i
Résilience des pessières à mousses du Québec aux
incendies peu sévères :
Conséquences pour le cycle du carbone à long terme
Thèse
Juliette BOIFFIN
Doctorat en sciences forestières
Philosophiae Doctor (Ph.D.)
Québec, Canada
© Juliette Boiffin, 2014
iii
Résumé
La dynamique du carbone en forêt boréale est contrôlée par celle des incendies qui consument partiellement la biomasse et la matière organique du sol, libérant du carbone dans l’atmosphère. À long terme, une perte de résilience peut conduire les forêts brûlées à évoluer vers des écosystèmes présentant différentes compositions, structures et cycles biogéochimiques. Les modèles prédictifs de la dynamique du carbone en forêt boréale simulent rarement ces différentes trajectoires successionnelles, ce qui biaise les prévisions à long terme. Nous avons étudié la résilience aux incendies et la dynamique du carbone des pessières à mousses du Québec. La régénération de l’épinette noire, du pin gris et du sous-bois a été inventoriée suite à trois années d’importante activité des feux dans la province. Trois à cinq ans après feu, une épaisse couche de matière organique résiduelle avait compromis l’établissement de l’épinette noire, provoquant une ouverture du couvert et un changement de dominance vers le pin gris. Les éricacées s’étaient régénérées en abondance dans la plupart des parcelles. La composition du sous-bois après feu était principalement déterminée par les caractéristiques du site et du peuplement. Elle reflétait les assemblages d’origine, les legs biologiques ayant été préservés par la faible sévérité des incendies. Nous avons ensuite modélisé les feux, la succession et la dynamique du carbone à l’échelle du paysage pour quantifier l’impact de la sévérité du feu et de la régénération des arbres sur le cycle du carbone. Une diminution de 13 % du carbone stocké dans le paysage était prédite au bout de 500 ans lorsqu’on simulait la sévérité du feu et son impact sur la régénération. Cette diminution était plus fortement liée à la faible régénération de l’épinette noire qu’à la combustion de la matière organique en elle-même. Les émissions liées au feu étaient déterminées par les variations de la surface annuelle brûlée. Ainsi, la perte de résilience des pessières à mousses suite aux feux légers affecte fortement le stockage du carbone à long terme. Le modèle utilisé pourrait encore être amélioré en incorporant la dynamique du sous-bois.
v
Abstract
Wildfire is a major driver of carbon dynamics in boreal forests. Immediate effects of wildfires include partial consumption of aboveground biomass and the forest floor, and carbon emissions to the atmosphere. Wildfires can also have long-term effect on carbon cycling. Indeed, when resilience of burned forests is exceeded, regenerating ecosystems differ from that of the pre-fire stands in composition, structure and biogeochemical cycles. Predictive models of carbon dynamics in boreal forests rarely take into account such multiple post-fire successional trajectories, which could bias long-term predictions of carbon storage and emissions. This study examined post-fire ecosystem resilience and carbon dynamics of black spruce-feathermoss forests of Quebec. Regeneration of black spruce, jack pine and understory plant communities were inventoried three to five years after the occurrence of a three-year episode of major fire activity in the province. In the studied plots, low burn severity had left intact a thick residual organic layer that impeded black spruce establishment. As a result, stem density of the burned stand decreased, while jack pine became dominant. Ericales resprouted abundantly. Understory regeneration was mainly driven by pre-fire site and stand characteristics. Post-fire understory composition reflected pre-fire species assemblages, because biological legacies were preserved by the low-severity fires. A model was used to simulate wildfires, succession and carbon dynamics at the landscape level, in order to assess the influence of forest floor combustion and post-fire tree regeneration on carbon cycling. After 500 years of simulation, modelling of burn severity and its influence on post-fire tree establishment caused a 13% decrease in predicted landscape carbon stocks. Simulation of the forest floor combustion alone caused a lower decrease in predicted carbon stocks than simulation of low spruce regeneration rates. Modelled fire emissions were mainly driven by variations in annual area burned. Loss of resilience of black spruce-feathermoss forests can have long-term consequences on carbon stocks. The model used in the present study could be further improved by incorporating explicit simulation of understory species dynamics.
vii
Table des matières
Résumé ___________________________________________________________________ iii
Abstract ___________________________________________________________________ v Table des matières _________________________________________________________ vii Liste des tableaux __________________________________________________________ ix Liste des figures ___________________________________________________________ xi Remerciements ___________________________________________________________ xiii Avant-propos _____________________________________________________________ xv Introduction générale ________________________________________________________ 1
I. Fonctionnement écologique de la forêt boréale fermée _________________________ 1 II. Caractéristiques et conséquences des incendies de forêt ________________________ 4 III. Résilience des pessières à mousses aux perturbations et aux changements climatiques 7 IV. Simulation de la dynamique des feux, de la végétation et du carbone à grandes échelle spatiale et temporelle ________________________________________________________ 12 V. Objectifs de la thèse ___________________________________________________ 14
Chapitre I ________________________________________________________________ 17 Three large fire years threaten resilience of closed crown black spruce forests in eastern Canada ___________________________________________________________________ 17 Abstract __________________________________________________________________ 18 Résumé __________________________________________________________________ 19 I. Introduction _________________________________________________________ 20 II. Methods ____________________________________________________________ 22 III. Results _____________________________________________________________ 33 IV. Discussion __________________________________________________________ 40 Acknowledgments __________________________________________________________ 47 Chapitre II _______________________________________________________________ 49 Ecological controls on post-fire vegetation assembly at multiple spatial scales ________ 49 Abstract __________________________________________________________________ 50 Résumé __________________________________________________________________ 51 I. Introduction _________________________________________________________ 52 II. Materials and methods. _________________________________________________ 54 III. Results _____________________________________________________________ 61 IV. Discussion __________________________________________________________ 71 Conclusion ________________________________________________________________ 74 Acknowledgments __________________________________________________________ 75
viii
Chapitre III ______________________________________________________________ 77 Microclimate, burn severity and post-fire tree establishment affect predictions of long-term carbon dynamics in a boreal forest landscape ______________________________ 77
Abstract __________________________________________________________________ 78 Résumé __________________________________________________________________ 79 I. Introduction _________________________________________________________ 80 II. Material and methods __________________________________________________ 83 III. Results ____________________________________________________________ 100 IV. Discussion _________________________________________________________ 111 Conclusion _______________________________________________________________ 117 Acknowledgments _________________________________________________________ 117
Conclusion générale _______________________________________________________ 118
I. Sévérité des feux dans la pessière à mousses du Québec ______________________ 118 II. Synthèse des principaux résultats de la thèse _______________________________ 119 III. Applications pour l’aménagement forestier ________________________________ 122 IV. Amélioration de la modélisation de la dynamique du carbone en forêt boréale _____ 123
Bibliographie _____________________________________________________________ 125 Annexes _________________________________________________________________ 141
Appendix A: Candidate models and results of model selection with AICc for conifer
regeneration analysis (Chapter 1) _____________________________________________ 141 Appendix B: Supplementary material for Chapter 2 _______________________________ 146 Appendix C: Species input parameters for Chapter 3 ______________________________ 155 Appendix D: Construction of the initial community map for Chapter 3 ________________ 160
ix
Liste des tableaux
