processes, including infertility.92
Nitric oxide, produced by nitric oxide synthase 3 (NOS3), is considered to be an important mediator of oxidative stress in testicular tissue.93 Studies have shown that NOS is involved in sperm motility, capacitation, and acrosome reaction.94 Nitric oxide, as one of the most potent free radicals of nitrogen, reacts rapidly with superoxide (O2−) to form highly toxic peroxynitrite (ONOO−). Both superoxide and peroxynitrite have the ability to damage DNA directly. Excessive concentrations of nitric oxide in the semen of asthenozoospermic patients have overall negative effects on the kinetic characteristics of spermatozoa and, consequently, reduce sperm motility and sperm DNA
integrity.95 Nitric oxide concentration has been found to be significantly higher in the seminal plasma of some infertile males than in that of healthy males.96 Associations between NOS3 gene polymorphisms and male infertility have been reported.97 Multivariate logistic regression analyses revealed that carriers of the NOS3 rs1799983 variant among GT heterozygotes were associated with a marginally significant increase in the risk of male infertility.98 In the dominant model, combined rs1799983 genotypes (GT/TT) were associated with a significant 34% increase in the risk of male infertility. Buldreghini et al.99 (2010) showed that the T allele of NOS3 rs1799983 polymorphism (Table 4) contributed to poor sperm motility. Common genetic variants affecting uptake, distribution, transport, or metabolism of dietary antioxidants have been linked to variation in antioxidant serum levels and response to supplementation.100,101 Elucidating the relationship between common genetic variants and antioxidant status may have important health implications through identification of individuals and subgroups that benefit the most from dietary intervention or supplementation with antioxidants.
CONCLUSIONS
This paper provides an overview of the current science linking genetic variants to nutritional or supplemental needs with a focus on direct and indirect factors that influence male infertility. This concept is nutrigenomic which assesses interaction between nutrients and gene expression. It is a preliminary review of the potential impacts of various genetic polymorphisms, associated with efficiency of energy expenditure, antioxidant defense, and energetic metabolism, that influence male fertility. It is conceivable that the effects of some genes on fertility phenotypes may be nutrient sensitive. Indeed, diet composition may modulate gene expression through complex transcriptional mechanisms as well as more downstream processes involving the gene products.
Following the present review, a clinical trial is being set up to study the effect of a personalized lifestyle and nutrition program based on a specific set of genetic variants that we identify in this paper. The study was approved by the French ethics committee and registered in ClinicalTrials.gov with reference number NCT03475199.
With the advent of personalized medicine, identification of polymorphisms related to the reproductive function in men and elucidation of their functional importance remain an important area of research. Indeed, epigenetic modifications play a potential role in spermatogenesis via regulation of molecular pathways to maintain testicular homoeostasis. The best-known epigenetic process is DNA methylation. A recent genome-wide study has shown that aberrant DNA methylation is imprinted, and developmental genes may have a role in male infertility.102 In addition to genetic factors, environmental genotoxins, endocrine disruptors, and micronutrient deficiency play an important role in the increasing rates of human infertility. These factors may have deleterious effects on human reproductive health through numerous mechanisms and may also explain some cases of idiopathic infertility in men, especially when the male factor in infertility problems of couples has been neglected for a long time. Emerging observations support the conclusion that parental influences begin before conception and compel us to further explore preconception pathways by which parents contribute more than genetic material to offspring.103 There is now clear evidence that beyond genetic alterations alone, there is an epigenetic transmission from the father to his offspring. In fact, paternal smoking, age, and occupational chemical exposure are well known to be linked to increased risk of cancer and neurological disorders in children. It is less appreciated that the father’s body mass has a greater [Downloaded free from http://www.ajandrology.com on Tuesday, January 26, 2021, IP: 134.157.254.96]
impact than the mother’s on prepubertal child’s body fat and metabolic measures. In addition to sperm DNA damage, in some instances, there is accumulating evidence for pathways of parental transgenerational epigenetic effects, attributable to sperm and seminal fluid, that transmit the effects of environmental exposure to the next generation.
Ultimately, once pathways are defined and prioritized according to their importance for health outcomes, it will be possible to define how prospective parents can alter their lifestyles and food choices and adopt interventions to protect children from adverse outcomes. However, further research is required to fully clarify the potential effect of specific SNPs involved in metabolism on male infertility.
