Exceptional soft tissues preservation in a mummified frog-eating Eocene salamander
14
0
0
Texte intégral
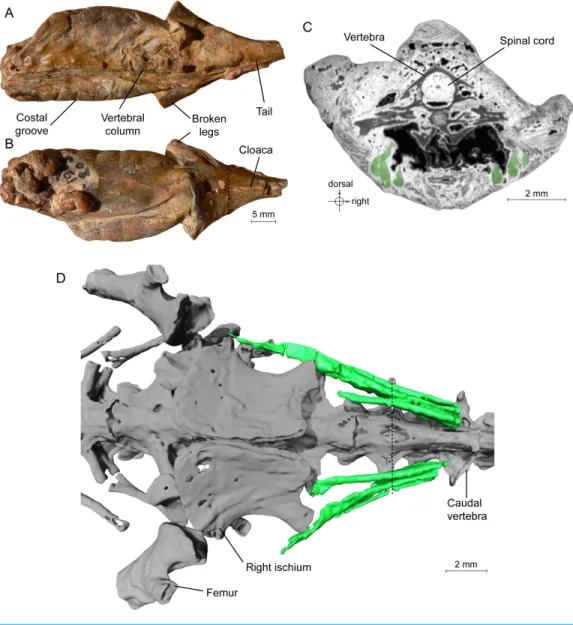
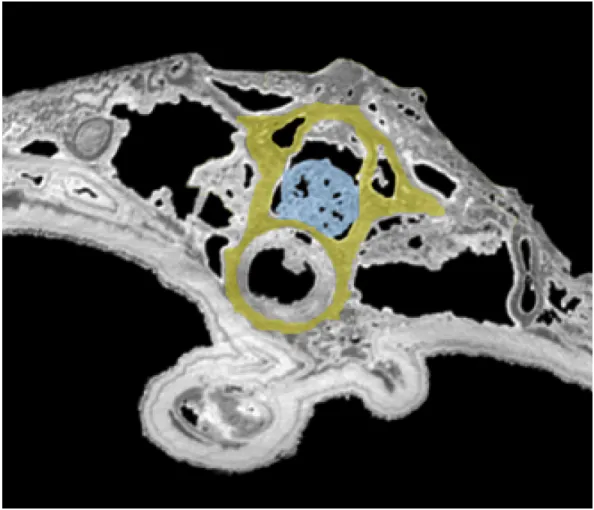
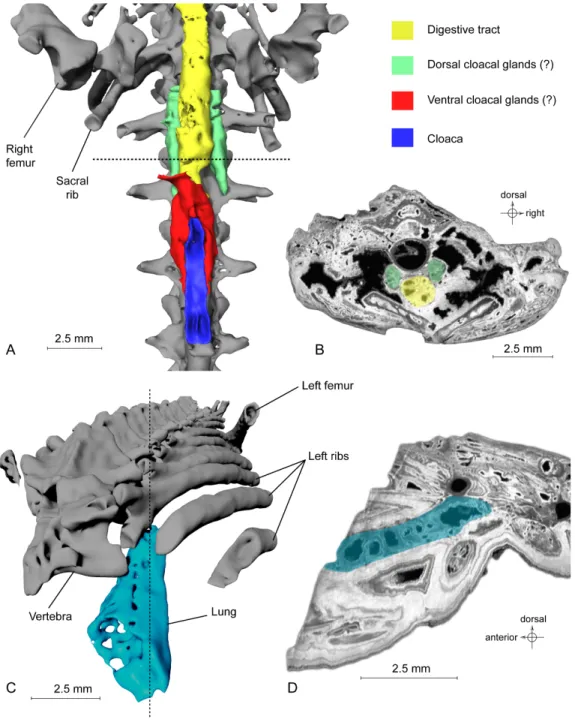
Figure

Documents relatifs