Automatic motor cortex activation for natural as compared to awkward grips of a manipulable object
11
0
0
Texte intégral
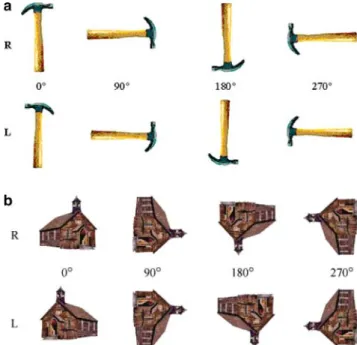
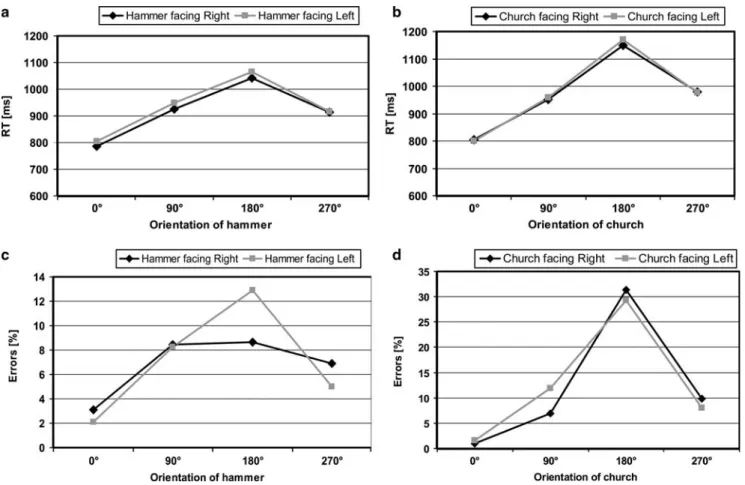
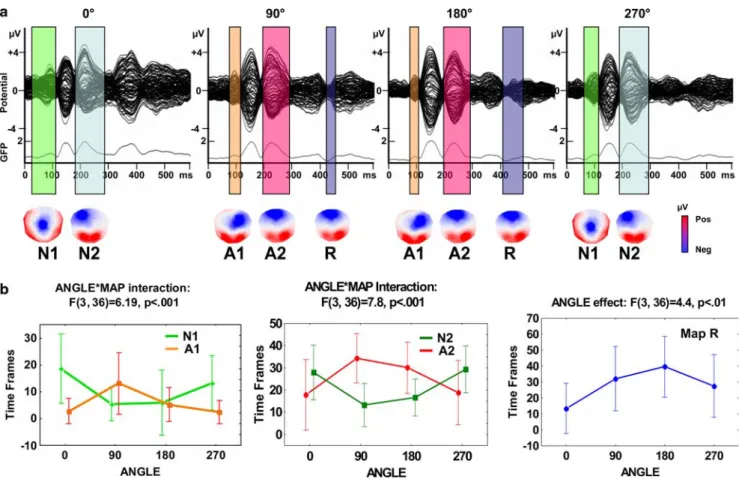
Figure

Documents relatifs