ROLE DES GENES HOMOLOGUES A terO DANS LE CYCLE VITAL DE STREPTOAdYCES COELICOLOR
Par Edith Sanssouci
thèse présentée au Département de biologie en vue de l'obtention du grade de docteur es sciences (Ph. D.)
FACULTE DES SCIENCES
UNIVERSITÉ DE SHERBROOKE
?F?
Library and Archives Canada Published Heritage Branch 395 Wellington Street Ottawa ON K1A 0N4 Canada Bibliothèque et Archives Canada Direction du Patrimoine de l'édition 395, rue Wellington Ottawa ON K1A 0N4 CanadaYour file Votre référence ISBN: 978-0-494-70618-3 Our file Notre référence ISBN: 978-0-494-70618-3
NOTICE:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
telecommunication or on the Internet,
loan, distribute and sell theses worldwide, for commercial or
non-commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accordé une licence non exclusive
permettant à la Bibliothèque et Archives Canada de reproduire, publier, archiver,
sauvegarder, conserver, transmettre au public par télécommunication ou par l'Internet, prêter, distribuer et vendre des thèses partout dans le
monde, à des fins commerciales ou autres, sur
support microforme, papier, électronique et/ou
autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author's permission.
L'auteur conserve la propriété du droit d'auteur
et des droits moraux qui protège cette thèse. Ni
la thèse ni des extraits substantiels de celle-ci
ne doivent être imprimés ou autrement
reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Conformément à la loi canadienne sur la
protection de la vie privée, quelques
formulaires secondaires ont été enlevés de
cette thèse.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu
manquant.
1+1
Le 11 août 2010
lejury a accepté la thèse de Madame Edith Sanssouci
dans sa versionfinale.
Membres du jury Professeure Carole Beaulieu
Directrice de recherche
Département de biologie Professeur Ryszard Brzezinski
Membre
Département de biologie
Monsieur Hani Antoun Membre externe Université Laval Professeur Claude Déry
Président rapporteur
SOMMAIRE
Les streptomycètes, des bactéries au développement morphologique complexe, sont étudiés
depuis des décennies en raison de leurs caractéristiques particulières incluant leur capacité à
produire une grande diversité d'enzymes hydrolytiques et de molécules complexes telle que
des centaines d'antibiotiques. Streptomyces coelicolor est l'organisme modèle pour l'étude des
streptomycètes et le séquençage de son génome a ouvert la voie à l'étude de centaines de
gènes dont la fonction est jusqu'à présent inconnue.La protéine TerD du Serratia marcescens joue un rôle dans la résistance au tellurite chez cet
organisme. Plusieurs orthologues de cette protéine ont été identifiés chez le S. coelicolor. Sur
la base de la similarité de leurs séquences avec la protéine TerD, ces protéines du S. coelicolor
ont été désignées comme des protéines de résistance au tellurite. Ces mêmes protéines ont été
identifiées comme étant induites ou réprimées dans de nombreuses conditions exemptes de
tellurite, laissant croire qu'elles participent à d'autres voies métaboliques.
Au cours de ce travail de doctorat, des souches mutantes du S. coelicolor ont été produites
pour les gènes SC02367, SC02368 et SC04277, trois gènes homologues à ierO, afin
d'évaluer les répercussions de la perte de ces gènes chez la bactérie. Les souches ont été
caractérisées morphologiquement et nous avons démontré que la deletion de ces gènes nuit
sérieusement au développement de la bactérie, à la production de spores et à la vitesse de
croissance.
Parallèlement, une approche protéomique a été utilisée afin de caractériser les effets de la
deletion ou de la surexpression du gène SC02368 sur la biosynthèse des protéines intracellulaires et extracellulaires de la bactérie. Ceci a permis de démontrer qu'une grande
quantité de protéines reliées au stress sont induites chez les deux souches mutantes, laissant
présumer qu'un déséquilibre dans le taux de cette protéine induit un stress physiologique
important. Par ailleurs, différentes isoformes de la protéine SC02368 ont été identifiées tant
au niveau intracellulaire que dans le milieu extracellulaire. Par l'analyse des modifications
post-traductionnelles de ces isoformes, nous avons prouvé que celles-ci étaient méthylées sur
plusieurs acides aminés.
De plus, nous avons démontré que la transcription du gène SC02368 n'est pas induite par la
présence de tellurite. Ces résultats démontrent que les gènes homologues à terD ont une autre
fonction que la résistance au tellurite et qu'ils jouent vraisemblablement un rôle dans la
morphogénèse.
Streptomycète, différenciation, morphologie, sporulation, stress, tellurite, protéomique,
REMERCIEMENTS
La deuxième décennie du 21e siècle débute au moment où j'écris ces lignes. Avant de passer à
une autre étape de ma vie, j'aimerais prendre le temps de remercier toutes les personnes qui
m'ont soutenue et encouragée durant mes années d'études doctorales. D'abord, je remercie
grandement ma directrice de recherche, Carole Beaulieu, pour la confiance qu'elle m'a
accordée, pour son positivisme et sa précieuse aide. Elle a toujours cru en moi et en mon projet de recherche, et ce, jusqu'à la fin.
Je remercie également mes conseillers, Ryszard Brzezinski et Claude Déry pour leurs conseils
ainsi que leur pensée critique. Ils m'ont aidée à percevoir mon projet sous différents angles
afin de mieux le développer. Merci également à François Shareck avec qui j'ai collaboré pour la création des souches mutantes du S. coelicolor et qui a été membre externe sur mon comité
d'examen prédoctoral. Je suis également reconnaissante aux organismes subventionnaires qui
ont financé mon projet et desquels j'ai obtenu des bourses, soient le CRSNG et le FQRNT.
J'ai passé de très beaux moments dans le laboratoire de Carole Beaulieu et c'est en grande
partie grâce à toutes les personnes géniales qui y sont passées au cours des années. Pour leur
amitié, leurs conseils, leur écoute, leur aide, je remercie particulièrement Marie-Pierre, Gabriella, Nathalie, Isabelle, Véronique, Sandra, Geneviève, Sylvain et Anne-Marie. Merci également aux stagiaires qui m'ont aidée dans certaines étapes du projet, spécialement Véronique Barrés. Je remercie également mes parents et ma famille pour leur soutien et leurs encouragements, et ce, depuis toujours. Merci à mon conjoint Chandi pour son support
TABLE DES MATIÈRES
SOMMAIRE ?
REMERCIEMENTS · iv
TABLE DES MATIÈRES v
LISTEDESTABLEAUX vii
LISTE DES FIGURES ix
INTRODUCTION : J
CHAPITRE 1
1.0 Les gènes codant pour des protéines ayant un domaine TerD : une classe de
gènes impliqués dans la différenciation chez le Streptomyces coelicolor ....35
1.1 Sanssouci, E., Lerat, S., Grondin, G., Shareck, F. et Beaulieu, C. The TerD domain-encoding genes: a class of genes involved in Streptomyces coelicolor differentiation (soumis pour publication dans J. Bacteriol.) 37
CHAPITRE 2
2.0 Profils protéomiques des souches du S. coelicolor ayant une deletion ou
surexprimant le gène tdcfà codant pour une protéine ayant un domaine
2.1 Sanssouci, E. et Beaulieu, C. Proteomic profiles of Streptomyces coelicolor strains lacking or overexpressing the TerD domain encoding gene tddS (soumis pour publication dans J. Bacteriol.) 74
CONCLUSION 104
LISTE DES TABLEAUX
INTRODUCTION
1. Informations relatives aux protéines du S.coelicolor possédant un ou deux domaines
TerD 26
2. Résumé des publications portant sur des études protéomiques ou transcriptomiques chez
le S. coelicolor et faisant mention de l'un ou l'autre des gènes codant pour des protéines
homologues à TerD 33
CHAPITRE 1
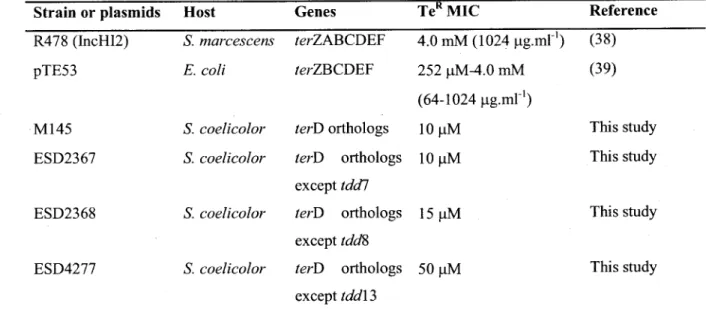
1. Bacterial strains andplasmids used in this study 47
2. Oligonucleotide primers used in this study 48
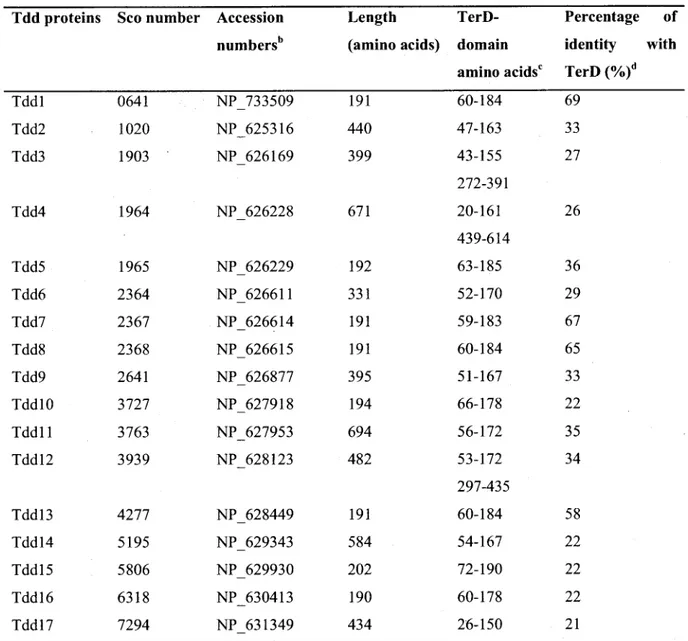
3. S. coelicolor TerD domain-containing proteins 49
4. K2Te03 Minimal inhibitory concentration (MIC) for bacterial species carrying ter
genes 51
5. Main characteristics of S. coelicolor strains differentially expressing tdc/8 53
S2. Expression of complementation genes and minimal inhibitory concentration (MIC) for
K2TeO3 in strains ESC2367, ESC2368 and ESC 4277 compared to wild-type strain M145.