AUTHOR CONTRIBUTIONS
SM reviewed the literature, collected data, and wrote the manuscript. CD, YE, EL, and RL collaborated in writing, revising, and editing the manuscript. All authors read and approved the final manuscript.
COMPETING INTERESTS
All authors declare no competing interests.
REFERENCES
1 Levine H, Jørgensen N, Martino-Andrade A, Mendiola J, Weksler-Derri D, et al. Temporal trends in sperm count: a systematic review and meta-regression analysis.
Hum Reprod Update 2017; 23: 646–59.
2 Tang Q, Chen Y, Wu W, Ding H, Xia Y, et al. Idiopathic male infertility and polymorphisms in the DNA methyltransferase genes involved in epigenetic marking.
Sci Rep 2017; 7: 11219.
3 García Rodríguez A, de la Casa M, Johnston S, Gosálvez J, Roy R. Association of polymorphisms in genes coding for antioxidant enzymes and human male infertility.
Ann Hum Genet 2019; 83: 63–72.
4 Laqqan M, Hammadeh ME. Aberrations in sperm DNA methylation patterns of males suffering from reduced fecundity. Andrologia 2018; 50: e12913.
5 Horne J, Madill J, O’Connor C, Shelley J, Gilliland J. A systematic review of genetic testing and lifestyle behaviour change: are we using high-quality genetic interventions and considering behaviour change theory? Lifestyle Genom 2018; 11: 49–63. 6 Fenech M, Ferguson LR. Vitamins/minerals and genomic stability in humans. Mutat
Res 2001; 475: 1–6.
7 Sermondade N, Faure C, Fezeu L, Levy R, Czernichow S. Obesity and increased risk for oligozoospermia and azoospermia. Arch Intern Med 2012; 172: 440–2. 8 Dupont C, Faure C, Sermondade N, Boubaya M, Eustache F, et al. Obesity leads
to higher risk of sperm DNA damage in infertile patients. Asian J Androl 2013; 15: 622–5.
9 Palmer NO, Bakos HW, Fullston T, Lane M. Impact of obesity on male fertility, sperm function and molecular composition. Spermatogenesis 2012; 2: 253–63. 10 Morrison CD, Brannigan RE. Metabolic syndrome and infertility in men. Best Pract
Res Clin Obstet Gynaecol 2015; 29: 507–15.
11 Dupont J, Pollet-Villard X, Reverchon M, Mellouk N, Levy R. Adipokines in human reproduction. Horm Mol Biol Clin Investig 2015; 24: 11–24.
12 Giahi L, Mohammadmoradi S, Javidan A, Sadeghi MR. Nutritional modifications in male infertility: a systematic review covering 2 decades. Nutr Rev 2016; 74: 118–30. 13 Lobaccaro JM, Brugnon F, Volle DH, Baron S. Lipid metabolism and infertility: is
there a link? Clin Lipidol 2012; 7: 485–8.
14 Safarinejad MR. Effect of omega-3 polyunsaturated fatty acid supplementation on
Table 4: Summary of genetic variants involved in redox balance, and their influence on male fertility Gene
symbol Gene name SNPs Gene consequence Phenotype impact Study Protein function Dietary recommendation based on SOD2 Superoxide
dismutase 2
rs4880 Missense variant
Idiopathic male infertility Low pregnancy rates in IVF Sperm concentration Sperm motility DNA fragmentation
114 Plays an important role in protecting spermatozoa from oxidative damage Genetic profile and sensitivity to antioxidative stress system matched to antioxidant supplementation, including genetic risks for oxidative stress.
