Calcium and Rhizodermal Differentiation in
Primary Maize Roots
F. JAUNIN AND R. M. HOFER1
Institute of Plant Biology and Physiology, University of Lausanne, CH-1015 Lausanne, Switzerland
Received 7 October 1987
A B S T R A C T
Jaunin, F. and Hofer, R. M. 1988. Calcium and rhizodermal differentiation in primary maize roots.—J. exp. BoL 39: 587-593.
Rhizodermal differentiation of maize (Zea mays L. cv. LG 11) roots cultured in humid air was influenced by a pretreatment for 2 h in CaCl2 or CaSO* solutions. This increased the number of hair-producing roots and the density of hairs. Ethylene glycol-bis-{/?-aminoethyl ether)/VJV'-tetraacetic acid (EGTA) was inhibitory. Root hairs emerged in the part of the cell nearer to the tip. Trichoblasts were shorter and elongated more slowly than atrichoblasts. The elongation of the lower part of the trichoblast was less than that of the upper part.
Key words—Cell length, cell number, hair position.
Correspondence to: Institute of Plant Biology and Physiology, University of Lausanne, CH-1015 Lausanne, Switzerland.
I N T R O D U C T I O N
Cells of the rhizodermis may produce root hairs which are important in nutrient uptake from soils (Lamont, 1982). In roots grown in different nutrient solutions, the length of the hairs depends among other factors on the pH and the concentrations of Ca2 + and nitrate
(Cormack, 1949; Ewens and Leigh, 1985). When roots are grown in solutions without added calcium, their growth is severely inhibited (Pilet and Belhanafi, 1961; Tanaka and Woods, 1972; Ferguson and Bollard, 1976; Burstrom, 1981; Ewens and Leigh, 1985) and the density and length of root hairs are reduced (Cormack, 1949; Burstrom, 1952; Tanaka and Woods, 1972; Ewens and Leigh, 1985). Nevertheless, Ekdahl (1953) reported that the concentration of exogenous calcium was not a determining factor; in moist air, without an external supply of calcium, root hairs developed very well. In maize roots cultured in humid air without applied calcium, some roots produce no hairs at all and the presence or absence of a hairy zone is not related to root growth (Jaunin and Hofer, 1986).
In most species, root hair forming cells, usually called trichoblasts, are distinguishable from hairless cells (atrichoblasts) on the basis of cell size, activities of several enzyme systems, density of the cytoplasm and the amount of RNA, nuclear DNA, histones and proteins (Avers, 1961; Cutter and Feldman, 1970; Dosier and Riopel, 1977,1978; Harris, 1979). In Zea mays roots, Clarke, McCully, and Miki (1979) did not observe any cytoplasmic or nuclear differences between trichoblasts and atrichoblasts. Moreover, no definite relation was
1 To whom correspondence should be addressed. © Oxford University Press 1988
observed between length of the rhizodermis cells and their capacity for producing hairs (Cormack, 1949; Clarke et al., 1979). In this material grown in humid air, cell divisions in the rhizodermis were asymmetrical with a short apical cell and a large basal cell (Ivanov and Filippenko, 1979).
In a study of cell polarity and the emergence of hairs in maize roots, Clarke et al. (1979) observed that the hairs form at the acropetal end of the rhizodermal cell. The final position of the hair on the cell depends on the growth of the different parts of the cell (Cormack, 1949). The aim of the present paper was, first, to investigate whether calcium influences the differentiation of the rhizodermis into trichoblasts and atrichoblasts in maize roots cultured in humid air and, secondly, to determine the size of the rhizodermal cells and the position of the hair on the cell along the root axis.
MATERIALS AND M E T H O D S
Plant material
The germination of Zea mays L. caryopses (cv. LG 11) has already been described (Jaunin and Hofer, 1986). After 48 h, intact seedlings with straight primary roots (14± 1-0 mm) were selected in dim green light [530 ± 20 nm, 1-2 x 10* W m~2 (Pilet, 1979)]. As observed with the scanning electron microscope, no outgrowths due to emergence of root hairs could be detected.
