Potential for acrylamide formation in potatoes: data from the 2003 harvest
Texte intégral
Figure
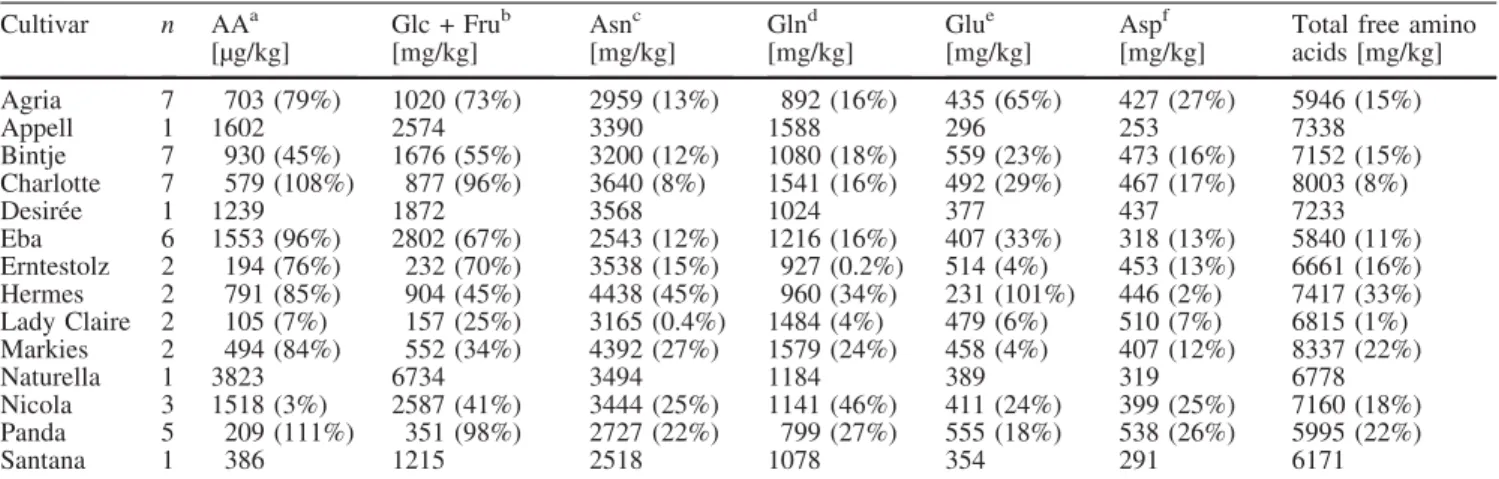
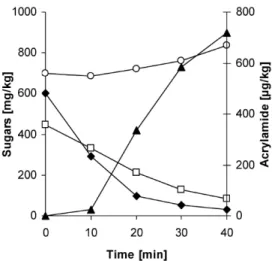
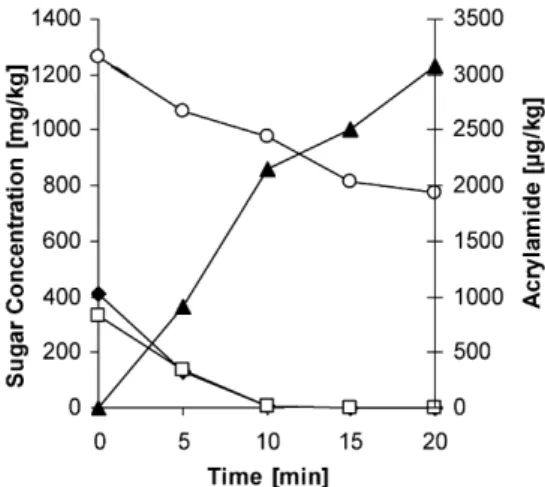
Documents relatifs
Ø The emissions abatement results from the effect of the different measures on the total mineral nitrogen applied, losses due to leaching
perform experiments on the following selected sets of similarity metrics: (i) Sim- Set1: Only name metrics; (ii) SimSet2: adding label metrics to SimSet1; (iii) SimSet3: adding
Although our development focused on synchronous co- located scenarios involving a stationary large display and an architecture with a standard wireless network and a
Anthracnose of bananas, caused by Colletotrichum musae and crown rot of bananas, caused by a broad unspecific parasitic complex; are the most important post-harvest diseases
The potential retention of an I - radioactive tracer can be calculated as a function of its pore concentration and the amount of sites available for isotopic exchange expressed as
Dans cette simulation, nous avons mis en évidence, les effets que peut provoquer la variation des résistances rotorique et statorique de la machine, dans le cadre
Similar to most of the NDACC spectrometers, the spectrum of the 0.5 ◦ wedged beam splitter shows a pronounced channeling with an amplitude of 5.7 ‰.. In contrast, the 1.27 and 2.2
Beside investigations that specifically characterize the state of the stratosphere (in particular ozone; key reservoirs and sinks such as NO 2 , NO, HNO 3 , HCl, ClONO 2 ; aerosols;
![Pour un enseignement obligatoire de la littératie numérique à l’école primaire et secondaire [Chronique - Technologies en éducation]](data:image/gif;base64,R0lGODlhAQABAIAAAP///wAAACH5BAEAAAAALAAAAAABAAEAAAICRAEAOw==)