Diverse cuticular remains in Cambrian (Series 2) SSF assemblages from China and the pioneer metazoan colonization of offshore environments
Texte intégral
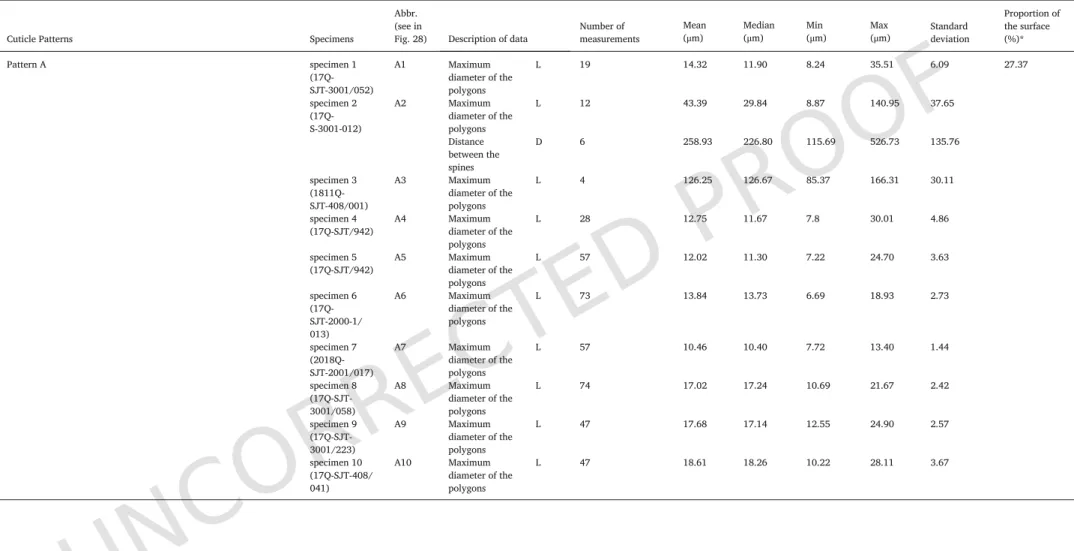
Figure









Documents relatifs
The Lower Devonian Posongchong Formation (Wenshan, Yunnan Province, southwestern China) comprises a series of continental clastic deposits, from which 28 vascular plant gen- era
Using UniFrac analysis, we showed that phosphorus con- centration (lake trophic state) was related to the changes in the global phylogenetic composition (16S OTUs) of the
To determine how the IAA treatment affected the sensory retina, the number of rows of nuclei in vertical sections of the outer and inner nuclear layer was counted in four locations
In order to understand the relationship between genetic architecture and adaptation, it is relevant to detect the association networks of the candidate SNPs with climate variables
After a hiatus of half a century, renewed fieldwork in the Cretaceous–Paleocene of the Pindos Unit in Eurytania, continental Greece, brought to light new localities and ray-finned
First, comparing with inflation where the primordial fluctuations become frozen at the moment of the Hubble exit, those primordial fluctuations on matter fields in bounce cosmology
The more or less explicit rationale that underlies process-outcome research is "the idea that teacher behaviour is the 'cause' and student learning is the 'ef- fect' (which)
New fossil remains of Fusuisaurus zhaoi (Sauropoda: Titanosauriformes) from the Lower Cretaceous of Guangxi, southern China... Natural History Museum of Guangxi Zhuang
