HAL Id: tel-01402657
https://tel.archives-ouvertes.fr/tel-01402657
Submitted on 25 Nov 2016HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Genetic architecture of seed yield and its components in
winter oilseed rape (Brassica napus L.) grown under
limiting nitrogen condition
Anne-Sophie Bouchet
To cite this version:
Anne-Sophie Bouchet. Genetic architecture of seed yield and its components in winter oilseed rape (Brassica napus L.) grown under limiting nitrogen condition. Plant breeding. Université Rennes 1, 2015. English. �NNT : 2015REN1S150�. �tel-01402657�
ANNÉE 2015
THÈSE / UNIVERSITÉ DE RENNES 1
sous le sceau de l’Université Européenne de Bretagne
pour le grade de
DOCTEUR DE L’UNIVERSITÉ DE RENNES 1
Mention : Biologie
Ecole doctorale Vie - Agro - Santé
présentée par
Anne-Sophie Bouchet
Préparée à l’Unité Mixte de Recherche 1349 IGEPP
Institut de Génétique Environnement et Protection des Plantes
UFR Sciences de la Vie et de l’Environnement
Architecture
génétique du
rendement et de ses
composantes chez le
colza d’hiver
(Brassica napus L.)
cultivé sous
contrainte azotée
Thèse soutenue à Rennes
le 24 novembre 2015
devant le jury composé de :Jacques LEGOUIS
Directeur de recherche INRA / rapporteur
Rod SNOWDON
Professor and Chair of Plant Breeding, Justus Liebig University, (Germany) / rapporteur
Anne-Marie CORTESERO
Professeur Université Rennes 1 / examinateur
Christian HERMANS
Chercheur, Université Libre de Bruxelles (Belgique) / examinateur
Sébastien PRAUD
Chef de projet Biogemma / examinateur
Michel RENARD
Directeur de recherche INRA / directeur de thèse
Anne LAPERCHE
Maître de conférenceAGROCAMPUS OUEST/
co-directrice de thèse
Nathalie NESI
ANNEXE 2 (Modèle dernière page de thèse)
VU : VU :
Le Directeur de Thèse Le Responsable de l'École Doctorale (Nom et Prénom)
VU pour autorisation de soutenance
Rennes, le
Le Président de l'Université de Rennes 1
Guy CATHELINEAU
VU après soutenance pour autorisation de publication :
Le Président de Jury,
TABLE OF CONTENTS
Glossary GENERAL CONTEXT – The challenges for sustainable rapeseed production ... 1 CHAPTER I – Introduction, Part A: Genetic diversity for nitrogen use efficiency in rapeseed ... 5 I.A‐ 1. Why improving nitrogen use efficiency (NUE) in rapeseed? ... 7 I.A‐ 2. What is the genetic diversity for NUE‐associated traits in rapeseed? ... 10 I.A‐ 3. Breeding for N‐efficient rapeseed cultivars with optimized oil yield: where do we stand? . 16 I.A‐ 4. Future prospects for NUE improvement in rapeseed ... 19 CHAPTER I – Introduction, Part B: Genetics and genomics of Brassica napus... 29I.B‐ 1. Extensive genomic duplications and rearrangements have modeled the current B. napus genome ... 30 I.B‐ 2. The B. napus genetic resources ... 31 I.B‐ 3. Evaluation of the Linkage Disequilibrium (LD) extend in the rapeseed genome ... 31 I.B‐ 4. Genotyping resources and genetic maps in B. napus ... 32 I.B‐ 5. Genomic data available for the Brassicaceae ... 33 I.B‐ 6. Current knowledge on the genetic control of seed yield related traits in rapeseed in relation with its genome structure ... 33 CHAPTER I – Introduction, Part C: Objectives of the thesis ... 35 CHAPTER II.A – Preliminary results ... 37 II.A‐ 1. Selection of two doubled haploid populations for linkage analyses ... 37 II.A‐ 2. Characterization of the population used for linkage disequilibrium analyses ... 37 II.A‐ 3. Selection of the model for linkage disequilibrium analyses ... 38 CHAPTER II.B – Article 1 ... 39 CHAPTER II.C – Article 2 ... 55 CHAPTER II.D – Complementary results: Characterization of the plant N constraint ... 85 CHAPTER III – Genetic control of oil content in Brassica napus ... 86 III.1. Introduction ... 87 III.2. Material and methods ... 88 III.3. Results ... 90 III.4. Discussion ... 93 CHAPTER IV – Discussion and prospects ... 99 IV.1. Main highlights of the thesis ... 99 IV.2. Controlling the environmental effects to evaluate the G × N interactions ... 100 IV.2.1. Strong G × E effects were involved in the control of seed yield related traits ... 100
IV.2.2. Optimization of trial designs for the evaluation of high number of genotypes under N constraints ... 101 IV.2.3. Crop models for the investigation of the G × E interactions ... 102 IV.3. New traits to decipher NUE components in large sets of rapeseed genotypes ... 103 IV.3.1. NUE traits at the canopy level ... 103 IV.3.2. NUE traits at the plant level ... 104 IV.3.3. NUE traits at the molecular level ... 105 IV.4. New challenges raised by this thesis ... 106
IV.4.1. Towards an evaluation of rapeseed genotypes grown under multiple and combined stress conditions ... 106 IV.4.2. Impact of polyploidy on the response to low N availability ... 107 IV.4.3. From QTLs to plant breeding ... 107 REFERENCES ANNEXES SCIENTIFIC ACTIVITIES
Glossary
ACK: Ancestral Crucifer Karyotype AM: Aviso × Montego DH population ANOVA: ANalysis Of Variance BC: Back Cross
BHCA: Brassica Hexaploid Common Ancestor Ch: Châteauroux
DH: Doubled Haploid Dij: Dijon
DK: Tenor × Express DH population DY: Darmor-bzh × Yudal DH population DTF: Days To Flowering
GDD: Growing Degree Day GFP: Grain Filling Period GLM: General Linear Model GS: Genomic Selection GSL: Glucosinolates h²: heritability values HI: Harvest Index IBS: Identity by State
K matrix: Identity by State Kinship matrix LA: Linkage Analysis
LAI: Leaf Area Index LD: Linkage Disequilibrium
LDA: Linkage Disequilibrium Analysis LF: Least Fractionated
LG: Linkage Group LR: Le Rheu
MAF: Minor Allele frequency Md: Mondonville
MF: Most Fractionated
MIM: Multiple Interval Mapping MLM: Mixed Linear Model N: Nitrogen
NAE: Nitrogen Assimilation Efficiency NAM: Nested Association Mapping NHI: Nitrogen Harvest Index
NNI: Nitrogen Nutrition Index
NRE: Nitrogen Remobilization Efficiency NUE: Nitrogen Use Efficiency
NUpE: Nitrogen Uptake Efficiency NUtE: Nitrogen Utilization Efficiency O: seed Oil content
OSR: OilSeed Rape OY: Oil Yield PAI: Pod Area Index
PCA: Principal Components Analysis Pr: seed Protein content
Pre: Prémesques PrY: Protein Yield
Q matrix: Matrix of the structure of the population QTL: Quantitative Trait Locus
RIE: Radiation Interception Efficiency RIL: Recombinant Inbred Line RUE: Radiation Use Efficiency Sel: Selommes
SN: Seed Number per m²
SNP: Single Nucleotide Polymorphism SPAD: Soil Plant Analysis Development SSR: Simple Sequence Repeat
SY: Seed Yield
TN: Tapidor × Ningyou DH population TSW: Thousand Seed Weight
Ver: Verpillères
WGD: Whole Genome Duplication WGT: Whole Genome Triplication WOSR: Winter OilSeed Rape
[00]: low erucic acid/low GSL content varieties [0+]: low erucic acid/high GSL content varieties [+0]: high erucic acid/low GSL content varieties [++]: high erucic acid/high GSL content varieties
GENERAL CONTEXT – The challenges for sustainable rapeseed production
1
GENERAL CONTEXT –
The challenges for sustainable rapeseed production
•
General context for worldwide seed production and position of oilseed crops
The worldwide demand in vegetable oils and proteins is being experiencing an important increase for the last decades, due to growing population and higher living standards. In parallel, agriculture production is facing the changes induced by global warming and must adapt to new climate parameters and reduce its footprint on the environment. Therefore, high seed yield and high seed quality are major goals for crop production, while at the same time there is a need to stabilize seed production under fluctuating environments (various biotic and abiotic stresses) and to reduce the environmental impacts of agriculture by lowering the inputs (fertilizers, pesticides, herbicides). The seed quality criteria vary depending on its uses: agronomical, food or industrial applications. The physiological quality is assessed on criteria such as the sanitary status, the germination performance and the ability to generate vigorous seedlings that are crucial prerequisites for crop establishment and high yield levels in crop production. For food and industrial purposes, the quality refers to the content and composition of storage products as well as their extraction yield.