The expression of tddl and tdd\2> was restored at approximately 30% in the
complementation strains ESC2367 and ESC4277, respectively. ESC4277 has an
intermediate phenotype between the wild strain and deletion strain regarding the MIC
for K2TeO3
70
CHAPITRE 2
1. Strains and plasmids used in this study
84
2. Function of intracellular and extracellular proteins differentially expressed within
LISTE DES FIGURES
INTRODUCTION
1. Représentation schématique du développement vertical chez les Streptomyces
(Manteca et al, 2007) 5
2. Réorganisation des processus biologiques durant la différenciation des hyphes
aériennes en chaînes de spores (Flärdh et Butiner, 2009) 8 3. Fonctions extracellulaires , dépendantes de MdA chez le S. coelicolor
(Chater et Chandra, 2008) 10
4. Modèle du rôle de certaines enzymes (et de certains de leurs produits) de la voie métabolique de la cysteine en réponse à la présence de K^TeOs
(Chasteen et al, 2009) 18
5. Comparaison des gènes de résistance au tellurite ter selon les différentes sources
(Taylor et al, 2002) 23
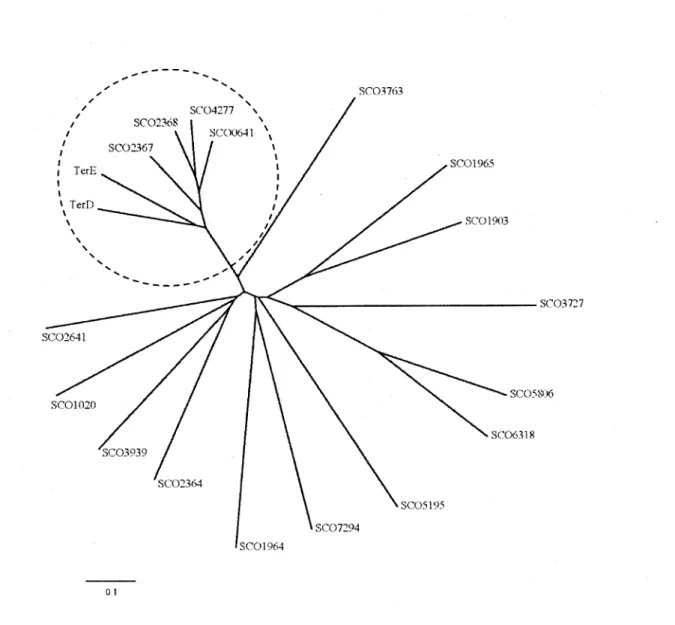
6. Arbre phylogénétique des protéines du S. coelicolor ayant un domaine TerD et des
CHAPITRE 1
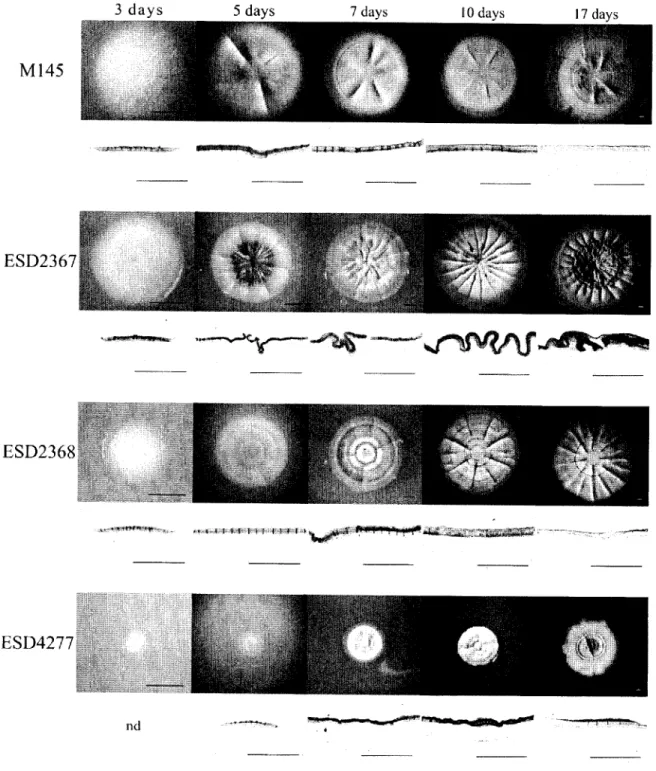
1. Deletion effect of iddi, tddS and tdd\3 on the morphology of Streptomyces coelicolor colonies. M145, ESD2367 (Atddl), ESO236S{AtddS) and ESD4277 (Atddl3) colonies
after 3, 5, 7, 10 and 17 days of growth on SLM-3 medium and corresponding transversal
cuts of the colonies observed under 50x magnification. Bars: 1 mm, nd: not
determined 52
2. Growth of (A) S. coelicolor M145 and ESD2368 (AtddS) and (B) ESM145 (carrying the
pFDES empty vector) and the over-expression strain ESS2368 in TSB. Vertical bars
indicate SD 53
3. Spores production by S. coelicolor M145, ESD2368 (AtddS), ESM145 (carrying the
pFDES empty vector), the over-expression strain ESS2368 after 6 days of growth on
SLM-3 medium, the complementation strain ESC2368 and the control strain ESCM145
(carrying the pSET152m empty vector) after 8 days of growth on SLM-3 medium. The
same strain was streaked twice at opposite positions. The gray pigmentation is associated
with the presence of spores. ESD2368 appears to produce more spores than the wild strain
whereas ESS2368 produces few spores. The complementation strain ESÇ2368 exhibits a
normal phenotype
54
4. Phase contrast microscopy of spore chains of M145, ESD2368 (AtddS), ESM145
(carrying the pFDES empty vector) and the over-expression strain ESS2368 after 4 days
of growth on SLM-3 medium. Bars: 10 µ??. ESD2368 produces very long chains of spores
compared to M145 whereas ESS2368 produces few spores of various size and shape
compared to ESM145. Complementation of ESD2368 (strain ESC2368) restores the
original phenotype as shown in the insert ofthe up left section ofthe figure
56
5. (A) Scanning electron micrographs of S. coelicolor strains M145, ESD2368 (AtddS), ESM145 (carrying the pFDES empty vector) and the over-expression strain ESS2368; bars: 1 µ??. (B) Transmission electron microscopy of S. coelicolor strains M145,
ESD2368, ESC2368, ESM145 and ESS2368; bars: 100 nm. ESD2368 spores tend to be shorter than those of M145 and exhibit a cubic shape. ESS2368 produces few spores of
variable sizes compared to ESM145. Spores of the complementation strain ESC2368
show a normal shape 57
Sl. Morphological characteristics (isolated colonies and 50x-transversal cuts) of the complementation strains ESC2367 and ESC4277 grown for 10 days on SLM-3. ESC2367
colonies were less wrinkled than in the deletion strain ESD2367 (Fig. Sl). The folding in
the mycelium appeared at 7 days instead of 5 days of growth. The diameter of 4277 is intermediary between those ofmutant sauvage. Bars: 1mm 70
CHAPITRE 2
1. Protein concentration and catechol 2,3-dioxygenase activity associated with J medium culture ofS. coelicolor strains carrying the promotorless reporter gene xylTE (strain ES-I) or the same gene coupled to the promoter of tdd% (ES-2) 85 2. Effect of exposure to 50 µ? of K2TeO3. on the 2,3-dioxygenase specific activity of
S. coelicolor strains carrying the promotorless reporter gene xy/TE (strain ES-I) or the
same gene coupled to the promoter oftddS (ES-2) 86
3. Intracellular proteome of S. coelicolor strains M145 (A), ESD2368 (B) and ESS2368 (C)
4. Extracellular proteome of S. coelicolor strains M145 (A), ESD2368 (B) and ESS2368 (C)
obtained from 48 h cultures in MG medium 89
5. Separation of intracellular (A) and extracellular proteins (B) of of S. coelicolor strains
M145 (upper gels) and ESD2368 (lower gels) by two-dimensional gel electrophoresis in a
4 to 5 pH range. (C) Post-translational modifications of Tdd8 isoforms shown in panels A
INTRODUCTION
Le sol constitue un environnement de choix pour les micro-organismes qui y trouvent les éléments nutritifs nécessaires à leur croissance tels que le carbone, l'azote, le phosphate et les différents minéraux. Les milliards de bactéries, champignons, protozoaires, virus, nematodes
et microarthropodes qui peuplent le sol participent au recyclage de la matière organique et la
rendent ainsi disponible pour les plantes. Ces êtres vivent en communautés intimement liées et
entrent en compétition pour les ressources du milieu. L'écologie microbienne étudie le
comportement et les activités des micro-organismes dans leurs environnements naturels ce qui
inclut les interactions entre eux et avec leur milieu. Au sein de cet environnement changeant,
les micro-organismes sont sujets à de nombreux stress biotiques et abiotiques qui influencent
continuellement leur physiologie. Parmi ces stress, on retrouve les variations de température et
de pH, les stress oxydatifs et osmotiques, la disponibilité des minéraux, etc. Pour réussir à
occuper une niche écologique donnée, les micro-organismes mettent en branle de nombreux
processus physiologiques. Ils possèdent diverses classes de gènes leur permettant d'acquérir
les nutriments dont ils ont besoin et de se défendre contre la présence de molécules toxiquesde l'environnement. Parmi ces gènes, certains codent pour des enzymes hydrolytiques
participants à dégradation de divers polymères tels que la cellulose, le xylane, l'amidon et la
chitine; d'autres permettent la synthèse de metabolites tels que les antibiotiques. Par ailleurs,
de nombreux échanges génétiques ont cours entre les micro-organismes. Ainsi, des gènes de
résistance aux antibiotiques, aux métaux lourds et à certains composés organiques sont, entre
autres, ainsi disséminés entre les espèces en réponse aux pressions de sélection de
l'environnement. Ces gènes évoluent dans le temps et les protéines codées s'en trouvent ainsi
modifiées.