Roots attached to the caryopsis were immersed vertically for 2 h in a buffered solution (1-0 mol m~3 3,3-dimethylglutaric acid-NaOH, pH 6-0, 2-0 cm3 per root) without (control) and with CaCl2 (1-0 mol m ~3 or 10 mol m"3) or CaSO4 (10 mol m~3), or ethylene glycol-fcis-(/?-aminoethyl ether)N,iV'-tetraacetic acid (EGTA) (0-1 mol m~3). Roots were then rinsed for 1 s in the control solution. These solutions were prepared with double-distilled water and filter-sterilized, using 0-45 fan niters (Millipore filter unit). Then roots attached to the caryopsis were placed in a vertical position in sealed boxes for the following 20 h (dark, saturated atmosphere, 21 °Q.
Growth measurements
Root growth was recorded for 20 h in humid air by taking photographs every 2 h (light provided for 7 s per photograph by white fluorescent tubes, Philips 20 W, 9-1 x 10~2 W m"2) as already described (Jaunin and Hofer, 1986).
Location of the hairy zone
After 20 h in humid air, the roots were excised and individually fixed in 2% (v/v) glutaraldehyde (4 CC). Measurements were made, using a binocular microscope (Wild M5, x 12, x25) with an eye-piece graticule.
Determination of cell number, cell dimensions and position of hairs on the cells
The material was fixed overnight in glutaraldehyde at 2% (4°Q. After dehydration in a series of acetone baths, roots were dried through the C O2 critical point, sputter-coated with gold (about 70 nm) and viewed in a JEOL JSM-35 SEM at 25 kV (Centre for Electron Microscopy, University of Lausanne). Measurements were done on photographs (final enlargement 310 x ) in consecutive 0-75 mm segments along the root axis. This was performed between 3 0 mm and 6-75 mm counted from the tip. In regions distal to 3 0 mm, it was difficult to identify cell limits because of the homogeneity of the outer wall (Hofer and Pilet, 1986). In more basal regions, cell sizes could not be measured due to the overlapping of root hairs. Cell measurements were realized using a digitizing pad (Hi PAD, Houston Instruments, Austin, TX, U.S.A.) interfaced with a microcomputer (ABC 80, Luxor AB, Motala, Sweden). The number of cells which give rise to hairs was expressed in % of total number of cells. Cells which had an emerged root hair were counted as trichoblasts; the others were counted as atrichoblasts, even though a hair could be expected to form in some cells.
RESULTS
Calcium and formation of root hairs
The concentrations of calcium salts and of the calcium-chelating agent EGTA (Schmid and Reilley, 1957) were chosen not to affect the elongation rate of roots compared with control
T A B L E 1. Relative number (in %) of roots producing hairs Pretreatment Percentage (%) Buffer (control) CaCl2 1-0 mol m~3 CaCl2 10 mol m~3 CaSO4 10 mol m~3 EGTA 01 mol m"3 66 (56-77) 83 (73-100) 97 (92-100) 100 38 (22-57) Roots attached to the caryopsis were immersed 2 h in buffer without and with CaCl2) CaSO4 or
EGTA and then transferred to humid air for the following 20 h.
The numbers of roots tested were 84 for the control and 30 for the other pretreatments. Con-fidence intervals (P = 0-05) are in brackets.
specimens (pre-incubated in buffer). The elongation rate of the latter as a function of time has already been given (Jaunin and Hofer, 1986); the mean growth rate over 20 h was 0-85 mm h"1.
Hairs were produced in 66% of control roots (Table 1). Although the amounts of free Ca2 +
initially available in the solution would not be the same for CaG2 and CaSO4, these two salts
(at 10 mol m"3) both significantly increased the percentage of roots with hairs. The difference
observed between the control roots and the roots treated with 10 mol m"3 CaCl2 was not
significant. The calcium-chelating agent EGTA at 01 mol m~3 significantly decreased the
percentage of hair-producing roots.
After 20 h, calcium salts and EGTA did not affect the length of the hairy zone, but root hairs formed nearer to the tip only when roots were pre-incubated with CaCl2 at 10 mol m ~3
(Table 2).