The main oleaginous crops such as rapeseed, soybean and sunflower can provide solutions to increase the supply of plant oils and proteins in the future. However, these cultures are not equivalent in terms of quantity and quality. Indeed, rapeseed has an average grain yield higher than soybean and sunflower (∼ 3.5, 2.7, 2.5 t / ha respectively, in Western Europe) and higher oil yield (∼ 1.5, 0.5, 1.1 t / ha resp.), but a lower yield of proteins to that of soybean (∼ 0.6, 0.95 t / ha resp.). In addition, the quality of rapeseed meals is substantially lower than that of soybean meals in terms of protein content, amino acid balance and in vivo digestibility, which limits its use for livestock feeding and thus maintains Western Europe largely dependent on soybean meal for protein supply.
The substantial investments made by the breeders and research organizations since the 1950’s demonstrate that the improvement of seed yield and quality is a major strategic and economic issue. However, the high number of genes involved, the complexity of regulatory mechanisms and the impact of environmental factors make the understanding of the genetic and molecular determinism of the seed yield and quality complex despite extensive work on the seed so far.
1 2 3 4 1955 1960 1965 1970 1975 1980 1985 1990 1995 2000 0 old varieties Yield improvement of the (0+) varieties First double low varieties (00) Yield improvement of the (00) varieties Hybrid varieties
Seed yield in t/ha
First simple low varieties
(0+)
Figure 3. Yield evolution in rapeseed crop from 1950 (France)
00 (double low) refer to varieties with low contents in erucic acid (C22:1) and glucosinolates; 0+ (simple low) refer to varieties with low C22:1 content only.
Adapted from GNIS, Terres Inovia, CTPS.
Figure 1. Worldwide rapeseed production (Mt seeds) and main producing countries in 2013 From FAOSTAT, 2013. 0 100 200 300 400 500 600 700 800 1961 1971 1981 1991 2001 2011 Main producers in 2013 Seeds (in Mt) 35% 25% 20% 11% 9% Europe Canada China India Other 70 60 50 40 30 20 10 0 1961 1971 1981 1991 2001 2011
Figure 2. Rapeseed production areas in France (ha) in 2012 and location of main plants for seed treatment (crushing, oil refinery, esterification)
Charts reproduced from SCEES and ONIOL (left) and FNCG (right).
Area (in ha)
crushing refinery esterification
GENERAL CONTEXT – The challenges for sustainable rapeseed production
2
•
Specific issues related to rapeseed
Oilseed rape (Brassica napus L.) is an allotetraploid (genome AACC, 2n = 4x = 38) belonging to the Brassicaceae family. It arose from the spontaneous interspecific hybridization between turnip (B. rapa, AA, 2n = 2x = 20) and cabbage (B. oleracea, CC, 2n = 2x = 18) (U 1935). The B. napus genome is only ten times larger than that of the closely related Brassicaceae Arabidopsis thaliana (~1.1 Gbp vs. ~125 Mbp) but shows a highly complex segmental structure in comparison to the plant model. Indeed, due to several whole genome duplication events, genome fragmentation and homeologous non-reciprocal translocations between A and C sub-genomes that altogether occurred during the plant speciation, the Brassica napus genome is a patchwork with considerable variations in gene copy number and syntenic conservation (reviewed in Edwards et al. 2013).
The genetic diversity of rapeseed is highly structured into winter and spring types (Bus et al. 2011). Winter types are mostly grown in Western Europe where winters are mild, while, spring types are sown primarily in northern latitudes and Australia. The seed yields are usually higher for winter compared to spring types. In addition, the creation of the low erucic acid/low glucosinolates varieties (double low quality, “00”) is the cause of a genetic diversity bottleneck as only a few genetic sources were used for each conversion.
Rapeseed is the most important oilseed crop in France and Europe, and the third one worldwide with an annual production of ∼75 Mt of seeds (Figure 1). The main producers are Europe (25.6 Mt), Canada (18 Mt), China (14.5 Mt) and India (7.8 Mt) (Food and Agriculture Organization of the United Nations 2013). In France, most of the rapeseed production (∼ 5.2 Mt seeds) is ensured by winter varieties grown in the center and north-eastern areas (∼ 1.5 ha) (Figure 2).
The crop is primarily used for food and feed and has recently gained increasing interest as a source for bio-products. To date, rapeseed oil accounts for around 15% of the worldwide plant oil production. The major constituents of B. napus seeds are water, oil (45-50% w/w), protein (20-25% w/w) and residues which mainly include carbohydrates, fibers, minerals and ashes. The seed cake resulting from the extraction of oil contains more than 35 % protein. Seed oil content has remained almost stable over the past two decades; nonetheless, an increase in a few percent would lead to a major improvement of oil yield due to the crop worldwide potential. At the same time, there is a need to consider improvement in other seed components, with a special emphasis on seed protein content and composition in rapeseed for better valorization of the seed meal. Targets for seed quality improvement in rapeseed have been reviewed (Nesi et al. 2008; Abbadi and Leckband 2011). Average seed yield of rapeseed crop has shown an increase by up to 50% over the last six decades (Figure 3). It depends on variety type (inbred line, F1 hybrid or varietal association), environmental
Figure 4. Which optimal N input for optimizing OSR yield?
Nitrogen (N) is at the center of major processes including biomass production and yield elaboration. Rapeseed is a high N demanding crop with a low N use efficiency (NUE). As a matter of fact, N fertilization pratice as well as varieties need to be optimized to ensure the crop sustainability at the agronomic, environmental and economic levels.
GENERAL CONTEXT – The challenges for sustainable rapeseed production
3 and therefore vary from one country to the other, ranging from around 1.5–2 t/ha in Canada and Eastern Europe (extensive production of spring varieties) to 3.5 t/ha in Western Europe (intensive production of winter-type rapeseed). But grain yield is a very complex trait in rapeseed (Diepenbrock 2000). The complexity is mainly related to the sequential appearance of flowers and then of pods. Therefore, at a given date, flowers, young pods and mature pods can coexist on the same plant. As a consequence, the crop is able to compensate a yield component limitation by another one (e.g., development of extra ramifications when plant density decreases) and a given yield can result from different combinations of yield components. Understanding the control of yield elaboration is thus very challenging.