Des séquences de protéines ou de gènes sont dites homologues quand elles découlent d'un
ancêtre commun. Plus la similarité entre deux séquences est grande, plus la probabilité queleur origine soit indépendante et leur similarité due à la chance est basse. La similarité peut être au niveau de la séquence ou de la structure, et cette dernière est d'ailleurs bien mieux conservée durant l'évolution. L'évolution de séquences est dite convergente quand plusieurs séquences sont présumées avoir évolué de façon indépendante vers une séquence similaire. Elle est dite divergente quand une séquence est présumée être l'ancêtre commun de plusieurs séquences similaires. L'analyse de séquences d'ADN et de protéines est utilisée pour identifier des homologies et ainsi prédire la fonction biologique des gènes ou des protéines. Elle peut également servir à reconstruire l'historique de l'évolution (Koonin et Galperin, 2002). Il existe deux types fondamentalement différents de gènes homologues : les orthologues qui ont évolué de façon verticale à partir d'un ancêtre commun, et les paralogues qui découlent d'une duplication de gène (Koonin, 2005). L'étude des gènes homologues peut se révéler des plus complexe suivant les multiples événements de spéciation, de duplication, de transfert horizontal, de perte de gènes et de réarrangements génétiques pouvant avoir eu lieu au cours
de l'évolution.
Au cours des dernières années, le séquençage exhaustif de nombreux génomes a révélé une panoplie de nouvelles séquences. Des fonctions putatives ont été assignées à ces nouveaux gènes selon leur taux de similarité avec des gènes aux fonctions préalablement connues. Mais qu'en est-il des gènes qui n'ont pas de similarité avec des gènes de fonction connue? Et les fonctions assignées en raison de la similarité de séquences sont-elles toujours exactes? Énormément de travail de validation devra encore être réalisé afin de s'assurer de la véracité de ces assignations.
Les streptomycètes
Les streptomycètes font partie de l'ordre des actinomycétales et sont des bactéries aérobies, à Gram positif. Ils sont caractérisés par un génome riche en guanine et cytosine ainsi qu'un
chromosome linéaire de plus de 8 M de paires de bases. Ces bactéries au cycle de vie
complexe colonisent divers milieux de vie, tels les sols, l'air et l'eau (Kieser et al, 2000;
Peltola et al, 2001; Pathom-Aree et al, 2006). Ils comptent pour environ 10 % de la flore
microbienne du sol (Janssen, 2006). La plupart des streptomycètes sont saprophytes et ils sont
fréquemment retrouvés dans la rhizosphère où ils colonisent les racines des plantes. Bien que
la plupart des actinomycètes soient non pathogènes, certaines espèces comme Streptomyces
scabiei, l'agent causal de la gale commune de la pomme de terre, causent des maladies.
Plusieurs actinomycètes non pathogènes sont, par ailleurs, de bons agents de lutte biologique
(You et al, 2007; Aanen et al, 2009; El-Tarabily et al, 2009)
L'intérêt de l'étude de ces bactéries vient en grande partie de leur capacité à produire une
grande variété de metabolites secondaires aux activités biologiques diverses. Parmi ceux-ci, on
retrouve des immunosuppresseurs comme la rapamycine produite par Streptomyces
hygroscopicus utilisée comme agent antirejet lors de greffe d'organes, des agents
antitumoraux (ex : bléomycine, daunorubicine, mitomycine C) et de très nombreux
antibiotiques (ex : chloramphenicol, erythromycin, gentamicine, kanamycine, streptomycine)
(Kieser et al, 2000). De 5000 à 10000 antibiotiques ont été découverts chez les bactéries et les
champignons à ce jour et le seul genre bactérien Streptomyces serait responsable de plus de
50 % de cette production (Challis et Hopwood, 2003). Les streptomycètes sont par ailleurs de
grands producteurs d'enzymes hydrolytiques (ex. : cellulase, ligninase, xylanase, chitinase)
leur permettant de dégrader des polymères biologiques complexes. Streptomyces coelicolor et son cycle de vie
En 2002, le Streptomyces coelicolor A3(2) a été la première espèce de streptomycètes dont le
génome a été complètement séquence (Bentley et al, 2002). Celui-ci comporte 8 667 507
paires de bases avec 72 % de G+C. Ce flot d'informations nouvelles a propulsé les
connaissances sur cet organisme modèle pourtant étudié depuis des décennies. En effet, des gènes peuvent à présent être amplifiés par PCR très rapidement, des ARNm peuvent être quantifiés aisément grâce à la PCR quantitative en temps réel, des techniques d'interruption génique ciblées peuvent être utilisées pour la création de souches mutantes en remplacement des techniques de mutagenèse aléatoire', etc. Cependant, des 7825 cadres de lecture ouverts du S. coelicolor, seuls 5492 se sont fait assigner une fonction, alors que 2333 sont toujours de
fonction inconnue (Borodina et ai, 2005).
Le cycle de vie du S. coelicolor est étudié scrupuleusement depuis plusieurs années. Les données actuelles permettent de déduire les événements séquentiels suivants. Lorsque les
conditions environnementales sont favorables, le processus de germination des spores se met
en branle. Un ou plusieurs tubes germinatifs émergent d'une spore et s'allongent par extension des hyphes au niveau apical. Les hyphes coenocytiques forment des branchements et s'enchevêtrent pour donner naissance au mycélium basai. Celui-ci poursuit sa croissance en se
ramifiant dans toutes les directions afin d'atteindre les ressources du milieu (Kieser et al,
2000). Récemment, il a été démontré qu'en début de croissance, une portion du mycélium basai subit une mort cellulaire programmée. Ceci provoque une mosaïque de segments viables et non viables. Un second mycélium se développe alors, à partir des segments viables, sous forme d'îlots et s'étend progressivement (Manteca et al, 2007). Lorsque les ressources du milieu deviennent limitées, le processus de différenciation cellulaire débute. Le mycélium basai donne alors naissance au mycélium aérien qui quitte le milieu aqueux et émerge dans les airs. Ce nouveau mycélium est caractérisé par une couleur blanchâtre, un aspect duveteux et des hyphes non branchées. L'érection du mycélium dans les airs est rendue possible grâce à certaines molécules particulières. Premièrement, SapB (spore-associated protein B) est sécrétée durant la phase de développement végétative, en milieu riche seulement, et agirait en tant que surfactant en recouvrant les hyphes aériennes émergentes et l'interface eau-air afin de faciliter l'érection des hyphes dans l'atmosphère. Deuxièmement, des protéines nommées « long chaplin » sont exportées et viennent s'attacher de façon covalente à la paroi cellulaire des hyphes aériennes. Elles sont ensuite hétérodimérisées par les « shorts chaplins » afin de
former des filaments de chaplins très hydrophobes. Finalement, ces filaments sont organisés à
un niveau supérieur par les rodlins afin de former des paires de « rodlets ». Cette ultrastructure enveloppant les hyphes aériennes vient aussi faciliter l'érection dans les airs, et ce, en milieu
riche comme en milieu pauvre (Elliot et al., 2008; Flärdh et Butiner, 2009). Le second
mycélium basai se distingue par des hyphes recouvertes ou non de « rodlets » et les hyphes
avec « rodlets » sont considérées comme étant précurseures du mycélium aérien caractérisé
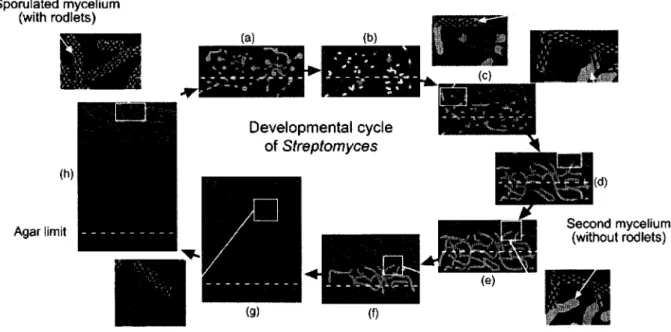
par la présence de ces ultrastructures (Manteca et al., 2007). La figure 1 illustre les différentes
étapes du développement des streptomycètes démontrées chez le S. coelicolor et le Streptomyces lividans. Sporulated mycelium/ ¡XL. Jl _*_ * (with rodlets)
*?3µ??