CAT Catalase rs1001179 Regulatory region variant
Lower susceptibility to male infertility
Idiopathic male infertility Higher distribution of fat
115 Plays a defensive role against oxidative stress.
Detoxifies both intracellular and extracellular H2O2 to water and oxygen
PON1 Paraoxonase 1 rs662 Missense variant
Coronary artery disease Decreased sperm motility Decreased semen quality
Decrease in antioxidant PON1 activity Increase in lipid peroxidation Higher oxidative stress levels
116 Removes harmful oxidized lipids Protects against the development
of atherosclerosis GPX1 Glutathione peroxidase 1 rs1050450 Regulatory region variant
Increased risk of damage caused by oxidative stress
Risk of male infertility
117 Catalyzes the reduction of organic hydroperoxides and H2O2 by glutathione, thereby protecting cells against oxidative damage
GSTT1 Glutathione S-transferase theta 1
Null alleles Null deletion Decreased detoxification Susceptibility factors of male
infertility
Susceptibility to spermatogenesis
118 Catalyzes the conjugation of glutathione to a wide range of potential toxins as the first step in detoxification
GSTM1 Glutathione
S-transferase mu 1
Null alleles Null deletion Idiopathic male infertility Oligoazoospermia Varicocele
119 Catalyzes the reaction of glutathione, a reaction that is sometimes a first step in a detoxification process leading to mercapturic acid formation
GSTP1 Glutathione S-transferase pi 1
rs1695 Missense variant
Development of male factor infertility High oxidative stress
92 Neutralizes xenobiotic reactive oxygen species on other molecules
NOS3 Nitric oxide
synthase 3 rs1799983 Missense variant Increased risk of male infertilityHigher levels of sperm DNA fragmentation
Oxidative stress Sperm DNA damage Poor sperm motility
93 Plays a key role in regulation of nitric oxide
Asian Journal of Andrology
Impact of nutritional SNPs on male infertility
S Mahbouli et al 8
semen profile and enzymatic anti-oxidant capacity of seminal plasma in infertile men with idiopathic oligoasthenoteratospermia: a double-blind, placebo-controlled, randomised study. Andrologia 2011; 43: 38–47.
15 Conquer JA, Martin JB, Tummon I, Watson L, Tekpetey F. Effect of DHA supplementation on DHA status and sperm motility in asthenozoospermic males.
Lipids 2000; 35: 149–54.
16 Hosseini B, Djafarian K. Dietary nutrients and male infertility: review of current evidence. Galen Med J 2015; 4: 123–9.
17 Chavarro JE, Toth TL, Wright DL, Meeker JD, Hauser R. Body mass index in relation to semen quality, sperm DNA integrity and serum reproductive hormone levels among men attending an infertility clinic. Fertil Steril 2010; 93: 2222–31.
18 Sermondade N, Faure C, Fezeu L, Shayeb AG, Bonde JP, et al. BMI in relation to sperm count: an updated systematic review and collaborative meta-analysis. Hum
Reprod Update 2013; 19: 221–31.
19 Kaukua J, Pekkarinen T, Sane T, Mustajoki P. Sex hormones and sexual function in obese men losing weight. Obes Res 2003; 11: 689–94.
20 Håkonsen LB, Thulstrup AM, Aggerholm AS, Olsen J, Bonde JP, et al. Does weight loss improve semen quality and reproductive hormones? Results from a cohort of severely obese men. Reprod Health 2011; 8: 24.
21 Ilacqua A, Izzo G, Emerenziani GP, Baldari C, Aversa A. Lifestyle and fertility: the influence of stress and quality of life on male fertility. Reprod Biol Endocrinol RBE 2018; 16: 115.
22 Gameiro S, Boivin J, Dancet E, Emery M, Thorn P, et al. Qualitative research in the ESHRE Guideline “Routine psychosocial care in infertility and medically assisted reproduction – A guide for staff.” Hum Reprod Oxf Engl 2016; 31: 1928–9. 23 Connor KL, Vickers MH, Beltrand J, Meaney MJ, Sloboda DM. Nature, nurture or
nutrition? Impact of maternal nutrition on maternal care, offspring development and reproductive function. J Physiol 2012; 590: 2167–80.
24 Mendiola J, Torres-Cantero AM, Moreno-Grau JM, Ten J, Roca M, et al. Food intake and its relationship with semen quality: a case-control study. Fertil Steril 2009; 91: 812–8.
25 Smits RM, Mackenzie-Proctor R, Yazdani A, Stankiewicz MT, Jordan V, et al. Antioxidants for male subfertility. Cochrane Database Syst Rev 2019; 3: CD007411. 26 Kumar N, Singh AK. Reactive oxygen species in seminal plasma as a cause of male
infertility. J Gynecol Obstet Hum Reprod 2018; 47: 565–72.