Finally, rapeseed crop is highly demanding on inputs compared to other crops, with particular high requirements in mineral N inputs (∼ 160-250 kg N/ha; Rathke et al. 2005). N fertilization is the main expense for farmers and represents 2/3 of the energetic cost at the production level. In addition, there is serious concern regarding N loss in the field, giving rise to soil and water pollution by nitrate leaching, as well as air pollution by greenhouse gas (GHG) emissions. Rapeseed is often reported as a poor N-efficient crop with a low ratio of seeds produced per N unit applied (∼15kg seed/kg plant available N; Figure 4), around half that for cereals (Sylvester-Bradley and Kindred 2009). Therefore, rapeseed is showing a low capacity in using the available N to produce seeds that defines the nitrogen use efficiency (NUE, Moll et al. 1982). Total NUE is split into two components: i) the ability of the plant to capture the N from the soil (that defines the nitrogen uptake efficiency, NUpE), and ii) the ability to use the N absorbed to produce seeds (nitrogen utilization efficiency, NUtE).
In conclusion, increasing abiotic stress tolerance, nutrient use efficiency, seed quality and seed yield in the face of changing environments and emerging production constraints are current challenging goals for sustainable rapeseed production. Agricultural practices may be optimized to provide new solutions in terms of fertilization management. Another option is the development of varieties with an improved NUE especially under low N inputs that can remove the nutrient applied to soil efficiently (higher NUpE), and further assimilate, recycle and store nutrients to produce high quantities of high quality seeds (higher NUtE). However, considering the intricate relationships between plant growth, nutrient dynamics (especially N) and seed yield, new approaches are required to analyze and dissect complex traits and associate their components to the underlying genetic mechanisms.
GENERAL CONTEXT – The challenges for sustainable rapeseed production
4
•
Scientific context for my thesis
My PhD work aims at understanding the organization of the genetic control of seed yield related traits in rapeseed under limiting N availability.
My PhD was funded by the French ministry of higher education and research and started on October, 1st 2012 within the doctoral school Vie-Agro-Santé of the University of Rennes 1. This thesis was conducted within the French National agronomy Institute (INRA) at the IGEPP unit (Institute of Genetics, Environment and Plant Protection) in the team entitled “Yield and Abiotic Challenges” (http://www6.rennes.inra.fr/igepp_eng/RESEARCH-TEAMS/Yield-under-Abiotic-Challenges) under the co-supervision of Anne LAPERCHE and Nathalie NESI (PhD supervisor: Michel RENARD). In addition, the data of my PhD were acquired within the framework of two national collaborative projects entitled GENERGY (ANR-07-GPLA-016) funded by the French National Research Agency (ANR) and RAPSODYN (ANR-11-BTBR-0004) funded by the program “Investments for the Future” (http://www.rapsodyn.fr/en/).
The present manuscript contains four chapters. Chapter I reviews the current knowledge about N dynamics and rapeseed genetic variability for NUE-related traits on the one hand (part A) and gives a general presentation on the genetics and genomics features of B. napus on the other hand (part B). The part C raises the scientific questions of the thesis. In the chapter II, the results of the genetic analyses of seed yield under N limitation are presented in two articles. Chapter III provides a focus on the organization of genomic regions controlling the oil content and their putative underlying functions. Finally, the manuscript ends up with a discussion on the results acquired and the prospects for possible improvements of yield and NUE in rapeseed (chapter IV).
CHAPTER I
Introduction
5
CHAPTER I – Introduction, Part A
Genetic diversity for nitrogen use efficiency in
rapeseed
This first introductive chapter aims to address the principal issues of mineral nitrogen (N) nutrition that interplay on the environmental and economic balance of rapeseed and to pinpoint the key levers to improve the sustainability of this crop towards its N dependency. The nitrogen use efficiency (NUE) will be first documented during the rapeseed plant cycle, along with the main mechanisms that are associated. The genetic diversity available in rapeseed for the corresponding traits and the current knowledge about their genetic control will be reviewed. Finally, a last part will be dedicated to the future prospect to improve the NUE in rapeseed with a focus on the new genetic structures, modeling and phenotyping tools that are under development for this crop.
This chapter has been written in collaboration with Andreas Stahl and Rod Snowdon from the Plant Breeding Department of the Justus Liebig University (Giessen, Germany) and will be soon submitted as a review paper.
6
What is the genetic diversity pool to improve the nitrogen use
efficiency in rapeseed?
Anne-Sophie Bouchet1*, Andreas Stahl2*, Christine Bissuel-Belaygue3, Anne Laperche3, Nathalie Nesi1 and Rod Snowdon2
1
INRA, UMR 1349 Institute for Genetics, Environment and Plant Protection, Domaine de la Motte, 35650 le Rheu, France
2
Department of Plant Breeding, IFZ Research Center for Biosystems, Land Use and Nutrition, Justus Liebig University, Heinrich-Buff-Rizg 26-32, 35392 Giessen, Germany
3
AGROCAMPUS OUEST, UMR 1349 Institute for Genetics, Environment and Plant Protection, Domaine de la Motte, 35650 le Rheu, France
*
These authors contributed equally to the manuscript
Abstract
Mineral nitrogen fertilizing has increased yield in major crops for the last century, but is also the cause of massive water, soil and air pollution through nitrogen leaching and greenhouse gas emission. Reduction of nitrogen inputs while maintaining high yields is therefore essential to ensure a more sustainable agriculture in the future. A key aspect for yield sustainability is the improvement of the nitrogen use efficiency (NUE) through the enrichment of gene pools. Identifying the key traits and the corresponding phenotyping methods, as well as evaluating the genetic diversity for these traits are prerequisites to resolve this challenge. Rapeseed (Brassica napus L.), one of the major oil crops worldwide, remains highly dependent on nitrogen fertilization due to a low NUE. While nitrogen metabolism in general has been investigated for a long time, more information is needed on the genetic diversity for the NUE in rapeseed. In this article we review aspects of NUE improvement in B. napus, with a special emphasis on winter oilseed rape. A focus on the phenotypic traits associated with the different NUE components, including N uptake, assimilation and remobilization, will be presented, as long as the genetic diversity available for those traits and the potential breeding strategies. This information will summarize the knowledge about N dynamics throughout the rapeseed crop cycle and outline crucial elements that should be considered in research and breeding in order to improve the competitiveness of this crop at the agronomic, environmental and economic levels.
Keywords
Brassica napus, seed yield, nitrogen uptake efficiency (NUpE), nitrogen utilization efficiency (NUtE), nitrogen remobilization, nitrogen losses, genetic diversity, genomic selection
7
I.A- 1.
Why improving nitrogen use efficiency (NUE) in rapeseed?
Rapeseed has a low NUE
Rapeseed (Brassica napus L.) is a crop of major economic importance mainly grown for its oil-rich seeds used for human consumption as well as industrial purposes, and for the proteins of the seed cake used in animal feeding. It is the third oil crop in the world, after soybean and palm oil (Food and Agriculture Organization of the United Nations 2013). However, rapeseed is highly demanding in inputs compared to other crops, particularly for mineral nitrogen (N) inputs (Rathke et al. 2005). In addition, rapeseed is often reported as a poor N-efficient crop with a low ratio of seeds produced per N unit applied, around half that for cereals (Sylvester-Bradley and Kindred 2009). Therefore, rapeseed is showing a low capacity in using the available N to produce seeds that defines the nitrogen use efficiency (NUE, Moll et al. 1982). Total NUE is split into two components that can be evaluated at the canopy or plant levels: i) the ability of the plant to capture the N from the soil (that defines the nitrogen uptake efficiency, NUpE), and ii) the ability to use the N absorbed to produce seeds (nitrogen utilization efficiency, NUtE) (Moll et al. 1982). When the plant level is considered, NUtE includes the capacity of the plant to assimilate N from the soil (nitrogen assimilation efficiency, NAE) and then to remobilize it into the seeds (nitrogen remobilization efficiency, NRE) (Masclaux-Daubresse et al. 2010).