(a bEeBBsII BMBH S
e) Bi Developmental cycle of Streptomyces h (d Second myceliumAgar limit (without rodlets)
M
<e
(9) ffl
Figure 1: Représentation schématique du développement vertical chez les Streptomyces. Le rouge représente des cellules mortes alors que le vert représente des cellules vivantes. Les lignes grises symbolisent les protéines de type rodlet. Les formes circulaires en phase (g)
correspondent au second mycélium coenocytique où les nucleoides amorcent leur division. Les lignes pointillées jaunes représentent la limite de Gagar. Les zones noires en (g) et (h) indiquent une absence complète de fluorescence sous le mycélium présporulant et sporulant due à la désintégration des hyphes et à la dégradation des acides nucléiques. Cette figure est
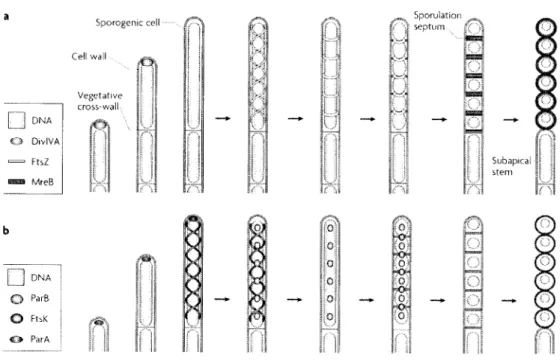
La dernière étape du cycle de vie des streptomycètes est la réorganisation de plusieurs processus cellulaires menant à la différenciation des cellules apicales en chaînes de spores. Durant les étapes initiales du développement des hyphes aériennes, un long compartiment apical coenocytique nommé cellule sporogénique se développe. Cinquante copies ou plus du chromosome peuyent être produites dans chaque cellule sporogénique. La croissance par extension apicale s'arrête et une multiple division synchronisée est initiée. Celle-ci est dirigée par la protéine FtsZ, un homologue bactérien de la tubuline, qui s'assemble en filaments hélicaux ensuite remodelés en anneaux régulièrement espacés, nommés « Z ring », qui définissent les sites de division. D'autres protéines de division sont également mobilisées pour
participer à ce processus. À la suite de la formation des septa, une épaisse paroi cellulaire est
synthétisée autour des préspores (Fig. 2 A). (Flärdh et Buttner, 2009). La ségrégation des
chromosomes se fait de façon concomitante à l'aide de diverses protéines qui séparent les chromosomes de façon à ce qu'il y en ait qu'un seul par préspore. Les nucleoides se condensent dans les spores en maturation et finalement, les chaînes de spores unigénomiques sont formées (Fig. 2 B). (Flärdh et Buttner, 2009). Les spores sont caractérisées par une coloration grise due à la présence d'un composé aromatique de type polycétide (Kieser et al., 2000). Ces spores de reproduction, résistantes à la dessiccation, fournissent un mécanisme de dissémination efficace vers de nouveaux environnements pour ces bactéries non mobiles et sans capacité de chimiotactisme (Elliot et al., 2008).
En plus de sa différenciation morphologique qui réfère strictement à la production de chaînes de spores, le S. coelicolor se différencie physiologiquement, c'est-à-dire qu'elle produit des metabolites secondaires tels des antibiotiques. En culture liquide, la production d'antibiotiques débute avec l'entrée de la culture en phase stationnaire alors qu'en culture sur agar, elle coïncide avec la différenciation morphologique. Le S. coelicolor produit plusieurs antibiotiques dont le plus connu est l'actinorhodine, un pigment rouge-bleu indicateur de l'acidité du milieu. Une forme modifiée, la ?-actinorhodine de couleur bleue, est sécrétée dans le milieu extracellulaire. Le S. coelicolor produit également l'undecylprodigiosine, un pigment rouge, qui est en fait un mélange d'au moins quatre prodiginines dont l'undecylprodigiosine et
la butylcycloheptylprodiginine forment les composantes principales. Le «calcium-dependent
lipopeptide antibiotic» (CDA) est, comme son nom l'indique, produit en présence de calcium
et inhibe la croissance de Bacillus mycoides. Finalement, la methylenomycine est produitegrâce à des gènes portés sur le plasmide SCPl (Kieser et al, 2000).
Les étapes de la différenciation cellulaire sont finement régulées au niveau génétique. Un
grand nombre de gènes requis pour la formation du mycélium aérien a été identifié. Ces gènes
incluent MdA, MdQ, MdC, MdD, MdG, MdH, MdJ, MdK, MdL, MdM, MdN, ramR, ramCSAB,
chpA-H, citA, acoA, cya, clp?\, catB, dasR et MgA. Plusieurs de ces gènes ont été désignés
Md (pour « bald ») en référence à l'absence d'hyphes aériennes chez les souches mutantes
pour ces gènes. La majorité des gènes identifiés comme étant importants pour la formation du
mycélium aérien code pour des protéines de régulation. Cependant, certains gènes codent aussi
pour des molécules de structure. Ainsi, les gènes du cluster ram permettent la synthèse du
peptide SapB alors que les gènes chpA-H codent pour les chaplins (Elliot et al, 2008).
D'autres gènes impliqués dans le développement, souvent désignés whi, ont été identifiés. Ces
gènes, lorsqu'ils sont mutés, permettent la formation du mycélium aérien par la souche, mais
non la différenciation morphologique en spores grises, d'où l'appellation whi en référence à la
couleur blanche (« white ») des colonies. Ces gènes incluent whiA, whiB, whiO, whiE, whiG,
whiH, whil, whii, sigVJtsZ, ssgA, ssgB, ssgC-F, ssgR et samR (Elliot et al, 2008).
Fait intéressant, des mutations des gènes MdA, MdB, MdC, MdD, MdG, MdH et MdS, abaissent
la production d'antibiotiques par la souche, reflétant le lien possible entre la régulation du
métabolisme secondaire et celle de la différenciation morphologique (Elliot et al, 2008). La
mutation du gène MdA constitue un exemple concret et bien étudié de ce lien. En effet, MdA
code pour le seul ARN de transfert (ARNt) permettant la traduction du codon rare UUA
codant pour une leucine (Lawlor et al, 1987). Typiquement, 2-3 % des gènes de
Sporogonie cell·· Qdna O DivlVA = FtsZ Era MreB Cell wall Vegetative cross-wall· D dna O ParB O FtiK O ParA ìM°*% r4 e ' 0 I ?? ? i 1-4 M Sporulation septum ?; ?? i ¦SO? (öj ?' (oí Il Subapical
Figure 2: Réorganisation des processus biologiques durant la différenciation des hyphes
aériennes en chaînes de spores, a. Orchestration de l'assemblage de la paroi cellulaire et de la
division cellulaire. L'hyphe aérienne croît par extension apicale et accumule la protéine
DivlVA à son extrémité. La formation d'une cellule sporogénique apicale (qui est souvent
enroulée et est généralement plus longue que celle indiquée dans le présent schéma simplifié)
implique l'arrêt de la croissance. FtsZ s'assemble en filaments hélicoïdaux, qui sont remodelés
en anneaux Z régulièrement espacés, qui dirigent la septation en spores. Après l'achèvement des septa, les préspores forment une épaisse paroi à l'aide de l'actine bactérienne MreB.
Initialement, MreB se localise au niveau des septa se refermant, mais ensuite se répand tout
autour des spores en développement, b. ségrégation des chromosomes. L'ATPase ParA est
d'abord retrouvée à l'extrémité des jeunes hyphes aériennes, puis formes des filaments
hélicoïdaux le long de la cellule sporogénique. Les protéines ParB s'assemblent en complexes
nucléoprotéiques au niveau des régions chromosomiques oriC. La répartition des foyers
ParB-oriC le long de la cellule sporogénique semble être dirigée par ParA. La croissance des septa
débute alors que la ségrégation du matériel génétique n'est pas encore terminée et la
translocase FtsK est dirigée vers les sites de division et aide à débarrasser l'ADN des septa se
refermant. Finalement, les nucleoides se condensent à l'intérieur des spores en maturation.
