Explaining fine-grained properties of human cooperation : Insights from evolutionary game theory
Texte intégral
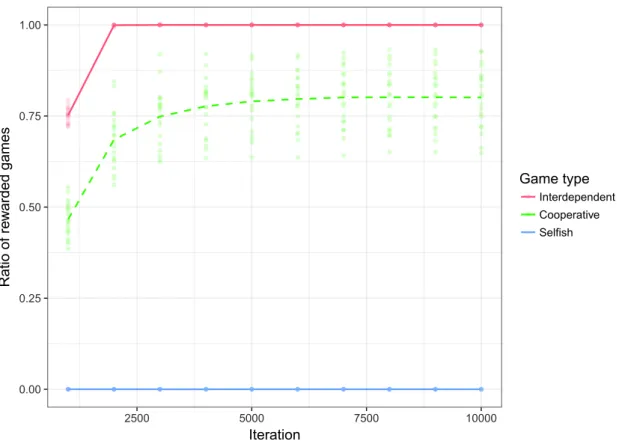
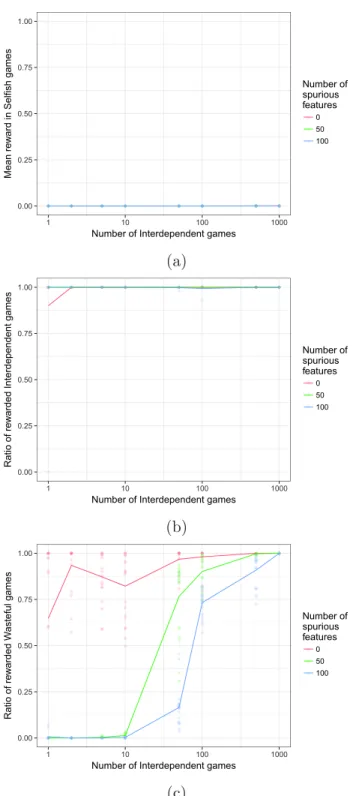
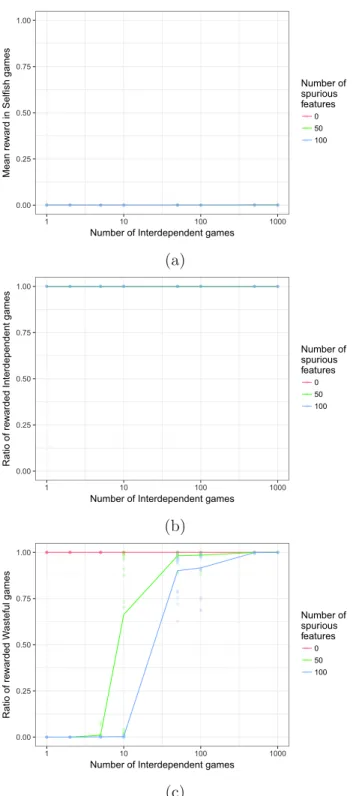
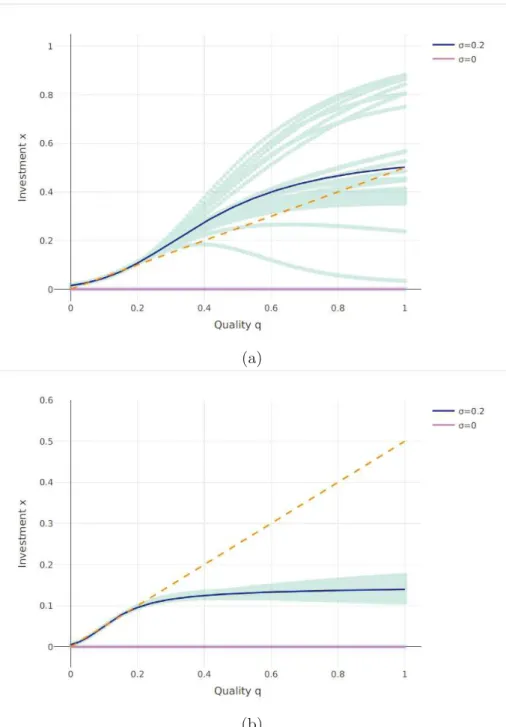
Figure


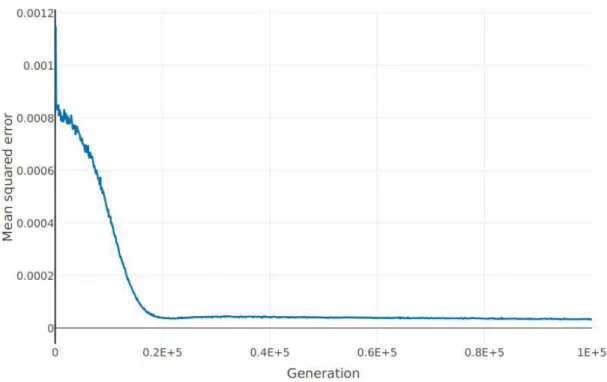
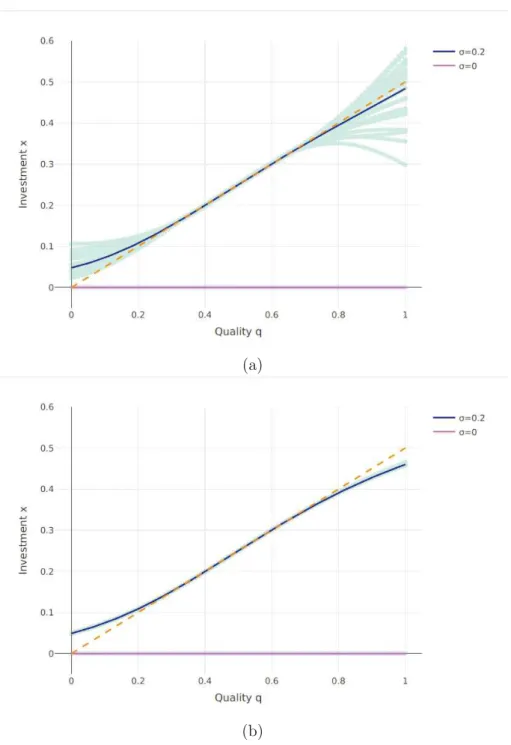
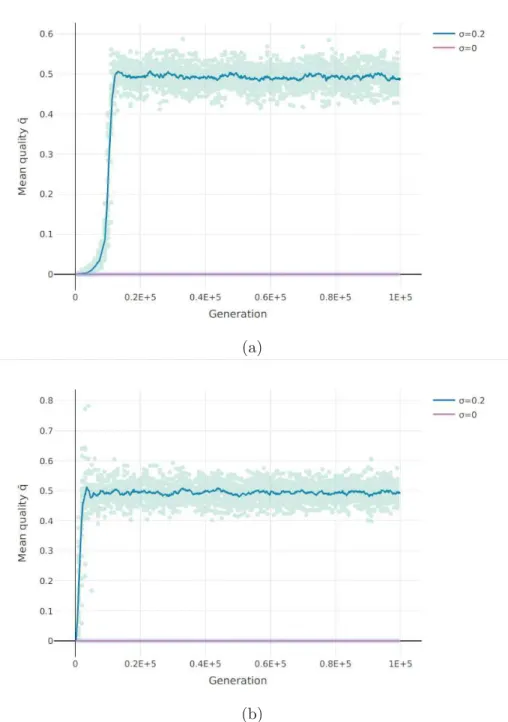
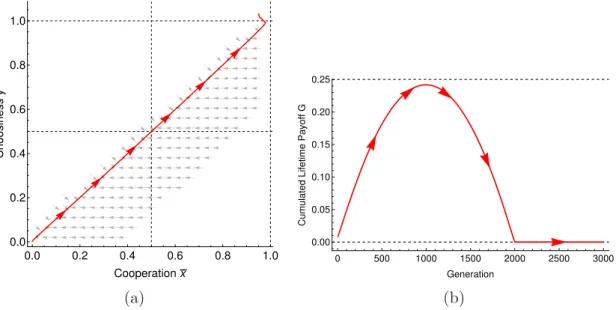
Documents relatifs
The forwarding time of cooperative users at position jth can be computed the same as Eq. The develop- ment is similar to that in the above computations. The messages marked with a
Segmentation results of individual and cooperative approaches applied to a real image issued from the BCU database [22] (a) Original image (b) Grayscale image (c) Active Contour
• We work out the limits of our techniques in two well-known dead ends of game semantics: non-associativity of composi- tion in Blass games [4] and non-stability of innocent
In light of this, we here focus on the last millennium (the tenth to twen- tieth centuries AD) to compare the historical events with re- lated geological findings and to
Keywords: agricultural economics, environmental and resource economics, exhaustible resources, growth and sustainable development, green paradox, environmental policies..
Tout d'abord, la prise de position de Louis sur les Roms demeurera- t-elle un incident unique en son genre ou annonce-t-elle, au contraire, une radicali- sation du parti,
Method that Produces Global Precipitation Estimates from Pas- sive Microwave and Infrared Data at High Spatial and Temporal Resolution, J. Hydrometeorol., 5,
Midi Minuit 3:10 PM MathsLibres.com... Midi Minuit 3:10