How to estimate the NUE?
Although the definition of the NUE is quite well established, its estimation is a more complex issue since this process can be approached at different levels as previously reviewed by Good et al. (2004) and Rathke et al. (2006). From an agronomic point of view, NUE may be considered as the direct increase in seed yield per additional unit of N fertilizer or the ability to convert N to seed yield by dividing the difference of seed yield between two N nutrition regimes with the N supplied (that defines the agronomic efficiency) (Good et al. 2004; Rathke et al. 2006). From a physiological point of view, NUE can be considered as the nitrogen/carbon balance in the shoots at harvest by measuring the relationship between the biomass and the N content of the shoots (Good et al. 2004). Finally, when the plant scale is considered, the N flux measurements by using isotope labelling techniques (15N) allow assessing precise N dynamics throughout the plant organs.
Pre-flowering NUpE
Pre-flowering NAE and NRE
Post-flowering NUpE
Post-flowering NAE and NRE Process
Vegetative growth Vernalization Bolting Flowering Seed filling Seed quality Yield potential elaboration
- Nb of leaves
- Nb of ramifications / plant - Nb of pods / ramification
- Nb of seeds / pod
=> Seed nb / m²
Yield potential expression
- Seed abortions => seed nb/m² - Seed filling => TSW
Seed yield
Foliar primordia initiation
Floral initiation
N remobilization
N loss through green house gas emissions
1 3 4 2 LAI PAI N loss through pod shattering N mineralization from fallen
leaves: leaching and N uptake N loss through fallen leaves
and leaching N losses N absorption N a v a ila b le i n t h e s o il N fertilization N fertilization Soil N balance N mineralization
NUE
Figure I.A- 1. N dynamics over the crop cycle in rapeseedThe figure depicts the interconnected relationships between plant growth, N dynamics and yield elaboration over the crop cycle. The N uptake is represented by blue arrows of which width indicates the relative amount of N absorbed at a given time point. The N losses are shown with black arrows of which width indicates the relative amount of N lost at a given time point. The critical stages for final NUE establishment are noted as following: 1) pre-flowering NUpE and sequential NRE, 2) post-flowering NUpE, 3) sequential and monocarpic NRE during the post-flowering and the seed filling periods, and 4) the interactions between NUE and LAI and PAI.
8 Plant growth, N absorption and yield elaboration make a complex story in winter rapeseed
Seed yield is a complex trait resulting from the combination of several components such as the plant density, number of ramifications per plant, pod number per ramification, seed number per pod, single seed weight (Diepenbrock 2000). Sequential determination of the yield components allows splitting the crop cycle into two overlapping phases, with the establishment of the yield potential on the one hand and the expression of the yield potential on the other hand (Figure I.A- 1).
During the vegetative growth over autumn and winter periods, the dynamics of leaf area index (LAI) depends mainly on climatic conditions through radiation interception and radiation use efficiency (RIE and RUE). The N soil availability also conditions the LAI and depends on the pedo-climatic conditions: while in temperate climates, the soil N mineralization may be enough to cover for the plant N needs during autumn, N fertilization may be needed in colder areas. The root system quickly develops after seedling emergence and mineral N is efficiently absorbed from the soil and stored into the vegetative biomass. For instance, rapeseed shows a high NUpE at early stages with up to 100 kg N/ha absorbed making it a valuable catch crop during the fall (Rossato et al. 2001). As a matter of fact, plant biomass increases dramatically over the autumn. Simultaneously, N is remobilized from older to younger organs (sequential N remobilization), which promotes the initiation of foliar primordia as soon as the end of autumn. As a consequence, the numbers of ramifications and photosynthetic leaves as well as flowers and ovules are partly determined before the end of winter (Figure I.A- 1) and will prefigure the final seed number / m² that is highly correlated with the seed yield. Therefore, an N stress during that period might negatively impact the final yield. During winter, a part of the leaf area produced during this first period could be destroyed by frozen thus leading to important N losses through the fallen leaves (2-3.5% of leaf dry weight, Malagoli et al. 2005a).
A new LAI is generated during the spring period from the beginning of stem elongation to flowering. Therefore, soil N availability during the spring development greatly affects LAI expansion and duration. During that period, N is actively remobilized from senescing leaves to younger leaves and stems (sequential senescence) and then to pods and seeds (monocarpic senescence) (Figure I.A- 1). From the flowering stage onwards, leaves are no longer produced and their senescence increases quickly, with a different remobilization ability depending on their position on the main stem (Malagoli et al. 2005a). In parallel, photosynthesis is partly ensured by the pod area index (PAI). In addition, the N pool is also maintained through the N re-absorption from the leaves fallen during autumn, with an uptake efficiency that can reach 40% of the N content from the fallen leaves (Dejoux et al. 2000).
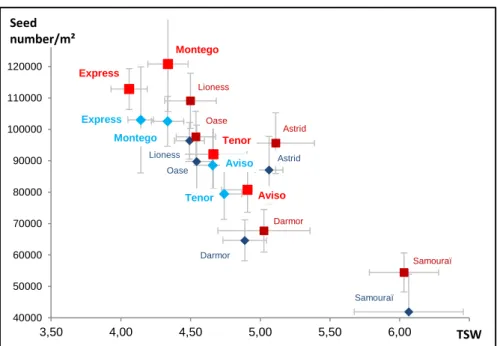
Figure I.A- 2. Seed yields of 92 winter oilseed rape genotypes grown under two contrasting N nutrition regimes (N- vs. N+) in Western France
Data were acquired from one trial conducted under field conditions in Le Rheu (France) in the growing season 2009-2010 as described elsewhere (Bouchet et al. submitted). Rectangles highlight the high yielding and/or N efficient genotypes (in red and blue respectively).
Seed yield in N- condition
S e e d y ie ld in N + c o n d it io n
High yield genotypes
N -e ff ic ie n t g e n o ty p e s
9 However, the ratio of the plant N content to the N supplied usually does not exceed 50% to 60% in rapeseed (Malagoli et al. 2005a and references therein). As a consequence, a major problem is the large N surplus which represents a significant shortfall for the farmers and has negative environmental impacts. To improve its economic and environmental competitiveness, breeding for N-efficient rapeseed varieties is a main target. Based on the N dynamics and the plant development throughout the crop cycle, a number of critical processes have to be investigated as indicated in Figure I.A- 1, including i) the pre-flowering NUpE and NRE to insure the establishment of the yield potential, ii) the post-flowering NUpE to maintain a N pool in the plant during the seed filling stage, iii) the sequential and monocarpic N remobilization from the flowering onwards to develop the crop yield potential, and iv) the interactions between the nitrogen and carbon balance involving photosynthetic processes such as LAI, PAI, and radiation use efficiency (RUE).
Is there a scope to improve NUE in rapeseed?