Since there was no significant difference between the number of hair producing roots with CaCl2 and CaSO4, subsequent experiments were performed with CaCl2 which is almost
entirely dissociated in solution at 10 mol m~3.
Root hair density was studied along the root axis for material pre-incubated for 2 h in CaCl2 at 10 mol m~3 and then transferred to humid air for the following 20 h. Results are
T A B L E 2. Length of the apical zone without hairs (zone 1) and of the hairy zone (zone 2) Pretreatment Buffer (control) CaCl2 1-0 mol m~3 CaClj 10 mol m~ EGTA 01 mol m"3 Length (mm)±s.e. Zone 1 4-4 ±03 3-8±01 3-3±01'1 3-8 ±02 Zone 2 15-0±O6 17-3 ± 0 8 15-0 + O8 14-5±1-1 Roots attached to the caryopsis were immersed 2 h in buffer without and with CaCl2, or EGTA and then
transferred to humid air for the following 20 h. " Value significantly different from control (P = 0-05).
590 UJ
I
UJ 50 30 20 10 1 \ -/ i i i i i CN /X /
\ /
'p 1 1 1 1 - 400 - 300 o2
a. hi a. 200 3 z o o - 100 UJ o 3 4 5 6 7DISTANCE FROM THE TIP IN mm
FIG. 1. Cell number (CN) counted per root (± s.e.) and percentage (P) (± s.e.) of cells which had given rise to hairs along the root. Roots attached to the caryopsis were immersed 2 h in buffer with CaCl2 at 10 mol
m"3 and then transferred to humid air for the following 20 h. Number of roots tested 10.
given as percentages of the total number of cells since cell number decreases towards the root base (Fig. 1). These values increased rapidly from 3-0 mm to 5-25 mm in the growing part of the root and reached a constant value at 6-0 mm from the tip where growth is about to cease (Pilet, Versel, and Mayor, 1983). Consequently, emergence of root hairs occurred in the extending part of the root and above this region cells cannot give rise to hairs (Jaunin and Hofer, 1986). In roots which had a hairy zone, percentages of cells which had given rise to hairs in control roots were compared with CaQ2-treated and EGTA-treated roots (Table 3).
This was performed 6'0 mm from the tip where the density of root hairs was highest. In this zone, in control roots, less than half of the cells formed hairs. This percentage was significantly higher for roots pre-incubated with CaCl2 and significantly lower for roots
pre-incubated with EGTA.
T A B L E 3. Relative number (in %) of cells which had given rise to hairs 6-0 mm from the tip Pretreatmcnt Percentage (%± s.e.)
Buffer (control) CaCl2 10 mol m"3 EGTA 01 mol m"3 43±1 54±1 30±l
Roots attached to the caryopsis were immersed 2 h in buffer without and with CaCl2, or EGTA and then
transferred to humid air for the following 20 h. Cell number counted: 2 000 for each pretreatmenL
591
Length of rhizodermal cells
The lengths of rhizodermal cells along the root axis were studied with scanning electron microscopy on roots pre-incubated 2 h in buffer with CaCl2 at 10 mol m"3 and then
transferred to humid air for the following 20 h (Fig. 2). Trichoblasts and atrichoblasts already showed significant differences in their length 3-0 mm from the tip, where the most apical hairs were found. The length of trichoblasts progressively increased between 30 mm and 5-25 mm from the tip and reached a constant value at 5-25 mm. The increase in cell length of atrichoblasts was larger and a constant value reached only at 60 mm. Consequently, the difference in length between trichoblasts and atrichoblasts depends on the region observed. Their extension growth between 30 to 6-75 mm behind the tip was 83% and 144% respectively.
0.20
o 0.05
3 U 5 6 7 DISTANCE FROM THE TIP IN mm
Fie. 2. Length (in mm ± s.e.) of trichoblasts (T) and atrichoblasts (A) along the root. Roots attached to the caryopsis were immersed 2 h in buffer with CaCl2 at 10 mol m "3 and then transferred to humid air for the
following 20 h. Number of roots tested 10.