Un des gènes de régulation du cluster act permettant la synthèse de l'actinorhodine, le gène
actII-ORF4, contient un codon TTA et est un activateur clé de la plupart, sinon de tous les
gènes de la voie métabolique de l'actinorhodine. Ainsi, l'incapacité à produire cet antibiotique
chez la souche mutante pour le gène bldA s'explique par l'incapacité à traduire le codon TTA.
Le gène adpA, originalement identifié bldH, contient également un codon TTA et est donc
dépendant de BIdA pour sa traduction. AdpA est un régulateur de type AraC requis pour la
formation du mycélium aérien (Bentley et al, 2002). Les cibles de ce régulateur incluent des
gènes codant pour des proteases, des inhibiteurs de proteases, un facteur sigma et plusieurs
autres protéines impliquées dans le développement (Chater et Chandra, 2008). Il a été
démontré que le phénotype « bald » de la souche mutante pour bldA est largement causé par la
présence du codon TTA dans le gène adpA, puisqu'en changeant ce codon, la capacité de
produire un mycélium aérien et des spores est restaurée (Nguyen et al., 2003; Takano et al,
2003). Ainsi, l'effet de bldA sur le développement implique probablement, du moins chez le
S. coelicolor, une perte d'expression d'une partie du régulon du régulateur AdpA.
Il a été démontré, chez d'autres streptomycètes que des inhibiteurs de proteases, régulés par
AdpA, sont impliqués dans une cascade de proteases extracellulaires. Des évidences indiquent
que cette cascade joue un rôle significatif dans le développement, apparemment dans un
processus d'autolyse par lequel la biomasse végétative est convertie en spores (Kim et Lee,
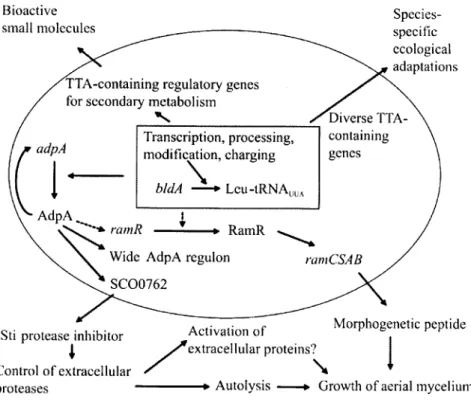
1995; Chater, 2006; Hirano et al, 2006; Manteca et al, 2006a). La figure 3 résume les
implications connues de bldA dans ces processus extracellulaires. Par ailleurs, des études
transcriptomiques ont démontré que l'expression de 147 gènes est modulée chez une souche
mutante pour le gène bldA. Ces gènes ne contiennent, pour la grande majorité, aucun codon
TTA. De plus, des études protéomiques ont également démontré que plusieurs protéines sont modulées chez la même souche mutante (Chater et Chandra, 2008). Il reste encore une grande quantité de travail à accomplir afin d'élucider les liens possibles entre tous ces interactants et
l'exemple du gène bldA démontre toute la complexité de la régulation génétique chez les
Bioactive small molecules
TTA-containing regulatory genes for secondary metabolism
AdpA.^ Species-specific ecological adaptations Transcription, processing, modification, charging NdA *Leu-tRNA„ ¦* RamR
Wide AdpA regulon
SCO0762
Sti protease inhibitor
Control of extracellular proteases Activation of Diverse containing genes ramCSAB Morphogenese peptide extracellular proteins?
Autolysis \Growth of aerial mycelium
Figure 3: Fonctions extracellulaires dépendantes de bidA chez le S. coelicolor. Les flèches
pointillées indiquent une étape démontrée chez S. griseus. Tous les autres liens sont répandus
au sein des streptomycètes. Chez S. griseus, les orthologues des gènes du cluster ram sont
connus sous le nom de gènes amfet ils mènent à la synthèse du peptide extracellulaire AmfS
(Ueda et al, 2002) alors que le produit des gènes ram est SapB (Kodani et al, 2004). Cette
figure est tirée de Chater et Chandra (2008).
En effet, malgré l'abondance d'informations accumulées au cours des années sur la
différenciation morphologique chez les streptomycètes, il est encore impossible de tracer un
portrait clair expliquant comment les voies de signalisation et de régulation génétique sont
La réponse aux stress chez les streptomycètes
La capacité de percevoir et de répondre aux changements environnementaux est cruciale pour
la survie des bactéries. Afin d'arriver à capter les signaux de l'environnement et à mettre enplace une réponse adaptative à ceux-ci, les bactéries utilisent différents mécanismes de
signalisation cellulaire. Ces trois mécanismes fondamentaux sont la régulation génétique à une
composante, à deux composantes et l'utilisation de facteurs sigma alternatifs. (Staron et al,
2009)
La régulation génétique à deux composantes implique deux protéines, une protéine senseur,
l'histidine kinase et une protéine de réponse agissant au niveau de la transcription.
Typiquement, la protéine senseur est liée à la membrane plasmique et possède un domaine
extracellulaire lui permettant de percevoir le stimulus, le domaine senseur ou «input domain».
L'activation de la protéine senseur induit une autophosphorylation d'un résidu histidine
spécifique du domaine histidine kinase. Le groupe phosphate est ensuite transféré à un résidu
aspartate spécifique du domaine receveur de la protéine régulatrice se trouvant dans le cytosol
de la bactérie. Ceci active le domaine effecteur ou «output domain» du régulateur qui instaurealors la réponse cellulaire adéquate. La majorité des systèmes de régulations génétiques à deux
composantes caractérisée expérimentalement régulent l'expression génétique au niveau de la
transcription à l'aide d'un domaine HTH pour «helix-turn-helix» qui se lie à l'ADN et activent
les gènes cibles (Ulrich et al, 2005). D'autres régulateurs contiennent des domaines
enzymatiques comme des di-guanylate cyclase (Paul et al, 2004). Dans le cas des systèmes de
régulation à une composante, le «input domain» et le «output domain» sont contenus dans une
seule et même protéine, il n'y a ni domaine histidine kinase ni domaine receveur et les signaux
perçus ne sont qu'intracellulaires. Ce mode de régulation génétique serait précurseur du mode
à deux composantes permettant de capter les signaux extracellulaires et ainsi de mieux
Le S. coelicolor possède 84 gènes codant pour des senseurs kinases dont 67 sont adjacents à
des gènes codant pour des régulateurs de réponse, au nombre de 80 au total (Hurchings et al,
2004). Jusqu'à présent, les fonctions et les cibles de ces 67 paires de gènes sont très peu
connues. On sait cependant que plusieurs sont impliqués dans la régulation de divers processus
cellulaires tels que !'osmoregulation, la croissance et la différenciation cellulaire (Hurchings et
al, 2004). Quelques exemples de systèmes de régulation génétique à 2 composantes bien
étudiés chez le S. coelicolor incluent le système PhoP-R qui régule la réponse au manque de
phosphate (Sola-Landa et al, 2003; Rodroguez-Garcia et al, 2007) et les systèmes CseB-C et
Van R-S qui activent des gènes impliqués dans l'intégrité de la paroi cellulaire lors de
l'exposition à la vancomycine et à d'autres antibiotiques spécifiques à la paroi cellulaire
(Paget et al, 1999; Hong et al, 2002; Hutching et al, 2006).
L'utilisation de facteurs sigma alternatifs constitue un autre moyen d'agir au niveau
transcriptionnel en réponse à des conditions changeantes. En effet, le facteur sigma est une·
composante essentielle de TARN polymerase et est responsable de la spécificité de celle-ci
(Helmann et Chamberlin, 1988). Le facteur sigma principal, qui est essentiel pour la
transcription dans la phase de croissance exponentielle, est associé de manière réversible à
GARN polymerase et peut être remplacé par un facteur sigma alternatif. Toutes les bactéries
ne possèdent qu'un seul facteur sigma principal responsable du niveau d'expression basai de
la plupart des gènes. Par contre, la plupart des génomes bactériens, spécialement ceux des
espèces au développement morphologique complexe, possèdent plusieurs facteurs sigma
alternatifs. Ceux-ci coordonnent l'expression de gènes impliqués dans diverses fonctions telles
que la réponse aux stress, le développement morphologique et l'acquisition du fer (Paget et
Helmann, 2003) en redirigeant l'ARN polymerase vers un promoteur alternatif, changeant
ainsi le type de gènes réprimés (Helmann et Chamberlin, 1988). En l'absence de stimulus, la
plupart des facteurs sigma sont séquestrés par un facteur antisigma (Brown et Hughes, 1995;
Les facteurs sigma sont divisés en deux grandes familles dont les séquences ont peu de
similarité, la famille deo 70 et celle de s54. La famille de s™ est subdivisée en quatre groupes
phylogénétiques. Le groupe 1 regroupe les facteurs sigma primaires essentiels, chacun étant
fortement relié au facteur principal chez le E. coli, s70. Les facteurs du groupe 2 sont fortement
reliés au facteur primaire, mais ne sont pas essentiels pour la croissance bactérienne. Lesfacteurs du groupe 3 sont moins similaires à s70 et activent habituellement des régulons en
réponse à des signaux spécifiques tels qu'un choc thermique ou un changement dans la phase
du développement. Finalement, les facteurs du groupe 4 sont les facteurs ECF pour
«extracytoplasmic function». Ces facteurs répondent à des signaux de l'environnement tels
que la présence de protéines mal repliées dans l'espace périplasmique.
