Impact of sar and agr on methicillin resistance in Staphylococcus aureus
Texte intégral
Figure
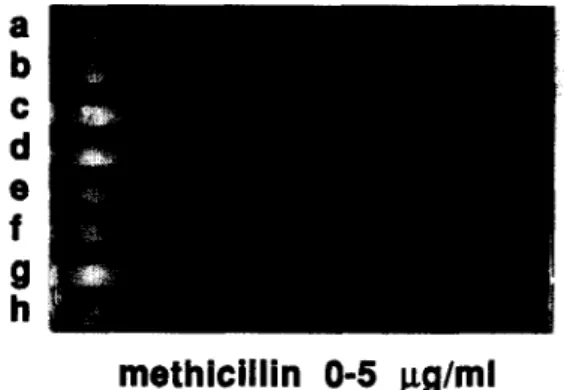
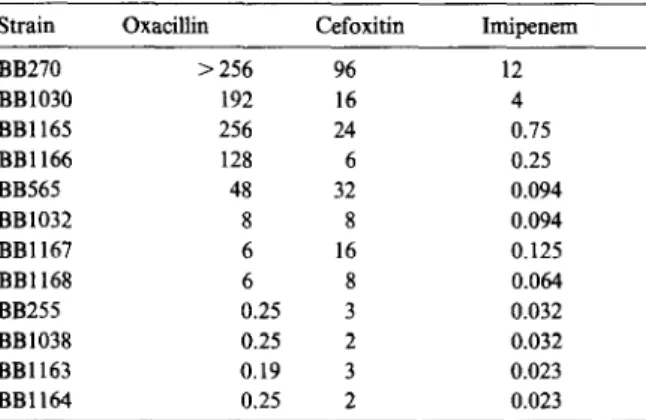
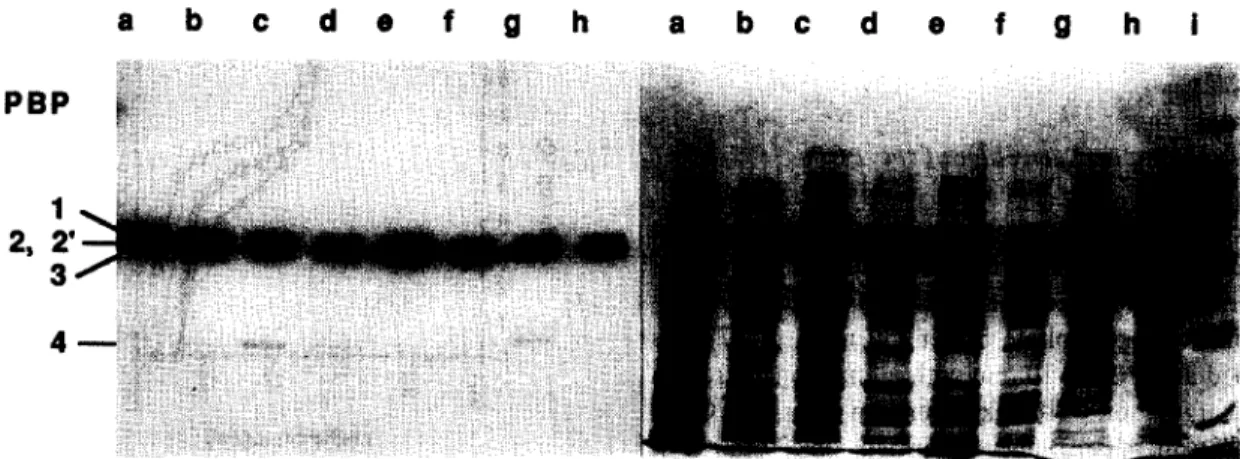
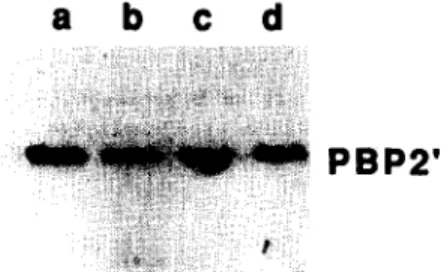
Documents relatifs
The reduction in acquired infections (AI) due to methicillin-resistant Staphylococcus aureus (MRSA) with the mupirocin/chlorhexidine (M/C) decontamination regimen has
We recently reported a high prevalence of GISA isolates in patients with persistent or recurrent MRSA bacteremia [33], in which low-level glycopeptide resistance was detected
The contribution of penicillin-binding protein 5 (PBP5) and the PBP5 synthesis repressor (Psr) to the -lactam resistance, growth, and cell autolysis of wild-type strain ATCC 9790
All sarA mutants displayed significantly reduced biofilm formation and binding to fibronectin but increased protease production in vitro, compared with their respective parental
A Small RNA Controls a Protein Regulator Involved in Antibiotic Resistance in Staphylococcus aureus.. Alex Eyraud, Pierre Tattevin, Svetlana Chabelskaya and
aureus resistance to ampicillin, penicillin G, clindamycin and tylosin showed that there was no interaction between province (Table 2) and season (Table 3); thus, these were
We recently reported a high prevalence of GISA isolates in patients with persistent or recurrent MRSA bacteremia [ 33 ], in which low-level glycopeptide resistance was detected