Brassica napus L. (2n=4x=38, AACC) results from the recent spontaneous interspecific cross between Brassica rapa (2n=2x=20, AA) and Brassica oleracea (2n=2x=18, CC) (U 1935). The short history of domestication of this species (400 to 500 years) and the concentration of breeding efforts to improve the quality of the oil in the 1970’s partly explains the narrow genetic diversity in rapeseed (Hasan et al. 2005). In addition, rapeseed is highly structured into spring versus winter types (Bus et al. 2011). Inbred lines have dominated the rapeseed production over the last decade and genetic diversity for NUE was addressed for that genetic type. For instance, when a diversity set of inbred lines was grown under contrasting N nutrition levels, it was possible to identify three kinds of genotypic responses, with i) low potential genotypes with low yield in both N conditions, ii) genotypes with high potential in N+ condition but a low response to N limitation, and iii) high N efficient genotypes showing high yield in both N conditions (Figure I.A- 2). The conversion into F1 hybrid lines since the
last decade has provided new genetic variations that could be exploited for NUE improvement. For instance, Kessel and coworkers (2012) compared the seed yield response to a change of N condition in modern cultivars, hybrids, resynthesized lines and old cultivars and found the hybrids to be the most productive genotypes whatever the N nutrition conditions. Several studies already explored the genetic diversity relative to N related traits under field or controlled conditions, in spring (Yau and Thurling 1986; Svečnjak and Rengel 2005, 2006; Balint et al. 2008a) and winter cultivars (Schulte auf'm Erley et al. 2007; Berry et al. 2010; Schulte auf'm Erley et al. 2011; Kessel et al. 2012; Ulas et al. 2013; Lee et al. 2015). Combining genetic variations from each type by backcrosses for example could enrich the genetic pool for NUE improvement. Moreover, introgressing genome portions from
N1 N2 1.5 2.0 2.5 3.0 3.5 10 15 20 25 10 15 20 25 NUE N c o n te n t in t h e a b o v e -g ro u n d b io m a s s a t fl o w e ri n g ( in % )
N content in the leaves at flowering and NUE correlation
r² ~ 0.42 *** r² ~ 0.41 *** N1 N2 0.5 1.0 1.5 2.0 10 15 20 25 10 15 20 25 NUE N c o n te n t in t h e a b o v e -g ro u n d b io m a s s a t fl o w e ri n g ( in % )
N content in the above-ground biomass at flowering and NUE correlation
r² ~ 0.35 ***
r² ~ 0.49 ***
a N- N+
N- N+
b
Figure I.A- 3. Correlations between NUE and N contents in the shoot (a) or in the leaves (b) of rapeseed genotypes at the flowering stage
Data were acquired from 30 winter rapeseed genotypes grown under semi-controlled conditions with two contrasted N nutrition regimes (N- vs. N+) as described by Stahl et al. (in prep).
10 related species such as B. rapa or B. oleracea, as it was performed by Seyis et al. (2003) or using resynthesized new-type B. napus like Wang et al. (2014) could also be a key for exploring new variations for NUE breeding.
In summary, potential genetic diversity exists for NUE breeding in rapeseed. Moreover, NUE in a broader sense has been studied for many decades in several crop species and is well documented (Fageria and Baligar 2005; Hirel et al. 2007; Garnett et al. 2009; Xu et al. 2012; Vincourt 2014). However, data are rather scarce concerning the key points of the NUE elaboration and the associated genetic variations in rapeseed.
I.A- 2.
What is the genetic diversity for NUE-associated traits in rapeseed?
Seed yield is a good indicator of the NUE in rapeseed
Improving seed yield under low N nutrition level is a first step towards the improvement of the NUE (Good et al. 2004). Therefore, the estimation of the seed yield (SY) under different N regimes is an indicator of the global NUE (Figure I.A- 2). In addition, the harvest index (HI, defined as the ratio of seed weight to total plant dry matter) and the nitrogen harvest index (NHI, defined as the ratio of seed N amount to total plant N amount at harvest) provide two estimators of the rapeseed capacity to mobilize the N and C assimilates to produce seeds. However, the main drawbacks of these indicators rely in the fact they are complex variables that are acquired at the end of the crop cycle, thus integrating additional processes linked to global life traits such as plant growth or resistance to stresses that all impact the estimation of the global NUE. In addition, breeding for seed yield or harvest index remains difficult because of their low heritability (h²), especially under low N environments. A possible strategy proposed by Thurling (1991) is therefore to decipher complex traits in simpler variables that can be acquired all along the crop cycle.
NUpE has to be improved to maintain a high N pool in the plant until the flowering stage
Several studies reported that the NUE was more correlated to the NUpE than to the NUtE under limiting N fertilization in field conditions (Berry et al. 2010; Schulte auf´m Erley et al. 2011; Nyikako et al. 2014). This suggests that the enhancement of rapeseed performances under low N nutrition condition would primarily require the improvement of the NUpE. In other words, the size of the N pool in the aerial biomass conditions its remobilization to the seeds during the seed filling period. For instance, using a set of 30 diverse winter oilseed rape accessions grown under semi-controlled
0 50 100 A la s k a A ra g o n B e lu g a C o b ra C x C D H D a rm o r D ip p e s E x p e rt G ro ? L ? e w it z e r J u p it e r K ro m e rs k a L ib ra d o r L ib ri tt a M a d ri g a l M a jo r M a rk u s M e s tn ij M S L 0 0 7 O lim p ia d e O N D H 5 P a c if ic P ir o la R a p id R e s y n _ H 0 4 8 R e s y n G S 4 S a v a n n a h S k z iv e rs k ij S ta rt V iv o l W o ta n Genotype R o o t d ry w e ig h t (g ) N N1 N2 0 30 60 90 A la s k a A ra g o n B e lu g a C o b ra C x C D H D a rm o r D ip p e s E x p e rt G ro ? L ? e w it z e r J u p it e r K ro m e rs k a L ib ra d o r L ib ri tt a M a d ri g a l M a jo r M a rk u s M e s tn ij M S L 0 0 7 O lim p ia d e O N D H 5 P a c if ic P ir o la R a p id R e s y n _ H 0 4 8 R e s y n G S 4 S a v a n n a h S k z iv e rs k ij S ta rt V iv o l W o ta n Genotype R o o ts l e n g th ( c m ) N N1 N2 a b N+ N-N+
N-Figure I.A- 4. Genetic diversity for root length (a) and dry weight (b) measured on 30 WOSR genotypes grown in N+
vs. N- conditions
The genotypes were conducted as
described by Stahl et al. (in prep).
Figure I.A- 5. Root morphology at harvest between winter oilseed rape genotypes grown under semi-controlled conditions with a low N input
a: Expert genotype showing a small amount of lateral roots. b: Groß Lüsewitzer genotype with intense lateral root network. Both pictures were taken at day of seed harvest from nine plants grown in container under reduced N fertilization level (75 kg N.ha-1as an equivalent) (Stahl et al. in prep).