Le nombre de membres des groupes 2, 3 et 4 varie selon l'espèce bactérienne, reflétant en
partie les caractéristiques physiologiques et développementales des différents organismes
(Paget et Helmann, 2003). Le séquençage du génome du S. coelicolor a révélé l'existence de 66 (Hahn et al, 2003) facteurs sigma putatifs dont 51 ECF (Paget et al, 2002). Ces facteurs jouent donc un rôle majeur dans la régulation de la transcription chez le S. coelicolor. Le facteur sigma principal du S. coelicolor est HrdB, un homologue de RpoD du E. coli. Parmi
les facteurs ECF du S. coelicolor on retrouve, entre autres, s? impliqué dans la détection et la
réponse au stress relié à la paroi,s R impliqué dans la détection et la réponse au stress oxydatif
et aBldN impliqué la différenciation morphologique (Paget et al, 2002).
Le tellurite, la résistance au tellurite, TerD et le domaine TerD
Le tellurium ou tellure (Te) est un élément semi-métallique rare de couleur blanc-argent
présentant des caractéristiques à la fois métalliques et non métalliques et donc qualifié de
métalloïde. Son numéro atomique est 52 et il fait partie du même groupe que l'oxygène, le souffre, le sélénium et le polonium. Ces éléments du groupe 16 réfèrent collectivement aux
chalcogènes, signifiant formeurs de minerai et se rapportant à l'abondance des oxydes,
sulfides, sélénides et tellurides parmi les minerais (Jensen, 1997; Fischer, 2001). La quantité
de Te dans la croûte terrestre est de 0,005 ppm. Il est retrouvé dans divers minerais, le plus souvent sous forme de tellurures (du cuivre, de l'or, du plomb et de l'argent, par exemple) et
parfois sous forme d'impuretés dans certains sulfures métalliques. Il est notamment obtenu
comme sous-produit lors de l'affinage électrolytique du cuivre. Le tellure est utilisé en
métallurgie, en chimie et en électronique (Taylor, 1999).
Les oxyanions de tellurium (tellurite, TeO32", et tellurate, TeO42") ont des effets toxiques chez
plusieurs organismes, et ce, même à de très faibles concentrations. En 1932, Alexander
Fleming a rapporté les propriétés antibactériennes de la pénicilline et du tellurite de potassium
(K2TeO3). À partir de cette date, le tellurite a été utilisé dans la composition de milieux
sélectifs pour l'isolement d'agents pathogènes tels que le Corynebacterium diphteriae, le
Staphylococcus aureus, le Vibrio cholerea et le Shigella sp. (Taylor, 1999). Aujourd'hui, il est
surtout utilisé pour l'isolement sélectif de souches Escherichia, coli STEC (Shiga
toxin-producing E. coli) 0157 :H7 responsables de colites hémorragiques et du syndrome urémique
hémolytique (De Boer et Heuvelink, 2000). Le milieu le plus utilisé est le cefixime-tellurite
sorbitol; MacConkey (CT-SMAC) qui inhibe la plupart des souches E. coli de la flore normale
intestinale alors qu'il permet la croissance des souches STEC 0157 :H7. La résistance de ces
souches au tellurite est due à la présence de sept gènes, terZ, terA, terC, terD, terE et ter?.
Une étude récente, portant sur la variabilité dans la résistance au tellurite et la présence de
l'opéron ter chez des souches STEC isolées d'humains, d'animaux et de nourriture, a
démontré que ce milieu est souhaitable pour la détection et l'isolement de souches NSF
(non-sorbitol-fermenting) STEC 0157 :H7/NM (non-mobile), mais non pour les souches
émergentes SF STEC 0157 :NM et la majorité des STEC non-0157, ces dernières ne
Les effets toxiques du tellurite se font ressentir à des concentrations 100 fois plus faibles que
dans le cas d'autres métaux et métalloïdes comme le sélénium, le chrome, le fer, le mercure, le
cadmium et le cuivre (Nies, 1999). Le mécanisme précis régissant la toxicité du tellurite n'est
pas connu et aucun consensus n'a pu être établi sur le mécanisme exact de résistance au
tellurite sur la base des séquences des gènes qui y sont associés en raison de la grande diversité au sein de celles-ci. Quelques mécanismes ont été proposés, mais ils ne sont pas
soutenus par des évidences expérimentales. Ceux-ci incluent l'exclusion directe du tellurite, la
conversion en des formes volatiles ou alkylées et la réduction enzymatique ou non
enzymatique en tellurium élémentaire (Te0) provoquant une accumulation de dépôts noirs à
l'intérieur de la cellule. Il a longtemps été considéré que la toxicité du tellurium était due à sespropriétés oxydantes puissantes. L'observation de dépôts de Te0 à l'intérieur de cellules hôtes
de plasmides de résistance au tellurite, en présence de tellurite, a mené à l'hypothèse que la
réduction du K2TeO3 en Te0 était le mécanisme de detoxification (Taylor et al, 1988).
Cependant, les cellules du E. coli, dépourvues de tels plasmides réduisent quand même le
tellurite grâce à leur nitrate reductase, mais meurent au cours du processus (Avazari et al,
1997). Ce mécanisme de réduction, partagé par la plupart des micro-organismes, serait ainsi
seulement responsable d'un niveau basai de résistance. Par ailleurs, des études utilisant du tellurite radiomarqué ont démontré qu'il n'y avait pas de différence dans l'entrée ou la sortie de tellurite entre des souches E. coli sans plasmide de résistance et celles contenant un ou
l'autre des plasmides de résistance au tellurite (Lloyd-Jones et al, 1991). L'exclusion directe
du tellurite ne semble donc pas expliquer la résistance de ces souches. Un autre mode de résistance est la transformation du composé toxique en une forme moins toxique. La méthylation du tellurite donne des produits volatils comme le diméthyltelluride (Basnayake et al, 2001). Le gène tmp de Pseudomonas syringae pathovar pisi code pour une thiopurine méthyltransférase. Les cellules du E. coli hôtes d'un plasmide contenant le gène tmp ont une
concentration minimale inhibitrice (CMI) au tellurite entre 128 et 256 µg ml"1. Il a donc été
proposé que la résistance au tellurite passe par une volatilisation du tellurite en
diméthyltelluride (Cournoyer et al, 1998). Cette piste n'est pas à négliger, mais reste encore à
Jusqu'à présent, les études scientifiques ont démontré qu'une trentaine de gènes ou de groupe
de gènes étaient reliés, de façon directe ou indirecte, à la résistance au tellurite (Chasteen et
al, 2009). Certains gènes conférant une résistance au tellurite ont été caractérisés chez les
bactéries à Gram négatif, dont plusieurs sont retrouvés sur des plasmides, vecteurs importants
de transferts horizontaux entre les espèces. Le chromosome du E. coli contient les gènes tehA
ettehB. Ces gènes confèrent une résistance au tellurite lorsqu'ils sont clones dans un vecteur à
haut nombre de copies. De plus, la protéine TehA confère une résistance aux antiseptiques et
aux désinfectants (Chasteen et al, 2009). L'opéron MlA [MaA, te/A (MaB), telB (MaQ] est
retrouvé sur le plasmide du groupe d'incompatibilité P (IncP) RK2, isolé de Klebsiella
aerogenes (Ingram et al, 1973). Une mauvaise régulation de MlA mène à la mort des E. coli
hôtes (Goncharoff et al, 1991). Ce locus a aussi été associé à la résistance au tellurite et à une
inhibition de la fertilité. La résistance au tellurite conférée par cet opéron est cryptique. Une
simple substitution de T en A dans le gène telB, changeant une serine en cysteine à la position
125, permet au plasmide de conférer une résistance. La présence des trois gènes de l'opéron
est requise pour la résistance et un niveau d'expression plus bas est associé avec des taux de
résistance plus élevés. De plus, deux résidus cysteine du gène telB sont requis pour la
résistance. La mutation de ces résidus mène à une baisse marquée du niveau de résistance quipasse de 256 µg ml"1 à 8-16 µg ml"! (Turner et al, 1994). Chez le Rhodobacter sphaeroides,
les gènes trgA et trgB codent pour des protéines associées à la membrane qui défèrent une
résistance lorsqu'ils sont introduits chez le Paracoccus denitrificans (O'Gara et al, 1997). Les
plasmides d'incompatibilité HI2 (IncHI2) confèrent une résistance à de nombreux
antibiotiques, à des métaux lourds et à des colicines, des toxines formant des pores dans la
membrane cellulaire de souches E. coli sensibles. Les plasmides IncHI2 R478 deS. marcescens et pMER610 de Alcaligenes spp. contiennent l'opéron ter (terZABCDEF). Cet
opéron semble, en plus de permettre la résistance au tellurite, contrôler la résistance à des
bacteriophages et à des colicines. Les séquences en acides aminés des protéines codées par ces
gènes ne présentent pas de similarité avec des séquences connues sauf celles de TerD et TerE
qui possèdent entre 43 et 51 % d'identité avec les protéines de liaison à l'AMP cyclique
(CABP-I et CABP-2) de Dictyostelium discoideum, une amibe se nourrissant de bactéries et
de champignons du sol (Grant et Tsang, 1990). Dans l'ensemble, il semble que les différentes
espèces bactériennes ont trouvé chacune un moyen unique de répondre à la toxicité du
tellurite. Néanmoins pour chacun des groupes de gènes responsables de la résistance au
tellurite, de nombreux orthologues existent chez d'autres espèces plus ou moins apparentées.