Table 1: Location and characteristics of the sampling sites _____________________________ 26 Table 2: Acronyms and definitions of the variables ___________________________________ 31 Table 3: Controls on pre-fire organic layer depth, organic layer burned, and post-fire organic layers ______________________________________________________________________ 35 Table 4: Pre- and post-fire characteristics of plots according to topographic position ________ 39 Table 5: Species functional traits extracted from TOPIC for vascular plants _______________ 56 Table 6: Results of the three distinct IndVal analyses: A- indicator species, B- indicator
environmental variables and C- indicator functional traits of the three PAM site clusters. Only indicators with P < 0.05 are represented. ___________________________________________ 64 Table 7: Partition of variation in post-fire community functional traits (CWM) at three spatial scales, using partial RDA analysis on Hellinger-transformed species cover data. R2 adj: CWM variation explained by model; DF: degrees of freedom of model. E = matrix of environmental variables (forward selected to obtain a parsimonious model) ___________________________ 66 Table 8: A posteriori correlation (R2) between environmental variables and ordination space of
the first two axis of RDA of traits vs. spatial predictors _______________________________ 70 Table 9: Comparison of C fluxes modelled and measured for 7 years at the EOBS eddy-flux tower site. Calibration was conducted in the absence of fire. __________________________ 102 Table 10: Landscape C stocks and C fluxes predicted by LANDIS-II after 500 years of simulation under S1, S2, S3. ____________________________________________________________ 107
xi
Liste des figures
Figure 1: Schéma des principales voies de succession après feu dans les peuplements d’épinette noire et de pin gris. Adapté de Chabot et al, 2009._____________________________________ 6 Figure 2 : Organigramme représentant la problématique de la thèse et l’articulation des trois chapitres ____________________________________________________________________ 16 Figure 3: Map of the study area. Black spruce regeneration (A) and jack pine regeneration (B) after 2005, 2006 and 2007 fires in the BS-feathermoss bioclimatic domain of Quebec. Red, orange and green circles represent, respectively, fire sites with negative, null and positive CD values. Labels refer to fire ID numbers given by the fire protection agency of Quebec presented in Table 1. _____________________________________________________________________ 24 Figure 4: Location of the 14 fire sites and schematic representation of the nested experimental design.______________________________________________________________________ 27 Figure 5: Relative frequency of post-fire seedbeds in sampling plots and proportion of black spruce and jack pine seedlings established on different seedbed types. ____________________ 36 Figure 6: Differential effect of TsP (total amount of precipitation 3 summers after fire event) on black spruce regeneration in sites of varying ROM (residual organic matter thickness). Symbols represent real data, solid lines represent model predictions, and dashed lines represent 95% confidence intervals ___________________________________________________________ 37 Figure 7: Diagram of the different data matrices needed for the study and analyses performed with respect to the three research questions. RDA: Canonical Redundancy Analysis, PAM: Partition Around Medoids (i.e. cluster analysis), CWM: Community Weighted Means (i.e. plots x traits matrix), Z and F: matrix of spatial variables at inter-zone and inter-fire site scales (identity matrices); S1 & S2: matrix of spatial variables at intra-zone and intra-fire site scale (staggered matrix filled with Moran eigen vector maps). _______________________________________ 59 Figure 8: Results of constrained ordination (distance-based Redundancy Analysis) showing the relationship of understory species composition to environmental variables, 3 to 5 years after fire events. Different symbols represent sites belonging to different clusters defined by PAM analysis, based on differences in species composition among sites. Environmental variables are represented by black arrows. %Pine = proportion of Jack pine in pre-fire canopy, Pcp = summer precipitations, Temp = summer temperatures, COM = consumed organic matter, BurnInd = burn index, Age= pre-fire stand age, %Clay = mineral soil clay content, Slope = slope inclination, DamCnpy = damage to the canopy, BA= basal area. __________________________________ 63 Figure 9: Results of redundancy analysis showing relationship of trait values (community weighted means) to spatial predictors at regional (A) landscape (B) and local scales (C). Traits are represented by black arrows with the circle of equilibrium. See Table 5 for trait code definitions and more information on the measured traits. Only traits that significantly contribute to ordination are represented. Spatial predictors are represented by blue dots. _________________________ 68 Figure 10: Map of study area with the five ecoregions ________________________________ 85 Figure 11: Schematic diagram of the Century Succession extension for LANDIS-II (S1, adapted from Scheller et al. 2011) and new carbon pools and transfers added in the modified versions of
xii
the model (used under S2 and S3). New pools and C transfers added in the modified version of the model are represented in thick black arrows and boxes and bold text. Brackets identify modifications of the soil module (used under S2 and S3) and of the post-fire establishment module (used under S3) ________________________________________________________ 87 Figure 12: Schematic diagram representing the fate of C stored in soil organic matter during a fire event as a function of soil water content. ___________________________________________ 92 Figure 13: Comparison of LANDIS-II Century Succession extension (S2 version) monthly net ecosystem exchange (NEE, g C m-2 month-1) and measured NEE at the EOBS eddy flux tower
sites in the region of Chapais, Quebec. ___________________________________________ 101 Figure 14: Map of soil water content (cm) at five years of simulation under S1 (A) and S2 (B). __________________________________________________________________________ 103 Figure 15: Average fire C emissions per area burned (A) and mean annual fire C emissions at landscape scale (B) under the three scenarios. Values represent the average of 500 years of simulation, with 10 repetitions for each scenario. Error bars correspond to a 95% confidence interval. ____________________________________________________________________ 104 Figure 16: Distribution of fuel types accross the study area after 500 years of simulation and corresponding maps of time since last fire _________________________________________ 105 Figure 17: Comparison of aboveground live biomass (g C.m-2) of black spruce (A), jack pine (B),
aspen (C), and mosses (D), at year 500 of S1, S2 and S3 simulations. Bars represent the average of ten model replicates; vertical arrows correspond to a 95% confidence interval. __________ 108 Figure 18: Comparison of monthly mean NEE (g C m−2) averaged across the whole simulation
period among simulations of baseline, S2 and S3 scenarios. Bars represent the average of ten model replicates; vertical arrows correspond to a 95% confidence interval. _______________ 109 Figure 19: Predicted carbon fluxes and carbon storage for the five ecoregions under the S3 scenario. ___________________________________________________________________ 110
xiii
Remerciements
Je remercie tout d’abord le Conseil National de Recherche en Sciences Naturelles et en Génie d’avoir financé cette recherche; ainsi qu’Abitibi Consolidated de m’avoir accordé une bourse.
La partie de mon travail que j’ai le plus appréciée au cours de ce projet de doctorat et qui me donne envie de poursuivre le travail de chercheur, a été la rencontre avec un grand nombre de personnes avec des compétences et des savoir-faire très diversifiés. J’ai été surprise de voir à quel point ces personnes ont pris le temps de m’aider dans mes différents travaux et avaient à cœur de transmettre leurs connaissances.
Je remercie ma directrice de thèse Alison Munson de m’avoir offert la possibilité de travailler sur ce projet passionnant. Je lui suis très reconnaissante pour son encadrement et pour cette excellente expérience au sein de son laboratoire, j’ai appris énormément au cours de ces quatre années.
Je tiens à remercier Florent Mouillot et Isabelle Aubin, qui m’ont accueillie en stage dans leurs laboratoires respectifs à Montpellier en France et à Sault Ste Marie en Ontario. Dans les deux cas j’ai été extrêmement bien accueillie et j’ai beaucoup apprécié leur collaboration et leur apport sur les manuscrits par la suite.
Un grand merci à Serge Payette pour ses remarques très pertinentes, en particulier sur mon premier chapitre et pour ses encouragements.
Merci également à Steve Cumming, pour m’avoir initiée et intéressée à la modélisation de la dynamique des feux et à Eliot McIntire pour ses conseils sur le dispositif expérimental.
J’ai énormément appris en statistiques et en programmation grâce à Marc Mazerolle, qui a également toujours été disponible pour m’aider et répondre à mes questions au cours de mes analyses de données.
xiv
Merci à Laure Soucémarianadin et Sylvain Pelletier-Bergeron, qui ont participé à l’élaboration du dispositif expérimental et avec qui j’ai partagé mes étés de terrain. Je m’estime très chanceuse de les avoir eu comme coéquipiers et j’ai beaucoup apprécié autant leur façon de travailler que leurs personnalités (ce qui est excessivement important quand on passe plusieurs mois en forêt ensemble).