27 Aitken RJ. Oxidative stress and the etiology of male infertility. J Assist Reprod Genet 2016; 33: 1691–2.
28 Gharagozloo P, Aitken RJ. The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Hum Reprod 2011; 26: 1628–40. 29 Loos RJF, Lindgren CM, Li S, Wheeler E, Zhao JH, et al. Common variants near
MC4R are associated with fat mass, weight and risk of obesity. Nat Genet 2008; 40: 768–75.
30 Jia G, Fu Y, Zhao X, Dai Q, Zheng G, et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol 2011; 7: 885–7. 31 Zhang M, Zhang Y, Ma J, Guo F, Cao Q, et al. The demethylase activity of FTO (fat
mass and obesity associated protein) is required for preadipocyte differentiation.
PLoS One 2015; 10: e0133788.
32 Landfors M, Nakken S, Fusser M, Dahl JA, Klungland A, et al. Sequencing of FTO and ALKBH5 in men undergoing infertility work-up identifies an infertility-associated variant and two missense mutations. Fertil Steril 2016; 105: 1170–9.e5. 33 Maqdasy S, Baptissart M, Vega A, Baron S, Lobaccaro JM, et al. Cholesterol and male
fertility: what about orphans and adopted? Mol Cell Endocrinol 2013; 368: 30–46. 34 Ramírez-Torres MA, Carrera A, Zambrana M. High incidence of hyperestrogenemia
and dyslipidemia in a group of infertile men. Ginecol Obstet Mex 2000; 68: 224–9. 35 Ergün A, Köse SK, Aydos K, Ata A, Avci A. Correlation of seminal parameters with
serum lipid profile and sex hormones. Arch Androl 2007; 53: 21–3.
36 Schisterman EF, Mumford SL, Browne RW, Barr DB, Chen Z, et al. Lipid concentrations and couple fecundity: the LIFE study. J Clin Endocrinol Metab 2014; 99: 2786–94.
37 Setarehbadi R, Vatannejad A, Vaisi-Raygani A, Amiri I, Esfahani M, et al. Apolipoprotein E genotypes of fertile and infertile men. Syst Biol Reprod Med 2012; 58: 263–7.
38 Hopewell JC, Seedorf U, Farrall M, Parish S, Kyriakou T, et al. Impact of lipoprotein(a) levels and apolipoprotein(a) isoform size on risk of coronary heart disease. J Intern
Med 2014; 276: 260–8.
39 Mahley RW, Rall SC. Apolipoprotein E: far more than a lipid transport protein. Annu
Rev Genomics Hum Genet 2000; 1: 507–37.
40 Corbo RM, Scacchi R, Cresta M. Differential reproductive efficiency associated with common apolipoprotein E alleles in postreproductive-aged subjects. Fertil
Steril 2004; 81: 104–7.
41 Kabisch S. Nutrigenetic effects in metabolic syndrome – a cornerstone for individualized therapy. J Diabetes Complications 2019; 33: 193–4.
42 Ordovas JM, Galluzzi JR. Genetic predictors of plasma lipid response to diet intervention. Curr Atheroscler Rep 1999; 1: 196–203.
43 Ng SF, Lin RC, Laybutt DR, Barres R, Owens JA, et al. Chronic high-fat diet in fathers programs β-cell dysfunction in female rat offspring. Nature 2010; 467: 963–6. 44 Kasturi SS, Tannir J, Brannigan RE. The metabolic syndrome and male infertility.
J Androl 2008; 29: 251–9.
45 Katsoulis K, Paschou SA, Hatzi E, Tigas S, Georgiou I, et al. TCF7L2 gene variants predispose to the development of type 2 diabetes mellitus among individuals with metabolic syndrome. Horm Athens Greece 2018; 17: 359–65.
46 La Vignera S, Condorelli RA, Vicari E, D’Agata R, Salemi M, et al. High levels of lipid peroxidation in semen of diabetic patients. Andrologia 2012; 44 Suppl 1: 565–70. 47 Murphy LE, Mills JL, Molloy AM, Qian C, Carter TC, et al. Folate and vitamin B12
in idiopathic male infertility. Asian J Androl 2011; 13: 856–61.
48 Liu K, Zhao R, Shen M, Ye J, Li X, et al. Role of genetic mutations in folate-related enzyme genes on male infertility. Sci Rep 2015; 5: 15548.