11 conditions, Stahl et al. (in prep) demonstrated that the NUE was significantly correlated with the shoot N amount (Figure I.A- 3-a ) as well as with the leaf N content at flowering (Figure I.A- 3-b). NUpE has to be investigated primarily at the beginning of the crop cycle for winter oilseed rape since a quarter to one third of the total N uptake is accumulated before winter (Cramer 1993) and most of the root system is fully developed before flowering (Barraclough 1989; Rahman and McClean 2013; Le Deunff and Malagoli 2014). Generally, the absorption of mineral ions by the plant depends first on their availability which is defined by the amount of elements present in the soil, the capacity of ions exchanges between the soil and the nutritive solution and the roots ability to take up the minerals. In the case of nitrate (NO3-) which is the main source of mineral N absorbed by plants, the soil retention is low and the roots action perimeter is wide (several cm). N absorption depends therefore essentially on the soil hydric reserves, the volume of soil investigated by the roots and the capacity of N absorption per unit root length. Genetic diversity of root weight, root length and root/shoot ratio was assessed in rapeseed at early stages of development and flowering time in field and controlled conditions on winter commercial varieties and DH lines (Kamh et al. 2005; Schulte auf'm Erley et al. 2007; Rahman and McClean 2013) and all assessed the importance of root length over the total root biomass. In addition, variations of the total root dry weight response to N constraints was evidenced at harvest within a 30 winter oilseed rape genotypes set (Stahl et al. in prep) (Figure I.A- 4-a and 1.4-b). As root growth and maintenance is costly in energy, the root architecture is a lever to optimize the balance between nitrogen absorption ability and metabolic costs (Lynch 2014). From this perspective, increasing the root surface by improving fine roots density was considered as one possible strategy in other crops (White et al. 2013) such as maize (Wiesler and Horst 1994), faba beans (Kage 1997) or Kentucky Bluegrass (Sullivan et al. 2000). Genetic diversity for root architecture was assessed in WOSR genotypes with the more N-efficient genotypes showing higher fine roots proportions (Figure I.A- 5) (Stahl et al. in prep). In addition contrasted root exploration profile was evidenced for two winter genotypes under field conditions (Albert et al. 2008), but the study of those traits remains fastidious and need further improvement. Although Kamh et al. (2005) suggested that NUpE of rapeseed is primarily determined by root growth rather than by the N uptake rate per root surface, this last trait must not be neglected. Genetic diversity for the activity of various nitrate and ammonium transporters which carry the N over membranes could be relevant for NupE in rapeseed (Schulte auf'm Erley et al. 2007) and should be further studied (reviewed in Xu et al. 2012).
Although a large body of the N amount is absorbed and stored in the shoot during the vegetative stage, N absorbed after flowering and during the seed filling period may sustain the plant N pool available, maintaining the global functioning of the plant, and may be remobilized later in the seeds.
12 The idea that the N taken up after flowering was negligible in rapeseed was questioned by Wiesler et al. (2001) who compared two winter oilseed rape genotypes following two different models of N use: on the one hand the “improved traditional ideotype” would show a better remobilization of N taken up before flowering, and on the other hand the “alternative ideotype” would show a higher NUpE during the reproductive stage. It was assessed in that study that the “alternative ideotype” showed better perspectives to improve the global NUE of rapeseed as seed yield was correlated with N uptake during reproductive growth and photosynthetic activity of leaves at the end of flowering. This was confirmed by several studies performed in the field that showed that post flowering NUpE had a significant effect on total NUE and was the most important phase to discriminate varieties for yield (Berry et al. 2010; Schulte auf'm Erley et al. 2011; Ulas et al. 2013). More precisely, Malagoli et al. (2005a) showed that 30% of the plant total N of a genotype grown in field condition was absorbed during seed filling, even though it represented only 27% of the total seed N. However, to our knowledge, no study associated the NUpE of rapeseed with the soil N resources at critical development stages such as flowering and seed filling stage, which may bias the actual post-flowering NUpE estimation as N is rarely available at that stage.
The N balance between source and sink organs needs optimization from the flowering stage onwards
Flowering is the paroxysm of the changes from vegetative to reproductive stage initiated at the end of the autumn period during the floral initiation (Figure I.A- 1) and is characterized by a massive change of source to sink relationships. The vegetative plant segments change from sink to source organs with associated modifications of N fluxes from older to younger tissues and reproductive organs (Le Deunff and Malagoli 2014). Labeling studies in Arabidopsis thaliana (Taylor et al. 2010) or B. napus (Rossato et al. 2001; Malagoli et al. 2005a) showed that the N accumulated in the seeds at harvest mainly originates from the protein degradation of plant organs. However, considering that the ramifications (main raceme vs. side branches) enter the generative phase sequentially, multiple overlapping source-sink situations are possible in rapeseed. This potential problem of synchronization between N source availability of old leaves and N needs in seeds along with an incomplete protein hydrolyze in the source organs lead to noteworthy N losses through fallen leaves (Aufhammer et al. 1994; Diepenbrock 2000; Rossato et al. 2001; Malagoli et al. 2005a; Ulas et al. 2013; Avice and Etienne 2014). Leaf senescence is therefore a critical stage at the crossroads between the improvement of N uptake and N remobilization in the reproductive organs (Schulte auf'm Erley et al. 2007).
13 Following the leaf senescence in rapeseed
Avice and Etienne (2014) pointed out the difficulty to precisely determine the progression of leaf senescence. While the direct measurement of the chlorophyll content is commonly used, several other biomarkers were tested to evaluate leaf senescence. For instance, Schulte auf’m Erley et al. (2007) showed a good correlation between SPAD meter values and photosynthetic rate. Gombert et al. (2006) developed a molecular method based on the kinetic expression of the two genes SAG12/Cab that were respectively up and down-regulated during leaf senescence. The inversion of the relative expression level of those two genes defines the beginning of leaf senescence. More recently, it was demonstrated that cellular structural changes associated with leaf senescence can be monitored by non-invasive 1H-NMR relaxometry (Musse et al. 2013; Sorin et al. 2015). In addition, the NMR signal captured choloroplast dismantling and was therefore a precise marker of leaf senescence (Sorin et al. 2015). Finally, the plant nutritional status was shown to impact leaf senescence and N remobilization abilities (Sorin et al. in prep).
The “stay-green” phenotype: an opportunity for the improvement of the source-sink relationships? Stay-green phenotype, i.e. mutants with altered chlorophyll catabolism (Gregersen et al. 2013), often refers to delayed leaf senescence in crop species (Thomas and Ougham 2014). It has been correlated with higher NUE in several species including Arabidopsis thaliana, maize and wheat (Spano 2003). Indeed, maintaining a high photosynthetic activity may allow the plant to carry on producing carbohydrates nutrients during late stages of the crop cycle that would be further remobilized to younger organs and seeds and may eventually increase the final yield (Habekotté 1993). Examples showing the relationships between the stay-green phenotype and the seed yield in several species were reported by Gregersen et al. (2013). However, the stay-green phenotype is not necessarily related to a prolonged photosynthetic activity. Indeed, in many cases, this phenotype translates an altered chlorophyll catabolism but with other senescence processes intact (Thomas and Howarth 2000). Hence, studies in rice and wheat reported no significant correlation or even negative correlations between the stay-green phenotype and the seed yield (see Gregerson et al. 2013 and references therein).
In rapeseed, the stay-green phenotype along with the LAI and RUE traits would be related with the span of the NUpE during the reproductive stage, as described previously (Diepenbrock 2000; Rathke et al. 2006) and may be a source of genetic diversity for the enhancement of the NUE. For instance, Schulte auf’m Erley et al. (2007) found significant genotypic difference for leaf senescence when comparing six contrasted N-efficient WOSR accessions under nutrient solution experiment and field
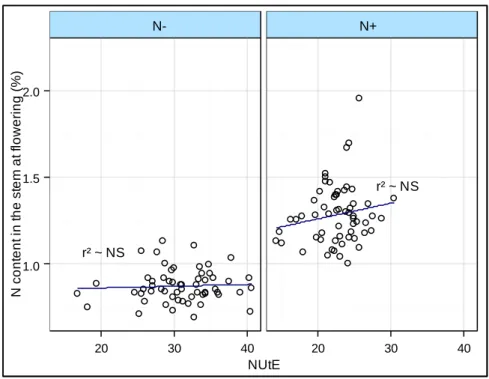
N1 N2 1.0 1.5 2.0 20 30 40 20 30 40 NUtE N c o n te n t in t h e s te m a t fl o w e ri n g ( % )
N content in the stem at flowering and NUtE correlation
r² ~ NS
r² ~ NS
N- N+
Figure I.A- 6. Correlation between NUtE and N content in the stems at flowering measured on 30 WOSR in N+ vs. N- conditions
14 conditions, and found positive correlations between delayed leaf senescence and N efficiency. In addition, Wiesler et al. (2001) found that the most N-efficient cultivar in their study showed better photosynthetic capacities at the end of flowering. However, contrary to other crop species such as cereals, the photosynthetic activity of the pods increases after flowering, making the relative contribution of leaf photosynthetic activity less important (Gammelvind et al. 1996). Besides, a late N remobilization may lead to greater N losses through leaves fall and high N:C ratio stems at harvest (Kaiser et al. 1998; Baggs et al. 2000). Hence, Koeslin-Findeklee et al. (2014) showed that the N-efficiency superiority of hybrids against inbred lines studied under N limitation was mainly related to the NRE and not correlated to delayed leaf senescence.