Position of the hairs on trichoblasts
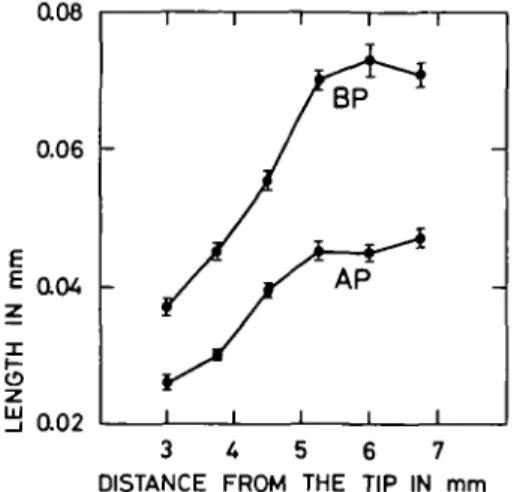
The position of the hair on the trichoblast is shown in Fig. 3 for roots pre-incubated in CaCl2 at 10 mol m"3. The distances from the apical and basal ends of the cell to the centre of
the hair were measured along the root axis. At 30 mm from the root tip, root hairs emerge nearer the apical end of the hair-forming cell. The basal and apical parts of the cell increased in length at the same rate when less than 4-5 mm from the root tip. From 4-5 mm to 5-25 mm, the length of the basal part of the cell increased more rapidly than the apical part. Consequently, the position of the hair is even further from the middle of the cell. A constant value for both parts was reached at 5-25 mm behind the tip. The total increase was 80% for the apical part and 92% for the basal part
DISCUSSION
The results show that under the growth conditions used, the differentiation of the rhizodermis into trichoblasts and atrichoblasts is dependent on calcium salts in maize roots
0.08
0.02
3 U 5 6 7 DISTANCE FROM THE TIP IN mm
FIG. 3. Position of the hair on the trichoblast along the root axis. AP: apical part of the cell to the hair centre (in mm±s.e.); BP: basal part of the cell to the hair centre (in mm±s.e.). Roots attached to the caryopsis were immersed 2 h in buffer with CaCl2 at 10 mol m ~3 and then transferred to humid air for the
following 20 h. Number of roots tested 10.
grown in humid air. EGTA at 0-1 mol m 3 did not completely inhibit rhizodermal
differentiation. Perhaps this concentration does not complex all the calcium involved in differentiation. Preliminary tests have shown that higher concentrations of this calcium-chelating agent prevent root hair formation, but also affect root elongation and even seem to be toxic for the roots. Trichoblasts are shorter than atrichoblasts and length curves along the root axis are different for the two types of cells. The difference in size was observed already at 3-0 mm from the tip; this could be due to asymmetrical divisions (Ivanov and Filippenko, 1979).
Root hairs always originate nearer the apical end of the cell. Similar results were obtained by Clarke et al. (1979) for maize roots. Moreover, the parts of cell apical and basal to the hair have different rates of elongation. It was reported in Gibasis geniculata roots that the site of root hair inception might be determined by the intrinsic polarity of the cell (Nakazawa and Yamazaki, 1982). Clarke and McCully (1985) noted a structural change in rhizodermal cell walls of Zea mays during hair initiation and observed a strongly callose-positive papilla proximal to an emerged root hair or nearer the site where a hair would be expected to form. Moreover, callose synthesis can be regulated by calcium (Kauss, 1985). With electron probe X-ray microanalysis, a large amount of calcium was observed in the peripheral layers of Nicotiana tabacum roots (Scheidecker, Chevalier, Andreopoulos-Renaud, Bizid, and Boumati, 1981) and in the epidermis of Zea mays roots (Chino, 1981). Further studies are required to investigate the ways in which calcium influences rhizodermal differentiation. A C K N O W L E D G E M E N T S
We are very grateful to Professor P. E. Pilet for critical reading of the manuscript and stimulating discussion. We also wish to thank Dr H. V. Martin for helpful comments.
LITERATURE CITED
AVERS, C. J., 1961. Histochemical localization of enzyme activities in root meristem cells. American
Journal of Botany, 48, 137-43.
