Geometry of the superior colliculus mapping and efficient oculomotor computation
15
0
0
Texte intégral
Figure

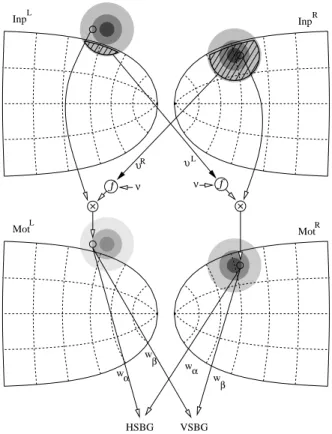

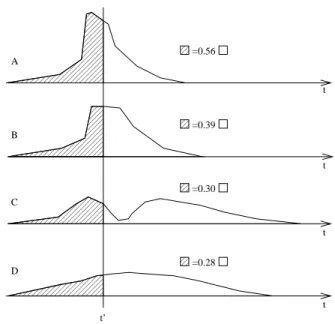
+5
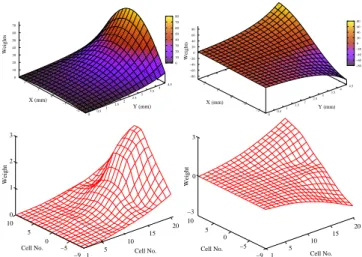



Documents relatifs