Il existe très peu de similarité entre les séquences des gènes conférant une résistance au
tellurite. Les mécanismes régissant cette résistance ne sont que spéculatifs, mis à part celui de
l'opéron asr qui permet la résistance à l'arséniate, l'arsénite, l'antimonite et au tellurite par un
mécanisme de pompe à efflux (Turner et al, 1992). Plusieurs études ont démontré que certains
gènes du métabolisme basai, lorsque surexprimés, augmentaient la tolérance au tellurite. C'est
le cas du gène çysK, codant pour une cysteine synthase, de plusieurs espèces dont le
Geobacillus stearothermophilus V, le Staphylococcus aureus, le E. coli et le Rhodobacter
sphaeroides (Vasques et al, 2001; Lithgow et al, 2004; Alonso et al, 2000; O'Gara et al,
1997) lorsqu'exprimé dans une souche hétérologue. Certaines informations indiquent que ceci
est en lien avec le mécanisme de toxicité du tellurite. En effet, il a été proposé que la toxicité
du tellurite soit due à l'oxydation des thiols cellulaires, comme le glutathion, ou à la
production d'anions superoxydes durant la réduction du tellurite, causant ainsi une instabilité
menant à un stress oxydatif intracellulaire (Fuentes et al, 2007). Chez les bactéries, le
glutathion est surtout présent chez les bactéries aérobies à Gram négatif, et très rarement
présent chez les anaerobes et les bactéries à Gram positif. En plus de son rôle clé dans le
maintien des protéines thiols dans un état oxydatif approprié, le glutathion protège contre les
baisses de pH, les composés chlorés, les stress osmotique et oxydatif et participe à la
régulation des niveaux de potassium (Masip et al, 2006).
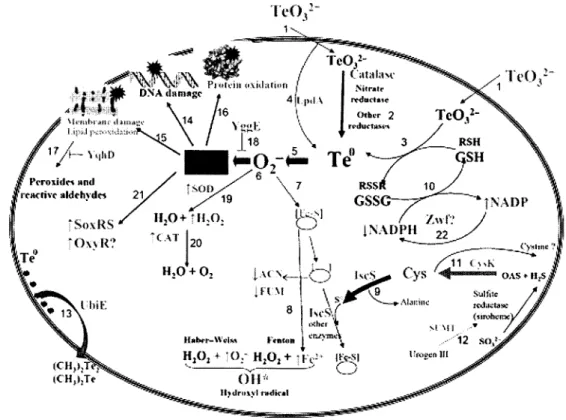
Le modèle le plus récent de la toxicité du tellurite chez le E. coli est illustré à la figure 4. Pour
exercer sa toxicité, le K2TeO32" doit entrer dans sa cellule cible, et ceci se fait
vraisemblablement par le système de transport du phosphate. Dans le périplasme, il interagit
avec les enzymes du système de catalyse des ponts disulfure, modifiant ainsi l'intégrité de la
membrane et de la paroi cellulaire. Le tellurite n'ayant pas réagi dans le périplasme est
transporté vers le cytoplasme par des transporteurs du phosphate où la nitrate reductase, la
catalase, la dihydrolipoamide déshydrogénase, des thiols réduits (particulièrement le
glutathion) et d'autres reductases non spécifiques le réduisent en tellurium élémentaire (Te0).
Te(V"
1-. -***
reo,2
^f^WWk
/
Cata l;i « IcOPi ill «xn alluci 1
?., »¡at
#ivr DNA (!¡minse 4 ?.?,?? nrdectaiç>ifi-*le
\
14 16 Otbcr 2 TcO,2VIi-IIlI \iu... Vi»E n»auttì4*t'*
Ir·.;, ? RSH 17/i WihD I
Or*" Te
6\
Peróxido <*n<I RSS 10reactive aWfhvd« GSSG INADP
E
uo>^ ZwI SoxRS CAT OwR 20 Í. valine I)¿2
Tè V 11 ( ^KH.O + O L-VfA V J NcS-* Cys '-^j O.VS ? H1S
FlWt \ S Sulftie
? a r:w e
thiE s ! uAf rcdiiclaw
{•¡mmenx
i>the S ??
ea/vnu:
II*lN*r-»> *i*s IiBtMR
íífcieen ill
H1O2+ i O,-H-O2+ 11· lïs-si
C"?
(CH.) -y»
Oll (CHi)1Tt
Hjtlruvvì radle*!
Figure 4: Modèle du rôle de certaines enzymes (et de certains de leurs produits) de la voie métabolique de la cysteine en réponse à la présence de K2Te03. Cys, cysteine; CysK, cysteine synthase; [Fe-S], noyau Fer-soufre; GSH and GSSG, glutathion réduit et oxydé, respectivement; IscS, cysteine désulfurasse; OAS, O-acetyl-L-serine; RSH and RSSR, thiols réduits et oxydés, respectivement; SOD, superoxyde dismutase; SUMT, S-adénosyl-L-méthionine uroporphirynogen III C-méthyltransférase; YqhD, aldéhyde reductase; ZWF, glucose-6-phosphate déshydrogénase; ACN, aconitase; FUM, fumarase; LpdA, E3 composante de la pyruvate déshydrogénase; YggE, protéine antioxydante; UbiE, C-méthyl transferase; SoxRS, régulon du stress oxydatif (sensible aux anions superoxydes); OxyR, régulon du stress oxydatif (sensible au peroxyde d'hydrogène). Cette figure est tirée de
Ceci génère des anions superoxydes (O2") augmentant du même coup les dérivés réactifs de
l'oxygène (DRO) déclenchant le stress oxydatif et les dommages cellulaires s'y rapportant. La présence d'anions superoxydes endommage les noyaux [Fe-S] des protéines et enzymes (acotinase et fumarase). La relâche de Fe peut générer des radicaux hydroxyles ("OH) via la
réaction de Fenton ou Habèr-Weiss et ces radicaux causent des dommages macromoléculaires,
spécialement à l'ADN. L'lscS desulfurase participe à la restauration des noyaux [Fe-S]. La
réduction du K2TeO32" abaisse également le niveau de thiols réduits, alors récupérés par
l'oxydation du NADPH en NADP. CysK contribue à la restauration du pool intracellulaire de
thiols réduits et SUMT participe à la biosynthèse du groupe sirohème de la sulfite reductase.
Le Te0 peut être subséquemment éliminé sous sa forme alkylée, produite par la
méthyltransférase UbiE. La présence d'anions superoxydes peut aussi induire la peroxydation de la membrane lipidique et l'oxydation de protéines. YqhD détoxifie la cellule des aldéhydes réactifs dérivés de la peroxydation de la. membrane lipidique et YggE abaisse le niveau d'anions superoxydes. L'augmentation de superoxyde dismutase (SOD) permet la superoxyde dismutation et le peroxyde d'hydrogène généré est ensuite décomposé par la catalase. Le stress
induit par le K2TeO32" induit l'expression du régulon SoxRS. Zwf est induit par ce régulon et
permet la restauration du pool de NADPH. En somme, l'intégrité des ribosomes, des protéines et la synthèse de l'ADN sont affectées et la membrane plasmique subit des dommages. Le résultat final est la mort rapide de la cellule dans les 15 min suivant l'exposition au tellurite (Turner, 2001; Chasteen et al, 2009). Une vision émergente concernant la toxicité du tellurite stipule que les bactéries font face à la toxicité de ce composé par un mécanisme d'adaptation générale semblable à celui en réponse aux radiations UV et aux chocs thermiques (Chasteen et
al, 2009).