Merci à André Beaumont, pour son aide sur le terrain et pour nous avoir évité quelques déboires grâce à son sens de l’orientation exceptionnel. Merci beaucoup à Marie Coyea pour son aide logistique.
Merci aussi à tous les assistants de terrain et de laboratoire qui ont grandement contribué à rassembler les données qui ont été utilisées dans ce travail: Julie Venne, Mathieu Frégeau, Mathieu Blanchette, Jérôme Bérubé, Thomas Payer et Caroline Paradis.
Merci à Ann Delwaide pour son aide précieuse en dendrochronologie, à Rémi St-Amant qui a passé du temps à m’aider avec BioSIM et les données météorologiques, à Carole Coursolle et Hank Margolis qui m’ont permis d’accéder aux données du site EOBS et ont répondu à toutes mes questions.
Merci à l’équipe du professeur Robert Scheller pour la formation au modèle LANDIS-II et en particulier à Melissa Lucash et Brian Miranda pour leur assistance technique.
Je voudrais aussi remercier sincèrement les amis que j’ai rencontrés ici au Québec, notamment ceux qui ont transité par le pavillon Abitibi Price. Je ne veux pas citer de noms parce qu’il y en a beaucoup, par chance et j’aurais trop peur d’oublier quelqu’un, mais ils se reconnaitront et ils savent qu’ils vont me manquer quand je serai rentrée en France.
Merci aussi à ma sœur et à mes parents qui ont toujours été là pour m’aider m’encourager. Et évidemment, merci à Albin pour son aide, son soutien et sa patience légendaire en toutes circonstances.
Je voudrais terminer en dédiant ce travail à mon encadrante de maîtrise et amie, Sandra Patiðo, que j’appréciais beaucoup, comme tous les gens qui ont eu la chance de la côtoyer, qui malheureusement nous a quittés en août 2011
xv
Avant-propos
Insertion d’articles
La présente thèse est composée de trois chapitres rédigés en anglais sous forme d’articles scientifiques. Pour chaque chapitre, j’ai établi les objectifs de recherche et formulé les hypothèses, planifié les dispositifs expérimentaux et supervisé la collecte des données, puis réalisé les analyses statistiques et les simulations, l’interprétation des résultats et la rédaction des articles scientifiques.
Chapitre I- Boiffin, J., and A. D. Munson. 2013. Three large fire years threaten resilience of closed crown black spruce forests in eastern Canada. Ecosphere 4(5):56.
Chapitre II- Boiffin, J., Aubin, I., and A. D. Munson. Initial response of boreal understory vegetation to low severity fires at local to regional scales. L’article est soumis.
Chapitre III-Boiffin J., Mouillot F. and A. D. Munson. Microclimate, burn severity and post-fire tree establishment affect long-term predictions of carbon dynamics in a boreal forest landscape. L’article sera soumis sous peu.
Mon travail a été encadré par ma directrice de thèse, Alison Munson, qui est donc co-auteure de tous les chapitres de cette thèse. Isabelle Aubin a fourni une partie des données utilisées pour le deuxième chapitre, a encadré l’analyse statistique et la rédaction du manuscrit, elle est donc co-auteure du second chapitre. Finalement, Florent Mouillot a été un collaborateur important pour le troisième chapitre, puisqu’il a participé aux étapes de programmation du modèle et a apporté ses révisions au texte du chapitre. J'adresse donc mes sincères remerciements à ces co-auteurs.
1
Introduction générale
I. Fonctionnement écologique de la forêt boréale fermée
1. Importance géographique et écologique du biome
La forêt boréale est le biome continental qui renferme la plus grande quantité de carbone, soit approximativement 37 % des stocks la biosphère terrestre (Smith et al. 1993). Elle occupe donc une place centrale dans le cycle du carbone global et joue un rôle déterminant dans les mécanismes de rétroaction des écosystèmes terrestres face aux changements climatiques. Cela s’explique d’abord par son étendue de 14,7 million km2, soit 11% de la surface terrestre (Bonan and Shugart 1989). Au Québec, la forêt boréale fermée constitue le domaine bioclimatique de la pessière à mousses et représente à elle seule 28% de la province, soit 412 400 km2.
2. Composition et structure des forêts du domaine de la pessière à mousses
La plupart des peuplements qui composent la forêt fermée occupent des sites caractérisés par des sols podzoliques qui se sont développés sur des dépôts glaciaires ou fluvio glaciaires (Robitaille and Saucier 1998, Soil Classification Working Group 1998). L’essence forestière la plus répandue est l’épinette noire (Picea mariana (Mill.) B.S.P), qui forme généralement des peuplements purs dont le couvert est supérieur à 40% (Viereck 1983). On peut également la retrouver dans des peuplements mélangés avec d’autres conifères, notamment le pin gris (Pinus banksiana (Lamb.)) qui est la deuxième espèce la plus abondante du domaine et qui peut dominer la canopée sur les sites chauds, secs, peu fertiles, caractérisés par des dépôts grossiers et un drainage rapide (Cayford and McRae 1983). Le sapin baumier (Abies balsamea (L.) Mill) est très fréquent à l’est et au sud du domaine bioclimatique. Enfin le bouleau à papier, le peuplier faux tremble, l’épinette blanche (Picea glauca (Moench)) et le mélèze laricin (Larix laricina (Du Roi) K. Koch) sont également présents dans l’ensemble du domaine (Saucier et al. 2009). Le sous-bois est typiquement composé d’une épaisse couche de mousses hypnacées (Pleurozium schreberii) et/ou de sphaignes. Les arbustes de la famille des éricacées sont généralement abondants (Rhododendron groenlandicum, Kalmia angustifolia, Vaccinium spp.) (Larsen 1980, Saucier et al. 2009). Le sous-bois présente une diversité floristique
2
relativement faible mais joue un rôle déterminant dans le fonctionnement de l’écosystème, puisqu’il module la transmission de la lumière, peut influencer la germination, la croissance et la survie des plantules d’arbres et affecte grandement les cycles biochimiques (Oechel and Van Cleve 1986, Mallik 1987, Aubin et al. 2000, Nilsson and Wardle 2005).
3. Cycles du carbone et de l’azote
Les forêts boréales se situent dans des régions marquées par des températures très basses, avec des fluctuations saisonnières extrêmes et une saison de croissance courte (Larsen 1980, Bonan et al. 1990, Saucier et al. 2009). La faible vitesse de décomposition de la matière organique des sols est en grande partie responsable de l’importante séquestration du carbone en forêt boréale en général et dans la pessière à mousses en particulier. Dans les forêts d’épinette noire, les sols sont le plus souvent des podzols (Larsen 1980) et l’humus de type mor, peu aéré, présente un pH très acide, avec un ratio C:N particulièrement élevé (Bhatti and Apps 2000). Les mousses et les sphaignes prospèrent dans ces environnements où les horizons organiques du sol sont froids et humides (Van Cleve and Viereck 1981, Van Cleve and Yarie 1986, Bonan 2005). Elles forment un tapis épais et continu qui isole le sol et le maintient à des températures basses et à des taux d’humidité élevés, ce qui contribue fortement à ralentir l’activité biologique des micro-organismes et donc les processus de décomposition (Oechel and Van Cleve 1986, Bonan 2005). La matière organique peut alors s’accumuler et atteindre des épaisseurs très importantes (de 10 cm à 1 m). L’activité biologique réduite dans le sol provoque un faible taux de minéralisation et le blocage des nutriments, notamment de l’azote, qui reste sous une forme inutilisable par les plantes et par la plupart des micro-organismes (Van Cleve and Yarie 1986). La disponibilité de l’azote et des autres nutriments est un facteur limitant de la croissance des arbres qui est de fait très lente. Les mousses agissent comme un filtre qui piège une grande partie des nutriments apportés par la pluie, l’eau qui goutte des arbres, les retombées de poussières et par la litière qui se décompose à leur surface. La plupart des plantes vasculaires ne pourront disposer de ces nutriments qu’après la décomposition des mousses qui se produit à une vitesse très faible (Oechel and Van Cleve 1986).
