Preconditioning of the Tumor Vasculature and Tumor Cells by Intermittent Hypoxia: Implications for Anticancer Therapies
Texte intégral
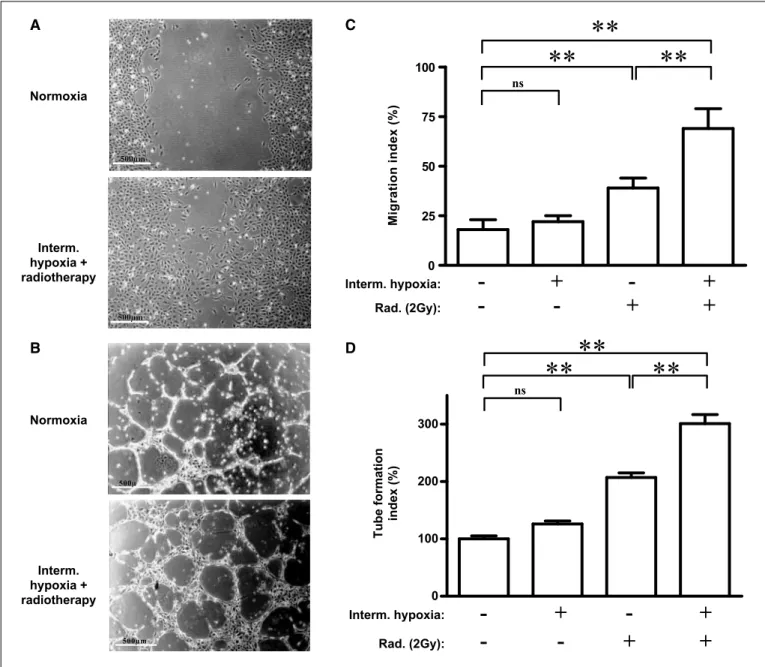
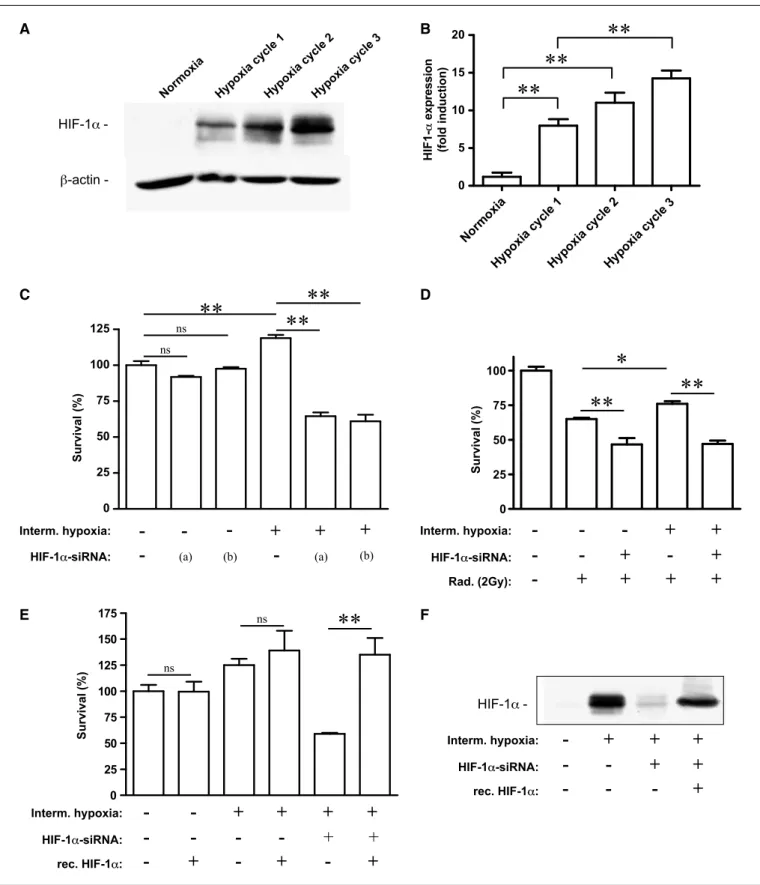
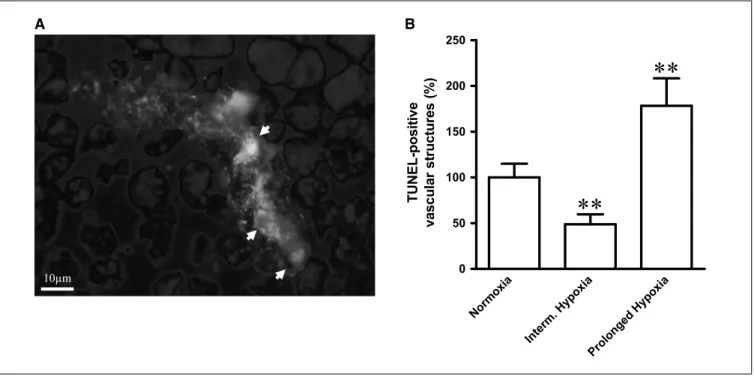
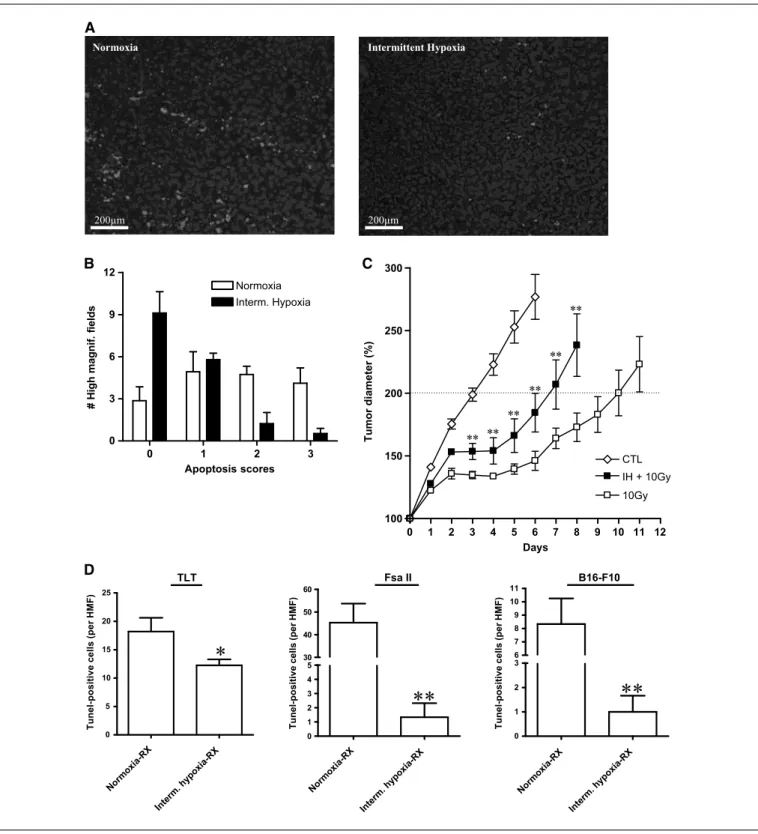
Figure

Documents relatifs
Enriching Treebanks with a Constraint-Based Description Our treebank enrichment consists in building an hybrid syntactic representation, one (the original) being
Experiments on the bioreporter icl-luc tested the expression of icl under various levels of Fe-limitation and different sources of organic carbon, while comparative experiments
Molecular characterization of circulating tumor cells from patients with metastatic breast cancer reflects evolutionary changes in gene expression under the
Au premier plan, à gauche, soldat romain armé d'une lance, à droite, trois personnages assis au bord d'un étang.. Au second plan,
Interferon gamma, an important marker of response to immune checkpoint blockade in non-small cell lung cancer and melanoma patients.. Ther Adv
Il y a une pilule pour réduire votre risque s’il est probable que vous soyez exposé au VIH. Parlez à votre fournisseur de soins de santé de la prophylaxie pré-exposition
Similarly, high lactate levels were shown to promote an aggressive phenotype in breast cancer cells [107] and have been associated with a more stem cell-like gene expression profile