BURSTRSM, H. G., 1952. Studies on growth and metabolism of roots. Vlll. Calcium as a growth factor.
1981. Mineral nutrition and growth responses of Pisum roots. Zeitschrift fur Pflanzenphysiologie, 102, 451-6.
CHINO, M., 1981. Species differences in calcium and potassium distributions within plant roots. Soil
Science and Plant Nutrition, 27, 487-503.
CLARKE, K. J., and MCCULLY, M. E., 1985. The occurrence of wall papillae in root epidermal cells of axenically grown seedlings of Zea mays. American Journal of Botany, 72, 1483-9.
and MIKJ, N. K., 1979. A developmental study of the epidermis of young roots of Zea mays L.
Protoplasma, 98, 283-309.
CORMACK, R. G. H., 1949. The development of root hairs in angiospcrms. The Botanical Review, 15, 583-612.
CUTTER, E. G., and FELDMAN, L. J., 1970. Trichoblasts in Hydrocharis. I. Origin, differentiation, dimensions and growth. American Journal of Botany, 57, 190-201.
DosffiR, L. W., and RIOPEL, J. L., 1977. Differential enzyme activity during trichoblast differentiation in
Elodea Canadensis. Ibid. 64, 1049-56.
1978. Origin, development and growth of differentiating trichoblasts in Elodea Canadensis. Ibid. 65, 813-22.
EKDAHL, I., 1953. Studies on the growth and the osmotic conditions of root hairs. Symbolae Botanicae
Upsalienses, 11, 5-83.
EWENS, M., and LEIGH, R. A., 1985. The effect of nutrient solution composition on the length of root hairs of wheat (Triticum aestivum L.). Journal of Experimental Botany, 36, 713-24.
FERGUSON, I. B., and BOLLARD, E. G., 1976. The movement of calcium in germinating pea seeds. Annals
of Botany, 40, 1047-55.
HARRIS, W. M., 1979. Ultrastructural observations on the trichoblasts of Equisetum. American Journal
of Botany, 66, 673-84.
HOFER, R. M., and PILET, P. E., 1986. Structural and cytochemical analysis of the cell walls in growing maize roots. Journal of Plant Physiology, 122, 395-402.
IVANOV, V. B., and FILIPPENKO, V. N., 1979. Symmetry in mitotic divisions and differentiation of cells of the rhizoderm in maize. Doklady Biological Sciences, 245, 763-5.
JAUNIN, F., and HOFER, R. M., 1986. Root hair formation and elongation of primary maize roots.
Physiologia plantarum, 68, 653-6.
KAUSS, H., 1985. Callose biosynthesis as a Ca2+-regulated process and possible relations to the induction of other metabolic changes. Journal of Cell Science (Suppl.), 2, 89-103.
LAMONT, B., 1982. Mechanisms for enhancing nutrient uptake in plants, with particular reference to mediterranean South Africa and Western Australia. The Botanical Review, 48, 497-689. NAKAZAWA, S., and YAMAZAKI, Y., 1982. Cellular polarity in root epidermis of Gibasis geniculata.
Naturwissenschaften, 69, 396-7.
PILET, P. E., 1979. Kinetics of the light-induced georeactivity of maize roots. Planta, 145, 403-4. and BELHANAFI, A., 1961. Action auxinique et chelation. Annales de Physiologie Vegetale, 3, 73-86.
VERSEL, J. M., and MAYOR, G., 1983. Growth distribution and surface pH patterns along maize roots. Planta, 158, 398-402.
SCHEIDECKER, D., CHEVALIER, S., ANDREOPOULOS-RENAUD, U., BIZID, A., and BOUMATI, P., 1981. Localisation histologique du phosphore, du calcium et du potassium au cours de revolution des racines de Tabac. Physiologie Vegetale, 19, 237-52.
SCHMID, R. W., and RHLLEY, C. N., 1957. New complexon for titration of calcium in the presence of magnesium. Analytical Chemistry, 29, 264-8.
TANAKA, Y., and WOODS, F. W., 1972. Root and root hair growth in relation to supply and internal mobility of calcium. Botanical Gazette, 133, 29-34.