3 Les éricacées, quant à elles, possèdent des rhizomes qui constituent un système racinaire étendu leur permettant de coloniser d’importantes surfaces de sol et d’y prélever l’eau et les nutriments du sol, exerçant ainsi une pression de compétition importante sur les arbres du couvert forestier (Mallik 1993, Inderjit and Mallik 1996). La litière composée de feuilles d’éricacées est riche en tannins qui peuvent former des liaisons stables avec divers composés azotés qui ne sont alors plus utilisables par les plantes et les micro-organismes (Inderjit and Mallik 1996, Bradley et al. 2000, Joanisse et al. 2009). Certaines éricacées produisent des substances inhibitrices de l’activité des communautés mycorhiziennes et des micro-organismes impliqués dans les cycles nutritifs du sol et le prélèvement des ressources par les plantes (Yamasaki et al. 1998). La présence d’un important couvert d’éricacées peut ainsi fortement nuire à la décomposition de la litière, à la formation de l’humus et ultimement, à la nutrition des arbres (Bradley et al. 2000). Les écosystèmes de forêt boréale sont soumis à de basses températures et sont situés à des latitudes nordiques, avec un faible angle incident des rayons solaires. Cet angle des radiations affecte grandement la productivité des arbres et renforce également l’influence de la situation topographique sur le déroulement des cycles biogéochimiques (Bonan and Shugart 1989). En effet, les variations des conditions microclimatiques et du drainage au sein d’un paysage vont entrainer des variations importantes des cycles du carbone et de l’azote qui sont fortement régulés par la température et l’humidité du sol (Bonan 2005, Milakovsky et al. 2012). Ainsi les sites occupant les dépressions et les cuvettes, ou encore les pentes exposées au Nord Est ont tendance à être plus froids et plus humides, ce qui limite la vitesse de décomposition de la matière organique, favorise son accumulation, et affecte la quantité de nutriments utilisables pour les plantes et les micro-organismes (Bonan and Shugart 1989).
En l’absence prolongée de perturbation, lorsque les forêts d’épinette noire vieillissent, la faible disponibilité des nutriments engendre une baisse de productivité du peuplement qui s’ouvre alors progressivement. Dans ces conditions, le retour à un peuplement productif ne peut survenir qu’après une perturbation telle qu’un feu sévère.
4
II. Caractéristiques et conséquences des incendies de forêt
1. Régime de feu dans le domaine bioclimatique de la pessière à mousses
Le feu de forêt est la principale perturbation naturelle des écosystèmes boréaux (Rowe and Scotter 1973, Wein and MacLean 1983). Le régime de feu d’une région donnée peut-être décrit par plusieurs attributs de la perturbation, parmi lesquels on retrouve notamment l’origine, l’intervalle de feu, la taille et la sévérité (Chabot et al. 2009). Dans le domaine bioclimatique de la pessière à mousses, la plupart des incendies sont déclenchés par la foudre. L’intervalle de feu est le temps moyen qui s’écoule entre deux incendies successifs dans une région donnée. Au Québec, il subit de fortes variations régionales, notamment liées à un gradient de précipitations qui sont plus abondantes à l’est qu’à l’ouest (Lefort et al. 2004, Chabot et al. 2009). L’intervalle de feu moyen varie de 100 ans dans la région du nord de Chibougamau située à l’est de la Baie James à près de 500 ans dans la région de la Côte Nord (Lesieur et al. 2002, Chabot et al. 2009). Le régime de feu en forêt boréale est caractérisé par des surfaces annuelles brûlées extrêmement variables et l’occurrence cyclique de grandes années de feux, au cours desquelles un nombre important de grands incendies surviennent à la faveur de longues périodes de conditions chaudes et sèches (Flannigan and Harrington 1988, Murphy et al. 2000, Stocks et al. 2003).
La sévérité des feux est l’impact du feu sur la végétation et le sol forestier (Brown and Debyle 1987, Lentile et al. 2006). Elle peut être extrêmement variable, aussi bien à l’échelle du paysage qu’à celle du peuplement. Elle dépend en effet de l’humidité des combustibles qui est elle-même fonction du type de végétation, des conditions météorologiques au moment du passage de l’incendie, mais également de la situation topographique du site brûlé (Kane et al. 2007, Turetsky et al. 2011). Ainsi, les dépressions mal drainées et les sites peu exposés au soleil sont potentiellement plus humides et donc susceptibles de brûler moins sévèrement. D’après une étude menée en Alaska, entre 10 et 90 % du sol forestier serait consumé lors d’un incendie (Kasischke et al. 2000a, Levine and Cofer 2000). On peut s’attendre à ce que le régime de feu de la pessière à mousses du Québec pour la période récente soit marqué par une sévérité
5 relativement faible au niveau du sol forestier (Lecomte et al. 2006, Girard et al. 2009). En effet l’intervalle de feu est plus long et les niveaux de précipitations sont plus élevés qu’en Alaska (Chabot et al. 2009, Barrett et al. 2011). De surcroit, plus de 50 % des feux surviennent au printemps (Ministère des Ressources Naturelles du Québec, MRN 2010), juste après ou pendant la fonte des neiges, lorsque les horizons du sol sont particulièrement humides (Heinselman 1981).
2. Conséquences des incendies aux échelles du paysage et du peuplement
Les incendies de forêt boréale sont un élément clé de la dynamique des peuplements et de la structure des paysages en forêt boréale (Rowe and Scotter 1973, Van Wagner 1978, Heinselman 1981, Kasischke and Johnstone 2005, Lecomte et al. 2005, Johnstone et al. 2010b). Les variations spatiales des régimes de feu sont très importantes et introduisent de l’hétérogénéité dans la mosaïque de végétation qui constitue les paysages de forêt boréale. Chaque compartiment géographique de la mosaïque est caractérisé par le temps écoulé depuis le dernier feu, dont dépendent la structure et le stade de succession atteint par les peuplements (Van Wagner 1978, Lecomte 2005). Les paysages marqués par différents régimes de feu comportent ainsi des proportions différentes de vieux peuplements inéquiens et de peuplements jeunes dominés par les essences pionnières (Rowe and Scotter 1973, Van Wagner 1978, Chabot et al. 2009).
À l’échelle du peuplement, les incendies se traduisent généralement par la mort d’une importante proportion des arbres de la canopée et par la destruction de la végétation du sous-bois et d’une partie de la matière organique du sol forestier. Dans la pessière à mousses, les feux sévères produisent des conditions propices à l’établissement d’un peuplement productif (Nguyen-Xuan et al. 2000, Harper et al. 2005, Pollock and Payette 2010). Le sol minéral exposé lors d’une destruction complète de la matière organique et le bois mort constituent en effet de bons lits de germination pour les conifères comme l’épinette noire et le pin gris (Charron and Greene 2002).
