A feedback control perspective on biological control of dengue vectors by Wolbachia infection
48
0
0
Texte intégral
Figure


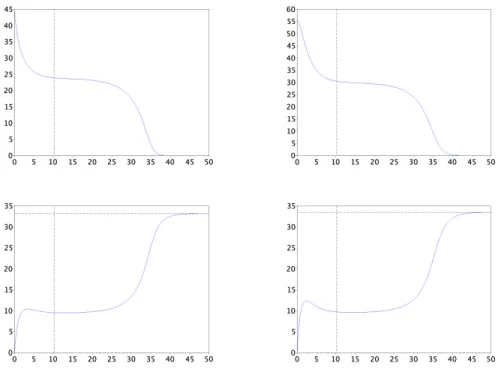
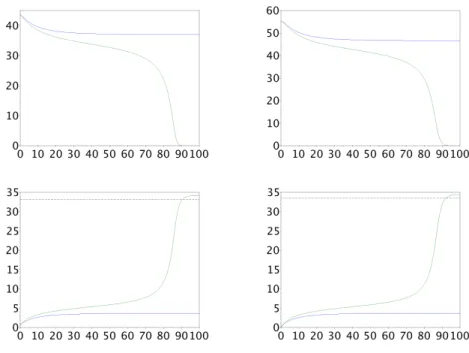
+4

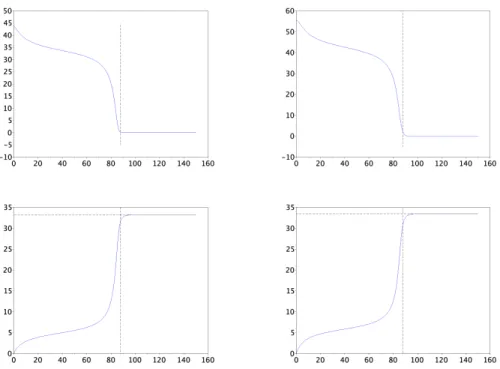

Documents relatifs