49 Finkelstein JD. Pathways and regulation of homocysteine metabolism in mammals.
Semin Thromb Hemost 2000; 26: 219–25.
50 Olthof MR, van Vliet T, Boelsma E, Verhoef P. Low dose betaine supplementation leads to immediate and long term lowering of plasma homocysteine in healthy men and women. J Nutr 2003; 133: 4135–8.
51 Sunden SL, Renduchintala MS, Park EI, Miklasz SD, Garrow TA. Betaine-homocysteine methyltransferase expression in porcine and human tissues and chromosomal localization of the human gene. Arch Biochem Biophys 1997; 345: 171–4.
52 Steenge GR, Verhoef P, Katan MB. Betaine supplementation lowers plasma homocysteine in healthy men and women. J Nutr 2003; 133: 1291–5. 53 Shelnutt KP, Kauwell GP, Chapman CM, Gregory JF, Maneval DR, et al. Folate status
response to controlled folate intake is affected by the methylenetetrahydrofolate reductase 677C-->T polymorphism in young women. J Nutr 2003; 133: 4107–11. 54 Jacques PF, Bostom AG, Wilson PW, Rich S, Rosenberg IH, et al. Determinants of
plasma total homocysteine concentration in the Framingham Offspring cohort. Am
J Clin Nutr 2001; 73: 613–21.
55 Watkins D, Ru M, Hwang HY, Kim CD, Murray A, et al. Hyperhomocysteinemia due to methionine synthase deficiency, cblG: structure of the MTR gene, genotype diversity, and recognition of a common mutation, P1173L. Am J Hum Genet 2002; 71: 143–53.
56 Shi TL, Wu Y, Li Y, Chen ZF, Ma YN, et al. The relevance of MTHFR C677T, A1298C, and MTRR A66G polymorphisms with response to male infertility in Asians: a meta-analysis. Medicine (Baltimore) 2019; 98: e14283.
57 Gong M, Dong W, He T, Shi Z, Huang G, et al. MTHFR 677C>T polymorphism increases the male infertility risk: a meta-analysis involving 26 studies. PLoS One 2015; 10: e0121147.
58 Wan L, Li Y, Zhang Z, Sun Z, He Y, et al. Methylenetetrahydrofolate reductase and psychiatric diseases. Transl Psychiatry 2018; 8: 242.
59 Li SS, Li J, Xiao Z, Ren AG, Jin L. Prospective study of MTHFR genetic polymorphisms as a possible etiology of male infertility. Genet Mol Res GMR 2014; 13: 6367–74. 60 Chan D, Cushnie DW, Neaga OR, Lawrance AK, Rozen R, et al. Strain-specific defects in testicular development and sperm epigenetic patterns in 5,10-methylenetetrahydrofolate reductase-deficient mice. Endocrinology 2010; 151: 3363–73.
61 Gava MM, Chagas Ede O, Bianco B, Christofolini DM, Pompeo AC, et al. Methylenetetrahydrofolate reductase polymorphisms are related to male infertility in Brazilian men. Genet Test Mol Biomark 2011; 15: 153–7.
62 Wu W, Shen O, Qin Y, Niu X, Lu C, et al. Idiopathic male infertility is strongly associated with aberrant promoter methylation of methylenetetrahydrofolate reductase (MTHFR). PLoS One 2010; 5: e13884.
63 Tüttelmann F, Simoni M, Kliesch S, Ledig S, Dworniczak B, et al. Copy number variants in patients with severe oligozoospermia and Sertoli-cell-only syndrome.
PLoS One 2011; 6: e19426.
64 Ebisch IM, Thomas CM, Peters WH, Braat DD, Steegers-Theunissen RP. The importance of folate, zinc and antioxidants in the pathogenesis and prevention of subfertility. Hum Reprod Update 2007; 13: 163–74.
65 Paracchini V, Garte S, Taioli E. MTHFR C677T polymorphism, GSTM1 deletion and male infertility: a possible suggestion of a gene-gene interaction? Biomark Biochem
Indic Expo Response Susceptibility Chem 2006; 11: 53–60.
66 de Angelis C, Galdiero M, Pivonello C, Garifalos F, Menafra D, et al. The role of vitamin D in male fertility: a focus on the testis. Rev Endocr Metab Disord 2017; 18: 285–305.