La majorité des semis d’épinette noire et de pin gris s’établissent dans les trois premières années qui suivent le passage du feu (St-Pierre et al. 1992, Charron and Greene 2002). Dans la pessière à mousses, la trajectoire successionnelle la plus fréquente après un feu sévère est le remplacement du peuplement d’origine par un peuplement de composition et
6
de densités similaires (Viereck 1983, Figure 1). L’épinette noire étant semi-tolérante à l’ombre, elle peut s’établir directement après l’incendie, parfois de façon concomitante avec des espèces pionnières comme le pin gris et le peuplier. Ces deux espèces sont caractérisées par une croissance rapide, une moindre longévité et une intolérance à l’ombre. Ainsi l’épinette noire peut croître sous leur couvert à un rythme plus lent; en l’absence prolongée d’un feu, elle remplace progressivement le peuplier ou le pin dans le couvert du peuplement lorsque ceux-ci commencent à mourir (respectivement au bout de 120 et 150 ans, Harper et al. 2005, Chabot et al. 2009, Figure 1). Elle pourra ensuite se régénérer par marcottage jusqu’à ce qu’un nouvel incendie ou qu’une autre perturbation ne viennent ré-initier la succession (Pollock and Payette 2010).
Figure 1: Schéma des principales voies de succession après feu dans les peuplements d’épinette noire et de pin gris. Adapté de Chabot et al, 2009.
7
III. Résilience des pessières à mousses aux perturbations et aux changements climatiques
1. Définition de la résilience
La résilience d’un écosystème peut être définie comme sa capacité à subir une perturbation ou une modification progressive de son environnement tout en maintenant ses propriétés structurelles et fonctionnelles. On la caractérise parfois comme le temps nécessaire à l’écosystème pour retrouver ses principaux attributs (espèces dominantes, densité et structure du couvert, communautés du sous-bois) après une perturbation (Folke et al. 2004, Walker et al. 2004, Johnstone et al. 2010b). La résilience des forêts québécoises est une préoccupation majeure qui a conduit le Ministère des Ressources Naturelles à mettre en vigueur la Loi sur l'aménagement durable du territoire forestier en avril 2013. Cette approche d’aménagement vise à maintenir des écosystèmes sains et résilients en misant sur une diminution des écarts entre les paysages naturels et aménagés (Gauthier et al. 2009). Une des voies pour y parvenir est de s’inspirer des perturbations naturelles et notamment des incendies, lors de l’aménagement forestier. Il s’agit premièrement de rapprocher les propriétés des coupes des régimes de feu de forêt ; deuxièmement d’exploiter la ressource de façon responsable pour ne pas l’épuiser, en tenant compte du fait que les forêts sont également affectées par des perturbations naturelles auxquelles elles sont plus ou moins résilientes (Gauthier et al. 2009). Aussi, il est important de comprendre les mécanismes permettant la régénération des forêts d’épinette noire face aux incendies, de même que les conditions susceptibles d’entrainer des pertes de résilience.
2. Mécanismes permettant la résilience des écosystèmes
Les forêts du domaine de la pessière à mousses se maintiennent dans des milieux soumis aux incendies répétés à travers de longues périodes (plusieurs centaines à plusieurs milliers d’années) au cours desquelles surviennent des modifications significatives du climat (Prentice and Bartlein 1991, Masek 2001, Pollock and Payette 2010). Cela implique l’existence de mécanismes assurant la résilience des communautés, d’une part aux changements climatiques, d’autre part aux incendies de forêt. La résilience aux modifications du climat peut d’abord s’expliquer par l’amplitude de la niche climatique
8
des espèces de forêt boréale (Bonan et al. 1990, Camill and Clark 2000, Chapin et al. 2004). À l’intérieur de leurs aires de répartition, les arbres matures possèdent un réseau de capture des ressources complexe et étendu qui leur permet de faire face à des niveaux important de stress environnemental (Chapin et al. 2004, Johnstone et al. 2010c). Enfin, il existe de nombreuses rétroactions par lesquelles les organismes d’un écosystème influencent leur milieu de vie et cela tend à limiter les possibilités d’invasion par des espèces non-indigènes (Camill and Clark 2000, Johnstone et al. 2010a).
Les espèces qui constituent les communautés des pessières à mousses sont dotées d’adaptations leur permettant de perdurer dans des milieux où les feux sont récurrents (Rowe 1983, Allen 2008). L’ensemble de ces stratégies d’adaptation constitue les traits de réponse au feu et concerne la plupart du temps les modes de reproduction et de colonisation des sites brûlés (Rowe 1983, Schimmel and Granström 1996, Lavorel and Garnier 2002). Sur la base de ces traits fonctionnels de réponse, on peut classer les espèces de la pessière à mousses en plusieurs catégories ou groupes fonctionnels, qui ne sont pas mutuellement exclusifs (Schimmel and Granström 1996, Pausas et al. 2004, Allen 2008):
Les espèces formant des banques de graines aériennes. L’épinette noire et le pin gris possèdent des cônes sérotineux. Ces cônes sont enduits d’une résine qui les protège au cours des feux de cime et fond lentement sous l’effet d’une chaleur intense. Les cônes s’ouvrent lorsque la résine a fondu et les graines sont libérées après l’épisode de feu.
Les espèces capables de se régénérer par multiplication végétative grâce à des organes souterrains (rhizomes, drageons) qui peuvent survivre au passage d’un incendie modérément sévère.
Les espèces formant des banques de graines souterraines, grâce à une longue durée de vie des graines dans le sol.
Les espèces qui produisent des graines de petite taille, dont la dispersion peut s’étendre sur de longues distances. Les sites sévèrement brûlés présentent d’importantes proportions de sol minéral exposé, qui constitue un bon lit de germination pour ces petites graines, dispersées à partir de plantes vivantes dans le voisinage du brûlis. Le
9 peuplier faux-tremble et l’épilobe font partie de ce groupe fonctionnel, bien que ces deux espèces soient également capables de se régénérer par multiplication végétative.
Ainsi, quel que soit le cortège d’espèces présentes dans la communauté, la résilience de l’écosystème repose en grande partie sur l’existence de legs biologiques hérité du peuplement d’origine, incluant les arbres morts, les banques de graines ou de bourgeons, ou encore les îlots de végétation ayant survécu à l’incendie. L’abondance de ces legs biologiques dépend de la sévérité de l’incendie (Flinn and Wein 1977, Schimmel and Granström 1996).
L’intervalle de feu peut jouer un rôle important dans la composition des communautés végétales qui se régénèrent après feu, et sélectionner les espèces en fonction de leur stratégie de reproduction et de colonisation des milieux brûlés. Ainsi, d’après Pausas et al. (2004), les espèces capables de se régénérer par multiplication végétative seraient les plus flexibles vis-à-vis de la récurrence des incendies, tandis que les espèces possédant des cônes sérotineux et celles formant des banques de graines dans le sol seraient plus présentes dans les milieux où l’intervalle de feu est respectivement intermédiaire et long. Noble and Slatyer (1980) soulignent également l’importance de trois catégories de traits fonctionnels ou « attributs vitaux » des plantes qui, étant filtrés par la récurrence et la sévérité des feux, conditionnent l’assemblage des communautés au cours de la succession post-incendie, et le degré de résilience du peuplement brûlé :
Les traits qui déterminent l’arrivée d’une espèce sur un site après le passage du feu, ou la persistance de propagules sur le site pendant le feu,
Les traits qui déterminent la capacité à s’établir et à atteindre la maturité dans la communauté qui se met en place,
Les traits qui déterminent le temps nécessaire pour atteindre certains stades critiques du cycle de vie.