Nitrogen remobilization efficiency in rapeseed
According to Malagoli et al. (2005b), optimizing the NRE from vegetative to reproductive tissue could improve the yield by 15%. A first insight into the enhancement of the NRE would therefore be the improvement of the asynchrony between the N source availability and the N needs in the seeds. In rapeseed, the stems were described as N-storage organs that could compensate for this asynchrony (Hocking et al. 1997; Rossato et al. 2001; Malagoli et al. 2005a). Girondé et al. (2015) characterized a high NRE by a high amount of N in the stems at the beginning of the reproductive stage and a better N remobilization from the stems to the seeds. Genetic variations for the N stem remobilization were evidenced in rapeseed (Berry et al. 2010; Girondé et al. 2015) but were not always correlated to the NUtE under low N supply. This was supported by the data acquired on a 30 genotypes diversity set where no correlation was found between the N content of the stems at flowering and the NUE or the NUtE (Figure I.A- 6) (Stahl et al. in prep).
Tilsner and coworkers (2005) showed that phloem loading of amino acids was not limiting for the NRE and hypothesized that remobilization in rapeseed is rather bottlenecked by sink capacity than source strength. The final sink capacity of rapeseed can be first enhanced through the improvement of yield components per se. This was corroborated by several studies showing significant correlations between the HI and NUtE (Berry et al. 2010; Schulte auf´m Erley et al. 2011; Nyikako et al. 2014; Stahl et al. in prep), demonstrating that increasing the HI would increase the sink capacity for N and carbon. Increasing the total amount of proteins in the seeds may be another way to enhance the sink strength and the final NUtE of rapeseed. It appeared through several studies that genotypes with high NUtE values were characterized by low relative protein content of the seeds (Schulte auf´m Erley et al. 2011; Koeslin-Findeklee et al. 2014). Schulte auf’m Erley et al. (2011) showed that the genotypes with lower seed N content accumulated more dry matter in the seeds for a given amount of N accumulated, possibly explained by a higher photosynthetic rate of the pods or a higher
15 efficiency of oil production. At the molecular scale, the accumulation of a protease inhibitor, BnD22, in younger leaves in a low N context delays significantly their senescence and ensures the maintain of their sink strength (Etienne et al. 2007).
Which ideotype for an N-efficient rapeseed cultivar under field condition?
Both the N absorption and utilization are to be considered for the improvement of the global NUE in rapeseed (Ulas et al. 2013). They can be deconvoluted into sub-traits that were essentially studied at the plant level and showed a wide genetic variability.
The canopy architecture, involving interactions between the organs of a single plant as well as interactions between plants, must also be considered for efficient NUE breeding programs. For instance, the plant density and also the plant phenology may impact the total LAI and the photosynthetic activity of the lower leaves at the end of flowering (Malagoli et al. 2005a). Habekotté et al. (1997) proposed to combine traits like high LAI, apetalous flowers and erect pods to maximize the radiations intercepted by the plant and achieve high yielding. However, Zhao and Wang (2004) showed that the seed yield was not significantly higher in apetalous plants compared to their counterparts with petals, except when the plants were affected by Sclerotinia sclerotiorum.
As shown previously, the sink-source relationship seems to be the key of N remobilization in rapeseed, considering its undetermined growth. The timing of each development phase is therefore highly contributing to the final N-efficiency of the plants. The flowering marks the culmination of the switch of organ functions from N sink to source begun at the end of winter and conditions the efficiency of N remobilization from vegetative to reproductive organs. According to Malagoli et al. 2005a, an ideal genotype would exhibit early flowering in order to synchronize N mobilization with the N demand of pods. On the other hand, an early flowering may limit the vegetative growth of the plant and the N accumulated before flowering, which could cause a reduced N pool to remobilize afterwards.
Interactions between N and C metabolisms have to be taken into account when considering NUE. Thus, negative correlation between oil and protein content of the seeds has been assessed in rapeseed (Bouchet et al. 2014). As the main breeding objective is the oil yield, the question of the combination of those two opposite traits into breeding programs has to be considered. Indeed, an N-efficient genotype may result in a lower oil/protein ratio, especially in high N condition, which may not be the initial breeding goal. Hence, Masclaux-Daubresse and collaborators (2010) pinpointed that
16 breeding for delayed senescence could lead to higher yields but also to a decrease in NRE and protein content.
I.A- 3.
Breeding for N-efficient rapeseed cultivars with optimized oil yield:
where do we stand?
The G × E interactions have to be taken into account to orientate the breeding strategies
The relationships between the environments where the selection occurs (the selection environments) and the environments where the future varieties will be cultivated (the target environments) are major issues driving the breeding strategy. Indeed, on the one hand is the direct selection where the selection and target environments are closely related, and the varieties are therefore adapted to a narrow diversity of growing environments. On the other hand is the indirect selection where the selection and target environments are diverging and the varieties are more adapted to a wide panel of growing environments. However, indirect selection addresses the question of the transposition of trait breeding from one environment to another. Genotype × environment interactions (G × E) are therefore crucial in plant breeding and have been extensively studied in many crops.
In the case of NUE, the breeding strategy depends on the stress level of the target environment and the yield decrease between the selection and target environments. For instance, Bänziger et al. (1997) assessed that direct selection was preferable in maize when grain yield loss exceeded 43% between the selection and target environments. A moderately N-stressed target environment, with high genetic correlation between the selection and target environments, may therefore allow for indirect selection (Cormier et al. 2013). In rapeseed, the study of the interactions between the genotypes and the N nutrition regime (G × N) remains scarce with a limited number of reports (Gül 2002; Miro 2010; Bouchet et al. 2014). While Miro (2010) found significant QTL × N interactions on most of yield and NUE related traits, Gül (2002) and Bouchet et al. (2014) found a relative consistency of the QTLs between the N nutrition conditions. In addition, the QTLs detected in Miro (2010) and Bouchet et al. (2014) were varying between the different trials, demonstrating a strong effect of other environmental factors on N efficient loci. As a matter of fact, direct selection appears to be a convenient strategy for NUE breeding in rapeseed (Kessel et al. 2012), a statement that was also reported for wheat (Brancourt-Hulmel et al. 2005).