67 Zanatta L, Zamoner A, Zanatta AP, Bouraïma-Lelong H, Delalande C, et al. Nongenomic and genomic effects of 1α,25(OH)2 vitamin D3 in rat testis. Life Sci
2011; 89: 515–23.
68 Haussler MR, Jurutka PW, Mizwicki M, Norman AW. Vitamin D receptor (VDR)-mediated actions of 1α,25(OH)2vitamin D3: genomic and non-genomic mechanisms.
Best Pract Res Clin Endocrinol Metab 2011; 25: 543–59.
69 Vladoiu S, Dinu Draganescu D, Botezatu A, Anton G, Oros S, et al. Correlations between polymorphisms of estrogen 1, vitamin D receptors and hormonal profile in infertile men. Acta Endocrinol Buchar 2016; 12: 137–44.
70 Bhakat R, Chandra L, Saxena A, Sarda AK, Krishnamurthy K, et al. Evaluation of metabolic syndrome and vitamin D receptor gene polymorphism in male factor infertility. Indian J Clin Biochem IJCB 2017; 32: 468–72.
71 Hamadé A, Bhanini S, Saade T, Fakih Y, Fakih C, et al. Vitamin D levels in serum, vitamin D receptor polymorphisms and semen quality correlations in Lebanon: a [Downloaded free from http://www.ajandrology.com on Tuesday, January 26, 2021, IP: 134.157.254.96]
pilot cross-sectional study. Univers J Public Health 2014; 2: 118–24.
72 Agarwal A, Prabakaran S, Allamaneni S. What an andrologist/urologist should know about free radicals and why. Urology 2006; 67: 2–8.
73 Santi D, De Vincentis S, Magnani E, Spaggiari G. Impairment of sperm DNA methylation in male infertility: a meta-analytic study. Andrology 2017; 5: 695–703. 74 Kao SH, Chao HT, Chen HW, Hwang TIS, Liao TL, et al. Increase of oxidative stress
in human sperm with lower motility. Fertil Steril 2008; 89: 1183–90.
75 Mostafa T, Rashed LA, Osman I, Marawan M. Seminal plasma oxytocin and oxidative stress levels in infertile men with varicocele. Andrologia 2015; 47: 209–13. 76 Aitken RJ, Smith TB, Jobling MS, Baker MA, De Iuliis GN. Oxidative stress and
male reproductive health. Asian J Androl 2014; 16: 31–8.
77 Saleh RA, Agarwal A, Nada EA, El-Tonsy MH, Sharma RK, et al. Negative effects of increased sperm DNA damage in relation to seminal oxidative stress in men with idiopathic and male factor infertility. Fertil Steril 2003; 79 Suppl 3: 1597–605. 78 Kothari S, Thompson A, Agarwal A, du Plessis SS. Free radicals: their beneficial
and detrimental effects on sperm function. Indian J Exp Biol 2010; 48: 425–35. 79 Shiva M, Gautam AK, Verma Y, Shivgotra V, Doshi H, et al. Association between
sperm quality, oxidative stress, and seminal antioxidant activity. Clin Biochem 2011; 44: 319–24.
80 Bansal AK, Bilaspuri GS. Impacts of oxidative stress and antioxidants on semen functions. Vet Med Int 2010; 2010: 686137.
81 Brown KM, Pickard K, Nicol F, Beckett GJ, Duthie GG, et al. Effects of organic and inorganic selenium supplementation on selenoenzyme activity in blood lymphocytes, granulocytes, platelets and erythrocytes. Clin Sci Lond Engl 1979 2000; 98: 593–9. 82 Miller JC, Thomson CD, Williams SM, van Havre N, Wilkins GT, et al. Influence of the
glutathione peroxidase 1 Pro200Leu polymorphism on the response of glutathione peroxidase activity to selenium supplementation: a randomized controlled trial. Am
J Clin Nutr 2012; 96: 923–31.
83 Venkatesh S, Shamsi MB, Deka D, Saxena V, Kumar R, et al. Clinical implications of oxidative stress & sperm DNA damage in normozoospermic infertile men. Indian
J Med Res 2011; 134: 396–8.