3. Causes potentielles de perte de résilience des pessières fermées
Malgré les mécanismes de résilience qui favorisent l’inertie des communautés sur plusieurs cycles de perturbation, des changements importants de type de végétation ont été observés suite au feu dans les forêts d’épinette noire. Dans la région de l’écotone
10
forêt-toundra, les échecs de régénération de l’épinette noire suite au passage des feux semblent être à l’origine de la régression de la forêt au profit de landes arbustives caractéristiques de la toundra (Asselin and Payette 2005). À proximité des limites latitudinales ou altitudinales des arbres, une régénération déficiente de l’épinette noire après des incendies s’est traduite par un changement de dominance vers le peuplier faux tremble en Alaska (Johnstone et al. 2010b) et vers le pin gris dans le nord du Québec (Lavoie and Sirois 1998). Des intervalles de feu plus courts (<50 ans) que le laps de temps nécessaire à l’épinette noire pour atteindre l’âge optimal de production de graines ont été identifiés comme étant une cause importante de perte de résilience des peuplements d’épinette noire (Lavoie and Sirois 1998, Johnstone et al. 2009). À la limite de l’aire de répartition de l’épinette noire, les conditions climatiques en interaction avec les incendies peuvent également être à l’origine des faibles niveaux de régénération observés (Sirois and Payette 1989, 1991, Meunier et al. 2007). En effet, la capacité reproductive des arbres peut y être diminuée à cause du faible nombre de degrés-jour accumulés annuellement, ce qui a pour effet d’espacer les bonnes années semencières dans le temps.
Plus au sud, à l’intérieur de la forêt fermée, les écosystèmes semblent plus résilients, même si des changements de trajectoire de succession ont également été observés. En effet, Girard et al. (2008) ont démontré que près de 9 % des forêts fermées étaient devenues des pessières à lichens ouvertes au cours des 50 dernières années. Le changement s’était effectué après feu dans la plupart des cas. Les perturbations en rafale, telles qu’une épidémie de tordeuse rapidement suivie d’un feu, ou encore les incendies peu sévères peuvent provoquer les échecs de régénération de l’épinette noire et la formation d’une pessière ouverte à lichens (Jasinski and Payette 2005, Girard et al. 2009). Parmi les autres causes invoquées pour expliquer les pertes de résilience et les changements de végétation, les conditions météorologiques au moment de l’incendie et dans la phase initiale de régénération pourraient jouer un rôle important (Thomas and Wein 1985, Johnstone et al. 2010c). En effet les plantules qui s’établissent juste après une perturbation sont plus vulnérables au stress hydrique que les arbres adultes qui possèdent un réseau de capture des ressources plus étendu.
11 4. Création d’états alternatifs stables
Les changements de végétation observés suite à une régénération déficiente de l’épinette noire ne sont pas nécessairement réversibles à moyen ou à long terme. Lorsque le nouvel écosystème peut se maintenir à long terme dans les mêmes conditions climatiques, topographiques et édaphiques que l’écosystème d’origine, tout en étant soumis aux mêmes régimes de perturbation, on considère qu’il constitue un état alternatif stable (Sutherland 1974). Les pessières ouvertes sont un exemple d’état alternatif stable des pessières à mousses ; elles peuvent en effet se régénérer suite à un incendie et coexister avec des pessières à mousses fermées sur des sites aux conditions très similaires (Jasinski and Payette 2005). Les peuplements envahis par les éricacées peuvent demeurer sous forme de landes improductives car ces plantes nuisent fortement à l’établissement et à la croissance des semis de conifères (Mallik 1995). Dans le cas où le nouvel écosystème qui se met en place est un peuplement de pin gris, la probabilité de ré-invasion du couvert forestier par l’épinette noire au cours de la succession est relativement faible. En effet, l’intervalle de feu est inférieur à la longévité du pin gris (150 ans) dans une grande partie de la forêt boréale et les peuplements de pin gris présentent une inflammabilité élevée. Aussi il est probable qu’un nouvel incendie survienne et permette au pin de se régénérer, avant que la sénescence des peuplements ne fournisse des opportunités pour l’invasion de l’épinette noire sur le site (Lavoie and Sirois 1998).
Plusieurs aspects du fonctionnement des écosystèmes peuvent être affectés lorsqu’une perte de résilience se produit et conduit au remplacement du type de végétation dominant par un état alternatif stable. Un changement de l’espèce dominant le couvert influence les cycles du carbone et de l’azote à la fois directement et indirectement. La productivité de l’essence dominante, la quantité de lumière transmise au sol et de la composition chimique de la litière vont affecter l’activité des microorganismes et la vitesse de décomposition de la matière organique (Gower et al. 1997, Preston et al. 2000, Miyanishi and Johnson 2002, Yu et al. 2002). Gower et al. (1997), dans une étude menée sur des peuplements de forêt boréale au Manitoba et en Saskatchewan, ont montré que le stockage du carbone dans le sol, la biomasse aérienne et les mousses était jusqu’à sept fois plus important dans les forêts d’épinettes noire que dans les forêts de pin gris.
12
IV. Simulation de la dynamique des feux, de la végétation et du carbone à grandes échelle spatiale et temporelle
1. Observation de la résilience à différentes échelles spatiales et temporelles
En forêt boréale, les feux de forêt peuvent atteindre des superficies de plusieurs centaines de milliers d’hectares et au Québec, la longévité des arbres et l’intervalle de feu dépassent en général 100 ans. Il en découle que la résilience des forêts est une notion relative qui dépend de l’échelle spatiale et temporelle considérée. Ainsi, une perte de résilience observée à l’échelle d’un seul peuplement peut se révéler non significative lorsqu’on se place à l’échelle du paysage ou à l’échelle de la région (Chapin et al. 2004). De même, l’appréciation de la résilience d’un écosystème à une perturbation peut varier en fonction du temps écoulé depuis cette perturbation, puisqu’il est impossible d’affirmer qu’un changement du type d’écosystème faisant suite à une perturbation est définitif en l’absence d’observations à long-terme prouvant la stabilité du nouveau type de végétation sur plusieurs cycles de succession (Carleton 1982, Lesieur et al. 2002, Lloret et al. 2012). Pour pouvoir faire des prédictions réalistes des rétroactions exercées par la forêt boréale sur les changements climatiques, il apparaît nécessaire d’évaluer la résilience des écosystèmes aux incendies ainsi que les conséquences de changements de trajectoire de succession à l’échelle du paysage, et ce, sur plusieurs centaines d’années.
Pour des raisons pratiques d’investissement en temps et en moyens, les études empiriques portant sur la réponse des communautés végétales aux perturbations sont souvent été menées à l’échelle du peuplement. Cependant, l’assemblage des communautés végétales est le résultat de l’action de différents processus environnementaux (climat, disponibilité des ressources, perturbations) et biotiques (dispersion, hasards de la survie de la mortalité, et interactions entre les espèces, (MacArthur and Levins 1967, Díaz et al. 1998, Hooper et al. 2002, Grime 2006) qui interviennent à différentes échelles spatiales. Ainsi, on considère que les processus biotiques exercent un contrôle à l’échelle locale, tandis que les gradients climatiques, au contraire, filtrent les assemblages d’espèces à l’échelle régionale (Díaz et al. 1998, Hooper et al. 2002, Chalmandrier et al. 2013). Pour être en mesure de prédire la résilience aux perturbations à de multiples échelles spatiales, il est donc nécessaire de mieux comprendre quelle est la contribution de ces différents
13 processus biotiques et abiotiques dans le contrôle de l’assemblage des communautés après feu. Plus précisément, il est nécessaire de mieux connaître la façon dont les traits fonctionnels des espèces sont filtrés aux échelles locales, du paysage et de la région (Roberts and Gilliam 1995, Azeria et al. 2011, Swenson et al. 2012).
2. Modélisation de la dynamique de la végétation, du feu et du carbone
Les modèles permettant de simuler la dynamique de la végétation, du feu et/ou des cycles biogéochimiques peuvent être un outil de prédiction très efficace lorsque le recours aux études purement empiriques est compromis par l’importance des superficies et des échelles de temps considérées (Keane et al. 2004, Mouillot et al. 2014).
