Rhizosphere Processes and Root Traits Determining the Acquisition of Soil Potassium
Texte intégral
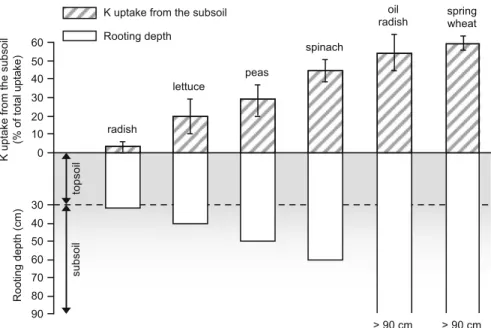
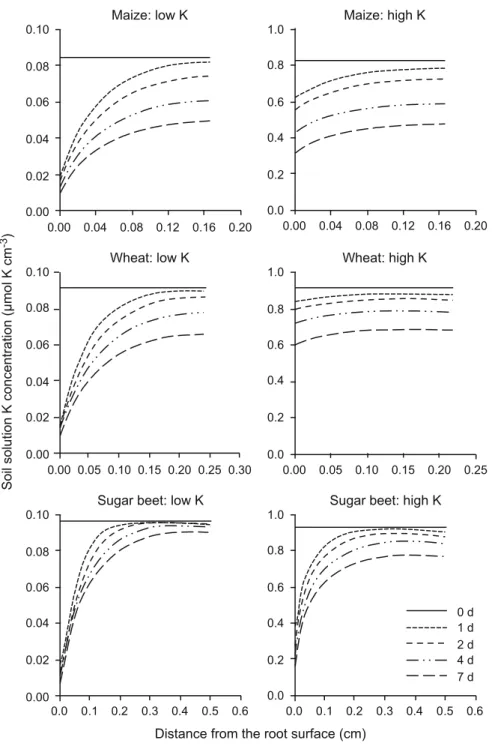
Figure

Documents relatifs
The PASTIS model was tested with experimental laboratory data that were obtained from two pesticides (Glyphosate and s-metolachlor) applied to soil columns where
L’application du géothermomètres Na-K-Ca sur l’ensemble des points à eau chaude de la zone d’étude; a donné des températures clairement supérieures à celles calculées par les
[r]
A scientific study for the systematic transfer of knowledge is essential because, first, the interdisciplinary essence of biologically inspired designs requires expertise
Temperatures given by various geothermometers (muscovite-phengite, chlorite) from stage I veins and alteration minerals range between 650 and 400°C. The main differences are: i)
These results consistently suggest that, in addition to the plant nutritional status, the initial bulk-soil pH and the specific metabolism of each plant species concomitantly drive
A study was undertaken to find out the impact of this change on the development of the root system (RS) of the trees. Individual root characteristics of each tree were noted
À la suite de cette décision de la Commission européenne, l’Agence nationale de sécurité des médicaments et des produits de santé a procédé à la levée de la décision