Summers et Jacoby (1977) ont été les premiers à corréler la résistance au tellurite de certaines souches bactériennes à Gram négatif à la présence de plasmides transférables. Tous les
Plasmides du groupe IncHI2 testés chez le Pseudomonas aeruginosa et quatre plasmides du
tellurite de potassium. L'appellation ter a alors été donnée au phénotype de résistance au
tellurite associé avec ces plasmides. Des années plus tard, Jobling et Ritchie (1987) ont pour la
première fois associé une région précise du plasmide conjugatif pMER610 de Alcaligenes au
phénotype de résistance au tellurite. Cette même équipe a ensuite publié la séquence
nucléotidique de la région complète permettant la résistance, qui s'est avérée contenir cinq
cadres de lecture ouverts, nommés ORF 1-5 (Jobling et Ritchie, 1988). Ces gènes ont par la
suite été désignés terA, terB, terC, terO et terE. Ils codent pour des polypeptides d'une masse
moléculaire prédite de 37, 14, 38, 20 et 20 kDa respectivement (Hill et al., 1993). Une région
du plasmide IncHI2 R478, isolé originellement de S. marcescens, permet une résistance au
tellurite (Ter), de même qu'une résistance à la colicine B (PacB) et une inhibition de la
propagation du phage T5 (PhiT5) chez IeE. coli (Whelan et al., 1995). Cette région contient
sept cadres de lecture ouverts dont les gènes ont été désignés terZ, -A, -B, -C, -E, et -F. Les
séquences des protéines TerA, B, C, D et E du plasmide R478 sont très similaires à celles du
plasmide pMERólO et les séquences de terZ et terF sont également présente dans pMERólO
(Whelan et al., 1995). L'insertion de transposon dans terO, terC et terZ réduit ou abolit
l'ensemble des phénotypes, alors que celles dans terE et ter? n'ont pas d'effet. L'insertion
dans terA réduit l'inhibition de la propagation de phage seulement (Whelan et al., 1995). Les
séquences de terO et terE étant très similaires, et il a été proposé qu'elles résultent d'une
duplication de gène. La portion N-terminale de la protéine TerA possède également certaines
similarités avec celles de TerD et TerE. Ces deux dernières seraient par ailleurs
interchangeables et il a été proposé que ces deux protéines forment un hétérodimère (Hill et
al., 1993; Whelan et al., 1995).
Les protéines codées par les gènes ter ne présentent pas de similarité avec des protéines
connues, mis à part TerD et TerE qui possèdent entre 43 et 51 % d'identité avec des protéines
liant l'AMPc de Dictyostelium discoideum, présentes chacune sous deux isoformes (CABP-IA, CABP-IB, CABP-2A, CABP-2B). Les protéines TerA, TerF, TerX et TerZ présentent
aussi une similarité, mais beaucoup plus faible. D. discoideum est un organisme eucaryote qui
croît d'abord de manière végétative sous forme d'amibes unicellulaires. Un manque de
nutriments induit subséquemment un processus de développement et de différenciation menant
ultimement à la formation d'un organe de fructification multicellulaire formé de spores et de
cellules porteuses « stalk cells ». L'AMPc joue un rôle de premier plan dans ce processus de
développement. Il agit comme chimioattractant pour diriger l'agrégation des amibes
individuelles lorsque le développement s'amorce, il régule la différenciation des spores et des
cellules porteuses et il contrôle l'expression de nombreux gènes impliqués dans le
développement (Bain et al, 1991). Des souches mutantes de D. discoideum ont été générées
par Bain et Tsang (1991). L'interruption d'un gène codant pour deux polypetides (p34 et p31)
reliés à CABP-I, car reconnu par des anticorps dirigés contre CABP-I, altère la vitesse de
croissance et le profil de développement de D. discoideum, mais seulement lorsque des
bactéries servent de nourriture (Bain et Tsang, 1991). CABP-I formée de deux sous-unités
(CABP-IA et CABP-IB) ainsi que p31 et p34 sont induites durant le développement (Kay et
al, 1987). Aucune étude récente n'a cependant poussé plus loin ni l'étude de CABP, ni celles
des polypeptides p34 et p31. L'implication de ces protéines dans le développement de
D. discoideum reste nébuleuse.
Quelques études ont été réalisées sur la régulation de l'opéron ter. Celui-ci semble être
exprimé de manière constitutive, mais aussi parfois de manière inductible selon la souche ou
l'espèce (Zannoni et al, 2008). En plus d'être présents sur tous les plasmides de type IncH12
(sauf R476b) et tous les plasmides IncHII isolés de différentes bactéries hôtes (Hou et Taylor,
1994; Taylor, 1999), des gènes hautement similaires à /erZABCDEF sont présents dans le
chromosome de plusieurs espèces dont le E. coli 0157:H7, le Yersinia pestis et le genre
Proteus (Taylor et al, 2002; Toptchieva et al, 2003). Le séquençage du génome de la souche
E. coli 0157 :H7 EDL933 (Perna et al, 2001) a révélé la présence de plusieurs grands îlots de
gènes (O-islands ou OI) absents chez la souche de référence E. coli K12. Parmi ceux-ci, deux
îlots identiques (OI#43 et OI#48) de 87 547 pb contiennent 96 cadres de lecture ouverts
codant entre autres pour une intégrase, des protéines phagiques, des protéines de résistance au
tellurite, une urease et une adhésine. Il existe une grande plasticité dans la présence, la
localisation et l'expression des ter OI chez le E. coli 0157 :H7 et il ne semble pas avoir de
relation entre la CMI du Te et la présence d'une ou deux copies des gènes ter (Taylor et al, 2002). La figure 5 compare les regroupements de gènes de résistance au tellurite ter provenant
de différentes espèces.
Plusieurs protéines de résistance au tellurite possèdent un ou deux domaines TerD dont le numéro d'accession pfam est le PF02342 (voir les informations disponibles sur le site http://pfam.sanger.ac.uk/family/terD). C'est le cas des protéines TerA, TerD, TerE, TerF, TerX et TerZ. Ce domaine est plus souvent retrouvé dans la portion C-terminale de la protéine et comporte 128 acides aminés. Il a été jusqu'à présent retrouvé dans 999 séquences de protéines provenant de 97 espèces, la plupart étant des bactéries. Dans plusieurs de ces espèces, plus d'une séquence de protéines possède le domaine TerD. Ce nombre atteint 17 séquences chez le S. coelicolor, mais dans l'ensemble il se situe plutôt autour de 4 à 5 séquences par génome. Des agents pathogènes comme le Yersinia pestis et le E. coli Ol57 :H7, des bactéries interagissant avec les plantes comme le Pseudomonas syringae et le Frankia de même que des bactéries sporulantes comme le Bacillus, le Clostridium et le Streptomyces comptent à elles seules pour 565 des séquences comportant un domaine TerD séquencées à ce jour. La plupart de ces protéines n'ont pas de fonction connue et sont présumées être des protéines de résistance au tellurite sur la base de leur similarité avec la protéine TerD des plasmides IncHI2. Dans plusieurs cas, les gènes codant pour ces protéines sont chromosomiques et ne sont pas organisés en opéron comme c'est le cas sur les plasmides
CMI EDL933 R 478 E.coli 0157:H7 (Strain 86-24""') pMJ606 PTE53 Proteus Deínococajs mdiadtimns TerZ Í>C TerZ 96.6%D€ TerZ
TerA TerB TerC TerD TerE TerF
192aa 191aa 103aa
TerA TerB
z>tzx --->rTerC TerD>cTerE TerF >
99.8% 994%
TerB
TipO TIrC TIrB TIrA
97.2%* 99.8% 99.5% 99.0%
TerC TerD TerE
75.2% 75.9% 3,2% J^-H J^L >
TerC TerA TerB
Dl X.
98,4%* 99.1% 99.4·»
TerA TerB TerC
=>l XZ3. 61,7% ?d.3% 80.9% 874% 82.9% 86.5% TerD TerE >c=x > 96.4% 969% TerD TerE >CZ>[ > ?d.3% TerA TerB <S00oc >j > 37% 32% 73,2% 67,5% TerD > 67»;
64-128 µ§ ml"1
(Taylor e/ al., 2002)1024 µ§ ml"'
(Taylor, 1999)1024 µ£ ml'1
(Hou et Taylor, 1994)64-1024 µd ml"1
(Taylor étal, 2002)> 250 µg ml"1
(Toptchieva et al., 2003)1024 µd ml"1
(Taylor et al, 2002)Figure 5: Comparaison des regroupements de gènes de résistance au tellurite ter selon les
différentes sources. Les cadres de lecture ouverts déduits sont montrés ainsi que le nombre
d'acides aminés pour les protéines prédites du E coli 0157 :H7 EDL933 sous les flèches
noires (Perna et al., 2001). Les protéines prédites du plasmide R478 du Serratia marscescens
sont les plus étroitement reliées à celles d'EDL933. Le pourcentage d'identité de chacun des
cadres de lecture ouverts prédits avec celui correspondant chez EDL933 est mentionné. TIpD,
TIrC, -B et A du £ coli 0157 : H7 souche 86-24NalR ont été nommés ainsi par Tarr et al
(2000). Le plasmide pMJ606 contient les gènes de résistance au tellurite clones à partir du
plasmide pMER610 isolé d'Alcaligenes sp. (Jobling et Ritchie, 1988), Le plasmide pTE53 provient d'une souche clinique du E. coli (Burian et al., 1998). Les deux dernières séquences
proviennent des bases de données du séquençage des génomes de Proteus sp. et de
Le tellurite étant retrouvé rarement dans l'environnement, la probabilité pour un organisme
d'y être en contact est plutôt faible.. Par contre, la découverte de nouvelles applications
industrielles pour ce métalloïde, au cours des dernières années, a fait augmenter le risque
potentiel. L'émergence de nombreux gènes codant pour une protéine possédant un domaine
TerD et ce, chez un grand nombre d'espèces bactériennes, incluant des agents pathogènes
humains, suggère que ces gènes procurent un avantage sélectif en environnements naturels,
qui pourrait ne pas être relié au phénotype de résistance au tellurite. Découlant de ceci, il a été
proposé que la résistance au tellurite soit un effet secondaire d'une autre fonction métabolique
particulière (Taylor, 1999).
Les gènes homologues à terD du S. coelicolor