Il existe deux grandes catégories parmi les modèles permettant de simuler la dynamique des écosystèmes (Liu et al. 2011). Premièrement, les modèles « à compartiments » représentent l’écosystème sous forme de différents réservoirs de biomasse et de carbone (feuilles, branches, débris ligneux, matière organique du sol etc.) et ne considèrent généralement pas la structure et la composition de la végétation de manière explicite : citons par exemple CENTURY (Parton et al. 1987), CASA (Potter 2004), Biome-BGC (Bond-Lamberty et al. 2005) et CBM-CFS3 (Kurz et al. 2009). Ils permettent l’étude des processus écophysiologiques et des cycles biogéochimiques et sont le plus souvent employés pour étudier l’impact des changements climatiques ou de l’enrichissement de l’atmosphère en CO2 sur les variations de croissance de la végétation et de la respiration
de l’écosystème. Deuxièmement, les modèles dits « démographiques » sont employés pour étudier l’impact des perturbations telles que les incendies sur la composition et la structure des forêts (Keane et al. 2004, Liu et al. 2011). Ils simulent de façon explicite la composition, la mortalité, la reproduction et la structure des peuplements. Des exemples de ce type de modèles sont : SEM-LAND (Li 2000), EMBYR (Hargrove et al. 2000) LANDIS-II (Scheller et al. 2007). Barrett (2001) propose également une comparaison de quatre de ces modèles (FETM, LANDSUM, SIMPPLLE, et VDDT). Dans cette catégorie de modèles, certains sont spatialement explicites, ce qui permet de prendre en compte des phénomènes tels que la dispersion des graines et les incendies, qui sont susceptibles de varier à l’intérieur d’un paysage et de générer de l’hétérogénéité spatiale dans la composition, l’âge et la structure des forêts.
14
Plus récemment, des modèles couplant l’étude de la dynamique des perturbations, de la succession et des cycles biogéochimiques à l’échelle du paysage ou du biome ont été développés (Keane et al. 2011, Scheller et al. 2011, Yue et al. 2013). Ces modèles offrent de nouvelles perspectives quant à la représentation explicite et complète des effets directs et indirects du feu sur les stocks et les flux de carbone. En ce qui concerne les effets directs du feu, la simulation de la combustion de la matière organique du sol forestier reste une lacune de ces modèles. Ces modèles pourraient encore être largement améliorés en représentant de façon plus mécaniste la sévérité du feu au niveau du sol, ses variations saisonnières et l’influence de la topographie, du microclimat et du type de végétation (Keane et al. 2004, Liu et al. 2011, Kasischke et al. 2013). Par ailleurs, la production de composés pyrogéniques (charbons) très stables et susceptibles de modifier les propriétés chimiques du sol est l’un des effets du feu qui est rarement représenté dans les modèles. Enfin, l’inclusion de différentes trajectoires de succession dans les prédictions des stocks et flux de carbone à long terme est une autre voie d’amélioration majeure de ces modèles (Conard and Solomon 2008, Turner 2010, Loehman et al. 2013)(Conard and Solomon 2008, Turner 2010, Loehman et al. 2013).
V. Objectifs de la thèse
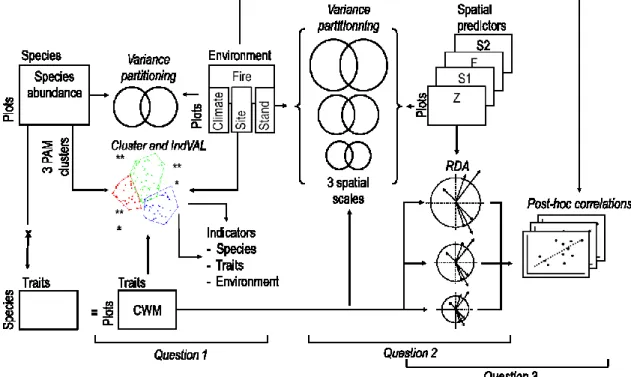
Cette thèse combine des observations réalisées sur le terrain et l’utilisation du modèle de simulation LANDIS-II pour évaluer l’impact de la sévérité des feux sur la résilience des pessières à mousses, sur la composition et l’organisation spatiale des communautés végétales qui se régénèrent après feu et sur le cycle du carbone (Figure 2).
Les études des deux premiers volets ont été basées sur l’observation de sites brûlés 3 à 5 ans plus tôt, en 2005, 2006 et 2007. Au cours de ces trois années, qui ont été marquées par des conditions chaudes et sèches, près de 1,2 million d’hectares de forêt ont brûlé dans la zone de protection intensive au Québec (zone située au sud du 51ème parallèle). De grands incendies ont touché l’ensemble du domaine bioclimatique de la pessière à mousses, ce qui représentait une opportunité d’étudier la régénération initiale après feu dans un grand nombre de sites répartis sur un gradient longitudinal de plus de 600 km, incluant des régions où les intervalles de feux sont longs.
15 Le premier chapitre de l’étude avait pour objectif d’évaluer la résilience des forêts d’épinette noire (Figure 2). Nous nous sommes plus particulièrement intéressés à la transition d’un peuplement dominé par l’épinette noire vers un peuplement dominé par le pin gris. Un tel changement de dominance a précédemment été observé suite à des incendies dans le domaine bioclimatique de la pessière à lichens (taïga) (Lavoie and Sirois 1998). Nous faisions l’hypothèse que cette transition vers le pin pouvait également se produire plus au sud, dans la forêt fermée, sous l’effet du régime de feu et des conditions météorologiques qui ont caractérisé les années 2005 à 2007. La sélection de modèle nous a permis d’estimer l’importance relative de la pluie de graines, de la qualité des lits de semences et des conditions météorologiques et microclimatiques sur le succès de la régénération du pin gris et de l’épinette noire.
Le même dispositif expérimental a été utilisé pour le deuxième volet qui portait sur l’étude de la régénération du sous-bois après l’incendie (Figure 2). Notre objectif était de comprendre l’organisation spatiale des patrons de végétation à l’échelle de la région, du paysage et du site brûlé et de déterminer quels facteurs environnementaux produisaient ces patrons. À chaque échelle spatiale, nous avons étudié la répartition des espèces et de leurs traits fonctionnels dans les différents sites, en mettant en relation les variations de la composition des communautés et la sévérité du feu, les caractéristiques du site et celles du peuplement brûlé.
Dans le troisième chapitre, nous avons utilisé LANDIS-II (Scheller et al. 2007), un modèle spatialement explicite permettant de simuler la dynamique des feux, l’établissement des cohortes végétales, la dynamique de la biomasse et les cycles du carbone et de l’azote dans le sol, afin de quantifier les effets directs et indirects du feu sur le stockage et les émissions de carbone à l’échelle du paysage et sur une période de 500 ans. Plus précisément, les effets directs considérés consistaient en la combustion de la biomasse aérienne et du sol forestier ainsi qu’en la production de charbon, tandis que les effets indirects étaient les changements de dominance de l’épinette noire vers le pin qui ont été observés au chapitre 1. Le but de ce chapitre était de démontrer la nécessité d’une meilleure représentation de la sévérité du feu dans les prédictions de stockage et d’émissions de carbone, ainsi que de fournir un outil de modélisation qui représente de
16
façon plus précise les effets du feu dans les pessières à mousses de l’est du Québec. Pour cela nous avons apporté des modifications à la façon dont le modèle calcule la température et l’humidité du sol, ajouté la destruction d’une partie de la matière organique et la formation d’un réservoir de charbon et modifié les règles d’établissement des espèces après feu pour qu’elles reflètent mieux les patrons de régénération étudiés dans les premiers chapitres.
