Shared premotor activity in spoken and written communication
Texte intégral
Figure
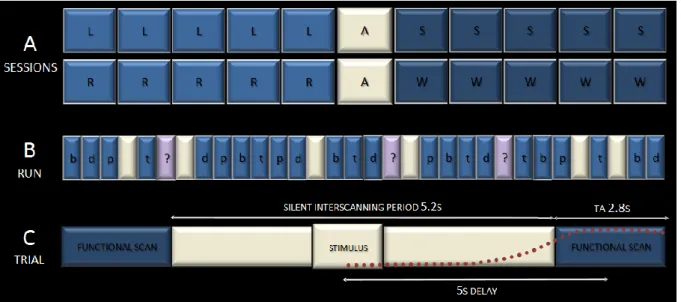
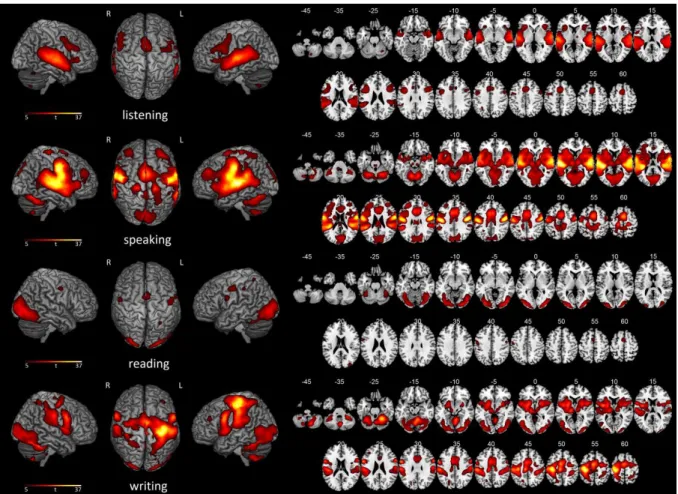



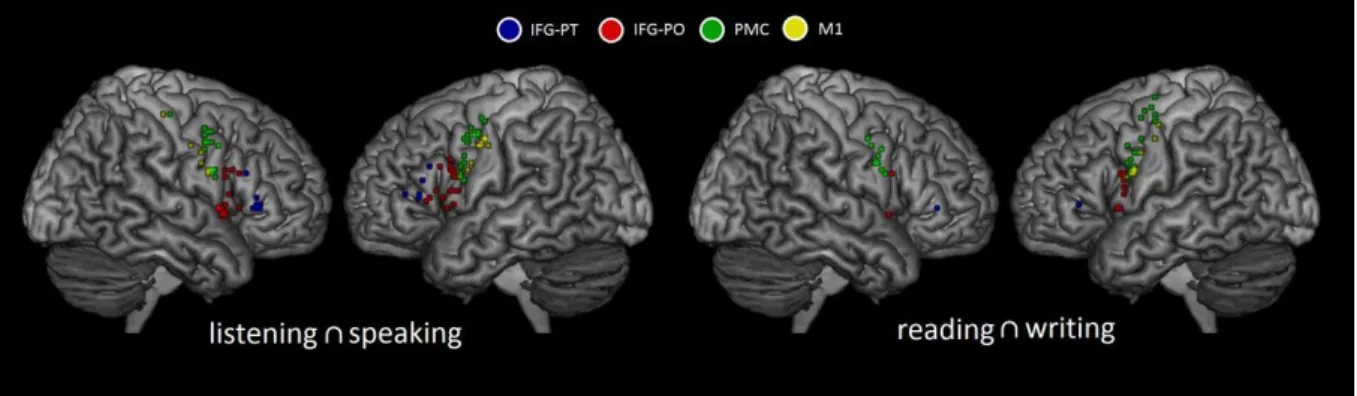
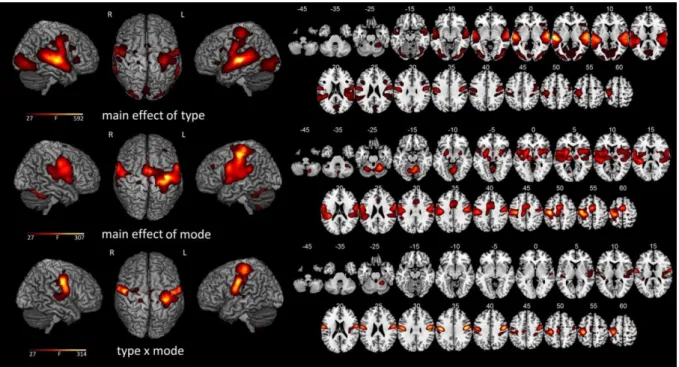
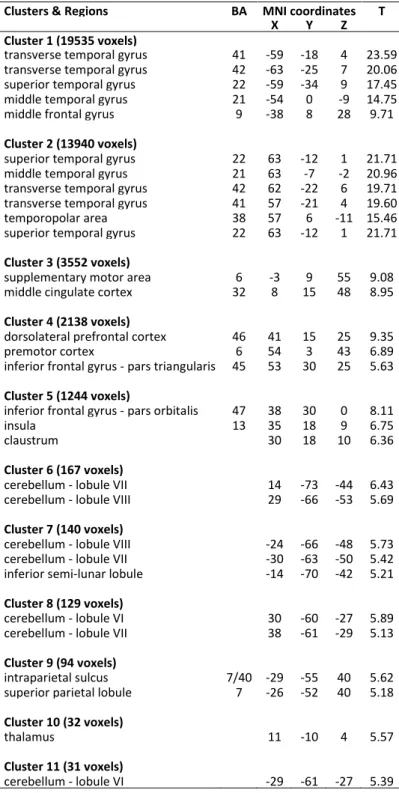
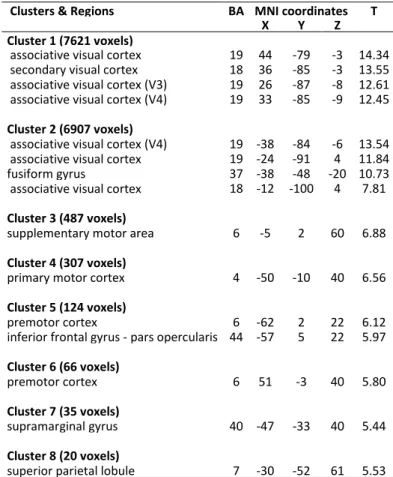
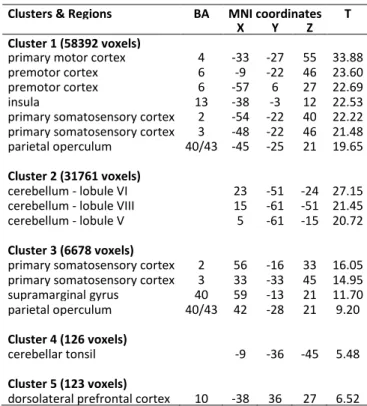
Documents relatifs
In this work, we follow the same general scheme, but we apply support vector regression to obtain a simpler surrogate model and we compare our preliminary results to those obtained
The systems have been trained on two corpora (ANCOR for spoken language and Democrat for written language) annotated with coreference chains, and augmented with syntactic and
The second cluster contained the following five grouped variables: SI1 (student-student interaction), IL3 (fostering metacognition), IL4 (benefit from the independent
Tracing of mono- and polysynaptic afferent connections between the main olfactory bulb and higher- order brain regions in the mouse. XXVth Annual Meeting of the European
In the simplest case of the planar JJ with the vortex trapped in one of the superconducting electrodes near the Josephson barrier, the vortex currents cross this barrier and induce
Du point de vue clinique, on associe cette méthylation du promoteur de SAMHD1 à la réduction de l’expression de a protéine dans certaines cancers (Wang et al., 2014). Mes
Integra- tions of kinetic equations of Alfve´n wave turbulence re- cover the predicted laws, while our MHD numerical flows follow them in planes at lowest parallel wave
Finally, let us mention that small ball estimates play a rˆole in other problems, such as invertibility of random matrices and convex geometry.. [RV09]