84 Lazaros LA, Xita NV, Hatzi EG, Kaponis AI, Stefos TJ, et al. Association of paraoxonase gene polymorphisms with sperm parameters. J Androl 2011; 32: 394–401. 85 Tavilani H, Fattahi A, Esfahani M, Khodadadi I, Karimi J, et al. Genotype and
phenotype frequencies of paraoxonase 1 in fertile and infertile men. Syst Biol
Reprod Med 2014; 60: 361–6.
86 Verit FF, Verit A, Ciftci H, Erel O, Celik H. Paraoxonase-1 activity in subfertile men and relationship to sperm parameters. J Androl 2009; 30: 183–9.
87 Li HL, Liu DP, Liang CC. Paraoxonase gene polymorphisms, oxidative stress, and diseases. J Mol Med Berl Ger 2003; 81: 766–79.
88 Lazaros L, Xita N, Hatzi E, Kaponis A, Makrydimas G, et al. Phosphatidylethanolamine N-methyltransferase and choline dehydrogenase gene polymorphisms are associated with human sperm concentration. Asian J Androl 2012; 14: 778–83.
89 Aston KI, Krausz C, Laface I, Ruiz-Castané E, Carrell DT. Evaluation of 172 candidate polymorphisms for association with oligozoospermia or azoospermia in a large cohort of men of European descent. Hum Reprod Oxf Engl 2010; 25: 1383–97. 90 Gattás GJ, Kato M, Soares-Vieira JA, Siraque MS, Kohler P, et al. Ethnicity and
glutathione S-transferase (GSTM1/GSTT1) polymorphisms in a Brazilian population.
Braz J Med Biol Res 2004; 37: 451–8.
91 Xiong DK, Chen HH, Ding XP, Zhang SH, Zhang JH. Association of polymorphisms in glutathione S-transferase genes (GSTM1, GSTT1, GSTP1) with idiopathic azoospermia or oligospermia in Sichuan, China. Asian J Androl 2015; 17: 481–6. 92 Safarinejad MR, Dadkhah F, Ali Asgari M, Hosseini SY, Kolahi AA, et al. Glutathione
S-transferase polymorphisms (GSTM1, GSTT1, GSTP1) and male factor infertility risk: a pooled analysis of studies. Urol J 2012; 9: 541–8.
93 Vučić NL, Nikolić ZZ, Vukotić VD, Tomović SM, Vuković II, et al. NOS3 gene variants and male infertility: association of 4a/4b with oligoasthenozoospermia. Andrologia 2018; 50: e12817.
94 Herrero MB, Gagnon C. Nitric oxide: a novel mediator of sperm function. J Androl 2001; 22: 349–56.
95 Balercia G, Moretti S, Vignini A, Magagnini M, Mantero F, et al. Role of nitric oxide concentrations on human sperm motility. J Androl 2004; 25: 245–9.
96 Huang I, Jones J, Khorram O. Human seminal plasma nitric oxide: correlation with sperm morphology and testosterone. Med Sci Monit Int Med J Exp Clin Res 2006; 12: CR103–6.
97 Safarinejad MR, Shafiei N, Safarinejad S. The role of endothelial nitric oxide synthase (eNOS) T-786C, G894T, and 4a/b gene polymorphisms in the risk of idiopathic male infertility. Mol Reprod Dev 2010; 77: 720–7.
98 Yan L, Guo W, Wu S, Liu J, Zhang S, et al. Genetic variants in nitric oxide synthase genes and the risk of male infertility in a Chinese population: a case-control study.
PLoS One 2014; 9: e115190.
99 Buldreghini E, Mahfouz RZ, Vignini A, Mazzanti L, Ricciardo-Lamonica G, et al. Single nucleotide polymorphism (SNP) of the endothelial nitric oxide synthase (eNOS) gene (Glu298Asp variant) in infertile men with asthenozoospermia. J Androl 2010; 31: 482–8.
100 Cahill LE, Fontaine-Bisson B, El-Sohemy A. Functional genetic variants of glutathione S-transferase protect against serum ascorbic acid deficiency. Am J Clin Nutr 2009; 90: 1411–7.
101 Major JM, Yu K, Wheeler W, Zhang H, Cornelis MC, et al. Genome-wide association study identifies common variants associated with circulating vitamin E levels. Hum
Mol Genet 2011; 20: 3876–83.