17
Chapitre I
Three large fire years threaten resilience of closed crown black spruce
forests in eastern Canada
1
1 Version intégrale d’un article publié : Boiffin, J., and A. D. Munson. 2013. Three large fire years threaten
18
Abstract
An emerging paradigm regarding vegetation response to climate warming is that the interaction of weather extremes and disturbance will trigger abrupt changes in ecosystem types by overcoming resilience of dominant species. Black spruce (Picea mariana (Mill.) B.S.P.) ecosystems are widespread across the North American boreal forest, because of ecophysiological adaptations that allowed these communities to thrive in fire-prone areas. We investigated resilience of spruce-moss forests to weather-disturbance interaction after a 3-year period (2005 to 2007) of major fire activity caused by extreme fire weather in eastern Canada. Pre- and post-fire conifer densities and environmental parameters related to seed rain, post-fire seedbeds, microclimate, and post-fire weather were measured in 133 burned stands throughout the closed-crown forest of Quebec. Critically low black spruce (BS) regeneration was observed in almost all of the stands, leading to a decrease in stand density and a shift of species dominance from BS to jack pine (Pinus banksiana (Lamb.), JP). The studied sites were characterized by thick residual organic matter, resulting in a predominance of charred duff, a seedbed associated with low water retention and high variation in temperature. While high levels of JP seedling establishment were reported on this seedbed, it was unfavorable to BS germination and survival in the context of warm and dry weather that prevailed in post-fire summers. In these ecosystems, early vegetation establishment patterns are generally reliable predictors of future stand composition and the exclusion of BS will presumably be maintained through succession. During large fire years, a particularly fire-conducive weather leads to the occurrence of large fires that cause annual area burned to peak. Such extended periods of warm and dry conditions are also susceptible to impair BS recruitment and to cause broad-scale loss of resilience in the large areas of spruce forests affected by fire. Specific attention should therefore be paid to the prediction of the timing and frequency of large fire years in the future, since they represent time-windows with high probabilities of broad-scale vegetation shifts in this biome.
Keywords
Burn severity; climate-disturbance interaction; jack pine; Picea mariana; Pinus
19
Résumé
L’interaction des perturbations et de conditions météorologiques extrêmes est une cause reconnue de perte de résilience des écosystèmes. Nous avons étudié la résilience des pessières à mousses après trois années durant lesquelles le Québec a été marqué par des conditions sèches et une forte activité des feux. Les densités de conifères ont été inventoriées trois à cinq ans après incendie dans 133 parcelles. La régénération de l’épinette noire était déficiente dans 84% des parcelles, provoquant l’ouverture du peuplement et un changement de dominance vers le pin gris. Si l’établissement du pin était relativement élevé sur l’épaisse couche d’humus noirci qui recouvrait les parcelles, ce substrat, combiné à des étés secs après l’incendie, avait compromis la régénération de l’épinette. La probabilité d’observer des pertes de résilience des pessières noire à grande échelle augmente durant les grandes années de feu, lorsque d’importantes surfaces sont soumises aux incendies et aux conditions sèches.
Mots clés
Sévérité; interaction climat-perturbation ; pin gris ; Picea mariana ; Pinus banksiana ; régénération après feu ; Québec; résilience; états alternatifs stables.
20
I. Introduction
Vegetation response to climate change has provoked numerous hypotheses and requires close monitoring in the next decades (Johnstone et al. 2010b, Walther 2010). One hypothesis suggests that vegetation will respond gradually to climate change and that species will migrate northward as temperature increases (Davis and Botkin 1985, Overpeck et al. 1991, Parmesan and Yohe 2003). Palynological data provide evidence to support this paradigm for North American vegetation dynamics during the last 18 000 years (Prentice and Bartlein 1991). The current distribution of vegetation in boreal biomes is also fundamentally tied to climate (Woodward and Williams 1987). However, at a time scale relevant to policy and forest management (decades to millennia, Chapin et al., 2004), and within species ranges, these ecosystems have shown to be resilient to climate change (Chapin et al. 2004, Pollock and Payette 2010, Lloret et al. 2012). This resilience is attributable to species longevity, ability to reproduce vegetatively and biological legacies that reduce opportunities for regeneration of alternate species (Chapin et al. 2004, Barrett et al. 2011). An emerging hypothesis is that climate extremes could provoke faster and more ample ecosystem responses than mean trends (Gutschick and BassiriRad 2003, Jentsch et al. 2007). Extremes in climate may alter disturbance frequency, size and severity which could cause abrupt changes in vegetation if resilience thresholds of the dominant species are exceeded (Scheffer and Carpenter 2003, Chapin et al. 2004). In the boreal forest of North America, fire activity fluctuates greatly from year to year, with episodic major fire years characterized by a long fire season and numerous large fires (Stocks et al. 2003, Kasischke and Turetsky 2006, Kasischke et al. 2010). Large fire years are related to extreme fire weather conditions, more specifically to extended periods with low daily precipitation, low relative humidity and high temperatures (Flannigan and Harrington 1988). When these warm and dry conditions persist, they can also affect post-disturbance regeneration. The few-year time window during which dominant species establish after fire is a critical phase for future stand development (Sirois and Payette 1989). Even if mature trees may cope with environmental stress because of an established network of resource capture, the cohort
21 that is germinating in the post-disturbance phase may be more vulnerable (Folke et al. 2004, Johnstone et al. 2010c). Interactions of climate and disturbance are thus likely to provoke the most important and long lasting effects on ecosystem structure and function (Jentsch and Beierkuhnlein 2003, Chapin et al. 2004). In the region of the forest-tundra border in eastern Canada, tree depletion in spruce stands after fire lead to expansion of tundra inside of the northerly boreal forest (Sirois and Payette 1991) or to major shifts in species dominance from black spruce (Picea mariana (Mill.) B.S.P, abbreviated BS) to jack pine (Pinus banksiana (Lamb.), abbreviated JP) (Lavoie and Sirois 1998). Forests in this ecotone of high latitudes are particularly responsive to climate-disturbance interaction, because of the decreased reproductive capacity of trees exposed to cold temperatures (Sirois and Payette 1991, Sirois 2000). Further south, in the commercial forest, the closed spruce-moss stands are expected to be more resilient (Sirois and Payette 1991) with fire initiating direct replacement by the same species assemblage (Black and Bliss 1978, Viereck 1983). However, Girard et al. (2008, 2009) revealed that, over the last 50 years, 9% of the area occupied by closed spruce-moss forests in Quebec has converted to open spruce-lichen woodlands after low severity fires or repeated disturbances. If fire can lead to important declines in spruce density within the BS feathermoss bioclimatic domain, then shifts in species dominance from BS to JP may occur in sites where spruce fails to regenerate. JP, like BS, possesses serotinous cones that allow the species to thrive in fire prone areas (Cayford and McRae 1983). It is a shade-intolerant moderately long-lived species that reaches an optimum in seed production earlier than BS (Cayford and McRae 1983). JP is better adapted to areas where the fire cycle is short and pine stands are eventually replaced in the extended absence of fire (Cayford and McRae 1983, LeGoff and Sirois 2004) while BS is a shade tolerant long-lived species that can reproduce vegetatively by layering 50 to 70 years after stand initiation (Black and Bliss 1980, Viereck 1983). Fire return interval is thought to play a major role in the respective spatial abundance of the two species in the landscape (LeGoff and Sirois 2004). Lavoie and Sirois (1998) hypothesized that fire severity and post-fire weather may be two additional drivers of the shift in species dominance, while Johnstone et al. (2010 a) found that post-fire establishment of BS was higher in moist sites characterized by low fire severity in Alaska.