The characterization of the plant N stress in each environment studied is a prerequisite to evaluate the G × N interactions and to determine the breeding strategy for NUE related traits. In forage grass,
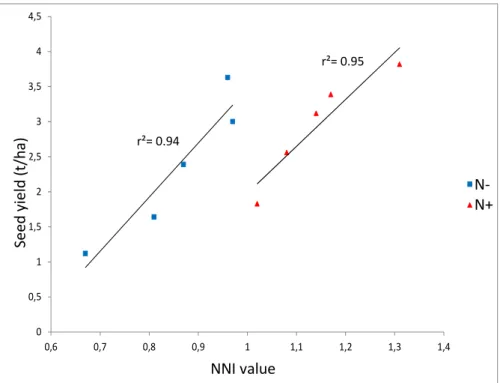
Figure I.A- 8. Correlation between NNI values and seed yield values for 92 winter oilseed rape genotypes grown under two contrasting N nutrition regimes (N- vs. N+) in Western France
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 0,6 0,7 0,8 0,9 1 1,1 1,2 1,3 1,4 S e e d y ie ld ( t/ h a ) NNI N1 N2 r²= 0.95 r²= 0.94 N-N+ S e e d y ie ld (t /h a ) NNI value N-N+ Figure I.A- 7. N dilution curve calculated for winter oilseed rape (Colnenne et al. 1998)
The critical N dilution curve (N=4.48 x DW-0.25) is represented by a black line. The two dotted lines represent the “envelope curves” (Nmax = 6.18 x DW-0.21; Nmin = 2.07 x
DW-0.17). The black dots represent N-limiting growth conditions; the white dots
17 Lemaire and coworkers (1984) assessed the negative relationship between the crop biomass and the minimum N concentration necessary to produce the maximum above ground biomass (the critical N concentration). This equation allowed establishing a theoretical critical dilution curve corresponding to the ideal nitrogen nutrition status for a range of aerial biomass. The ratio between the observed N concentration of a plant sample and the critical N concentration at a given aerial biomass, called the Nitrogen Nutrition Index (NNI), provides information on the N status of the sample studied. This index is commonly used to study NUE in major crops. For rapeseed, Colnenne et al. (1998) established the nitrogen dilution curve. Although it only implied one winter type genotype (Goeland) grown in different pedo-climatic regions of France, this curve still remains the reference to evaluate N nutrition status in rapeseed (Figure I.A- 7). More recently, a strong significant correlation was found between the NNI and the average seed yield of a diversity set of winter rapeseed lines grown in five environments (Figure I.A- 8).
Identification and characterization of N-efficient loci in rapeseed Genetic control of seed yield related traits
The genetic control of seed yield related traits by using linkage or linkage disequilibrium analyses has been extensively studied in rapeseed and came out with the identification of thousands of QTLs. The integration of those data may allow the identification of main genomic regions, potentially usable in breeding programs. For example, Zhou et al. (2014) integrated the information of 1960 QTLs for seed yield traits detected in 15 independent studies and identified 142 loci conserved between the different analyses and 25 multifunctional loci. This large body of information may also allow the identification of candidate genes. Few genetic analyses investigated the genetic control of seed yield related traits under N stress per se (Gül 2002; Miro 2010; Bouchet et al. 2014), but information is available on NUE related traits.
Genetic and molecular determinants of NUpE related traits
Several genetic studies addressed the genetic control of root system vigor in B. napus in the context of rapeseed adaptation to various environments (Rahman and McClean 2013) and especially under drought (Fletcher et al. 2014) and phosphorous stresses (Shi et al. 2012; Yang et al. 2010). The genetic control of root related traits, like vigor, biomass or length, was related to the flowering time, suggesting pleiotropic effects (Rahman and McClean 2013; Fletcher et al. 2014). Rahman and McClean (2013) hypothesized a trigenic dominant control of the root vigour based on the
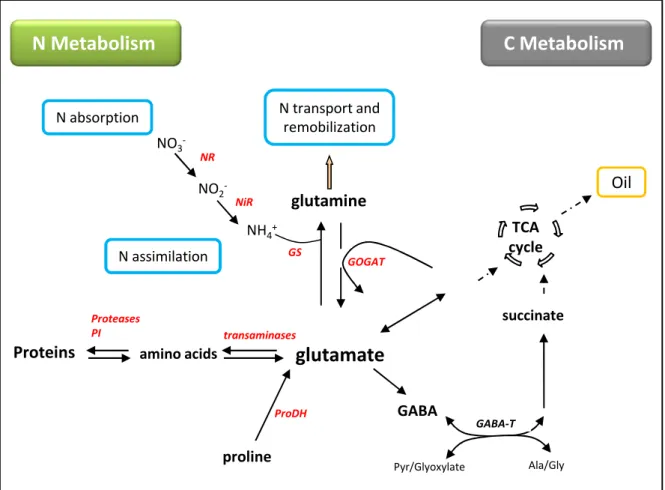
Figure I.A- 9. Schematic representation of the key steps of N metabolism in plants
Nitrogen is absorbed in the form of nitrate (NO3-) and assimilated through the nitrate reductase (NR) and nitrite reductase (NiR) pathways into the glutamine synthetase (GS) / glutamine oxoglutarate aminotransferase (GOGAT) cycle at the origin of the glutamate and glutamine synthesis.
ProDH: proline dehydrogenase; GABA: gamma-aminobutyric acid; PI: Protease inhibitor
NR ProDH proline GABA Pyr/Glyoxylate succinate GABA-T Ala/Gly
glutamate
Proteins
N Metabolism
amino acids Proteases PI transaminases TCA cycle GSglutamine
GOGAT N transport and remobilization NO3 -NH4+C Metabolism
NO2 -NiROil
N absorption N assimilation18 segregation ratio of a F2 population derived from reciprocal crosses between spring-type cv. and
winter-type canola lines. Yang et al. (2010) and Shi et al. (2012) detected clusters of QTLs for root length, lateral root number, root surface area, root biomass and root volume specific to the low phosphorous condition by linkage analyses. However, no study on root growth genetic control was performed under contrasted N conditions.
Although extensively studied in A. thaliana, little information is available on the genes implied in the root related traits and their regulation with N availability in rapeseed. In A. thaliana, root growth response to the availability of nitrate involves two distinct pathways (Zhang and Forde 1998, 2000): on the one hand, a direct effect of external nitrate on the MADS box transcription factor ARABIDOPSIS NITRATE-REGULATED1 (Zhang and Forde, 1998) and on the other hand, a systemic inhibitory effect by a basic Leu zipper and a LIM transcription factor (Tranbarger et al. 2003). More recently, the CLE-CLV1 signaling module was identified to play a crucial role in development of the lateral root system in N-poor environments (Araya et al. 2013). Additionally, nitrate transporters NRT1.1 (Remans et al. 2006a; Krouk et al. 2010) and NRT2.1 (Little et al. 2005; Remans et al. 2006b) are known to be crucial in nitrate sensing, independently from their uptake function (Malamy and Ryan 2001; Miller et al. 2007).
Genetic and molecular cues of NUtE related traits
As many other crops, the glutamine synthetase (GS) and nitrate reductase (NR) activities were associated to N transport in rapeseed (Figure I.A- 9). For example, Yea et al. (2010) found higher GS and NR activities in N-efficient genotypes under N stress condition. This was also supported by Miro (2010) who found that genes implicated in nitrate metabolism co-localized with N responsive QTLs. On the same line, Orsel et al. (2014) identified the genes encoding the cytosolic GS in rapeseed. In addition, two homoeologous genes (BnSAG12-1 and BnSAG12-2) coding for SAG12, a cysteine protease implied in N remobilization, were evidenced with their maximum expression level reached at early stages of leaf senescence (Noh and Amasino 1999). On the contrary, the down-regulation of the Cab gene (chlorophyll a/b binding protein) expression during leaf senescence was demonstrated in rapeseed. A recent study identified the genes differentially expressed between high and low N conditions in rapeseed and showed that N starvation promoted the down-regulation of genes related to photosynthesis, photorespiration and cell-wall structure while it enhanced the expression of genes related to mitochondrial electron transport and flavonoid synthesis (Koeslin-Findeklee et al. 2015). The polyploid nature of the B. napus genome implies the duplication of genes involved in NUE-traits within the genome and their organization into multigenic families, with possible variations in