102 Gunes S, Arslan MA, Hekim GN, Asci R. The role of epigenetics in idiopathic male infertility. J Assist Reprod Genet 2016; 33: 553–69.
103 Soubry A. POHaD: why we should study future fathers. Environ Epigenetics 2018; 4: dvy007.
104 Hunt SC, Stone S, Xin Y, Scherer CA, Magness CL, et al. Association of the FTO gene with BMI. Obes Silver Spring Md 2008; 16: 902–4.
105 Hinney A, Volckmar AL, Knoll N. Melanocortin-4 receptor in energy homeostasis and obesity pathogenesis. Prog Mol Biol Transl Sci 2013; 114: 147–91.
106 Liu XH, Xie CG, An Y, Zhang XX, Wu WB. Meta-analysis of the association between the rs7903146 polymorphism at the TCF7L2 locus and type 2 diabetes mellitus susceptibility. Genet Mol Res GMR 2015; 14: 16856–62.
107 Ding W, Xu L, Zhang L, Han Z, Jiang Q, et al. Meta-analysis of association between TCF7L2 polymorphism rs7903146 and type 2 diabetes mellitus. BMC Med Genet 2018; 19: 38.
108 Irfan M, Ismail M, Azhar Beg M, Shabbir A, Rashid Kayani A, et al. Association of the MTHFR C677T (rs1801133) polymorphism with idiopathic male infertility in a local Pakistani population. Balk J Med Genet BJMG 2016; 19: 51–62. 109 Xue G, Lu CJ, Pan SJ, Zhang YL, Miao H, et al. DNA hypomethylation of CBS promoter
induced by folate deficiency is a potential noninvasive circulating biomarker for colorectal adenocarcinomas. Oncotarget 2017; 8: 51387–401.
110 Ren B, Tian D, Wang L, Han H, Wang W, et al. Association between the BHMT gene rs3733890 polymorphism and the efficacy of oral folate therapy in patients with hyperhomocysteinemia. Ann Hum Genet 2019; 83: 434–44.
111 Johnson AR, Lao S, Wang T, Galanko JA, Zeisel SH. Choline dehydrogenase polymorphism rs12676 is a functional variation and is associated with changes in human sperm cell function. PLoS One 2012; 7: e36047.
112 García-Martín E, Agúndez JAG, Martínez C, Benito-León J, Millán-Pascual J, et al.
Vitamin D3 receptor (VDR) gene rs2228570 (Fok1) and rs731236 (Taq1) variants
are not associated with the risk for multiple sclerosis: results of a new study and a meta-analysis. PLoS One 2013; 8: e65487.
113 Lange CM, Miki D, Ochi H, Nischalke H-D, Bojunga J, et al. Genetic analyses reveal a role for vitamin D insufficiency in HCV-associated hepatocellular carcinoma development. PLoS One 2013; 8: e64053.
114 Zhu PR, Wu QY, Yu MM, Zhang MC, Ni MX, et al. [Nucleotide polymorphism rs4880 of the SOD2 gene and the risk of male infertility]. Zhonghua Nan Ke Xue 2017; 23: 137–41. [Article in Chinese].
115 Sabouhi S, Salehi Z, Bahadori MH, Mahdavi M. Human catalase gene polymorphism (CAT C-262T) and risk of male infertility. Andrologia 2015; 47: 97–101. 116 Ji G, Gu A, Wang Y, Huang C, Hu F, et al. Genetic variants in antioxidant genes
are associated with sperm DNA damage and risk of male infertility in a Chinese population. Free Radic Biol Med 2012; 52: 775–80.
117 García Rodríguez A, de la Casa M, Johnston S, Gosálvez J, Roy R. Association of polymorphisms in genes coding for antioxidant enzymes and human male infertility.
Ann Hum Genet 2019 ;83: 63–72.
118 Salehi Z, Gholizadeh L, Vaziri H, Madani AH. Analysis of GSTM1, GSTT1, and
CYP1A1 in idiopathic male infertility. Reprod Sci Thousand Oaks Calif 2012; 19:
81–5.
119 Wu C, Man Y, Mei L, Li L, Wen X. GSTM1 null genotype contributes to increased risk of male infertility: a meta-analysis. J Assist Reprod Genet 2012; 29: 837–45.
This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-NonCommercial-ShareAlike 4.0 License, which allows others to remix, tweak, and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.