© Charleen Salesse, 2017
Mécanismes développementaux des circuits
dopaminergiques et leur implication dans les
comportements hyperactifs
Thèse
Charleen Salesse
Doctorat en neurobiologie
Philosophiæ doctor (Ph. D.)
Québec, Canada
MÉCANISMES DÉVELOPPEMENTAUX DES CIRCUITS
DOPAMINERGIQUES ET LEUR IMPLICATION DANS LES
COMPORTEMENTS HYPERACTIFS
Thèse
CHARLEEN SALESSE
Sous la direction de :
Paul De Koninck, directeur de recherche
Martin Lévesque, codirecteur de recherche
iii
Résumé
Les neurones dopaminergiques du mésencéphale (mDA) sont impliqués de manière critique dans diverses fonctions clés du cerveau, y compris les mouvements volontaires, la récompense, l'attention et l'apprentissage. La bonne spécification des neurones dopaminergique, ainsi que l’établissement des circuits dopaminergiques sont nécessaires à un bon fonctionnement du cerveau. Le dysfonctionnement des circuits dopaminergiques est lié au développement de troubles neuropsychiatriques, y compris le trouble déficitaire de l'attention avec hyperactivité (TDAH), le trouble obsessionnel compulsif (TOC) et les troubles liés aux TOCs, comme le syndrome de Gilles de la Tourette.
L’obtention d’un circuit dopaminergique fonctionnel dépend du développement des neurones dopaminergiques. Les facteurs de transcription Lmx1a et Lmx1b font partie de la famille des LIM à homeodomain et sont des déterminants précoces de l’avenir des neurones dopaminergique. Lmx1a/b sont essentiels pour chaque étape de la différenciation des progéniteurs de neurone dopaminergique. Il a été démontré précédemment que les souris Lmx1a/b cKO ont une activité locomotrice augmentée par rapport aux contrôles. Ici, une caractérisation approfondie des souris Lmx1a/b a révélé que ces souris avaient un comportement hyperactif, en lien avec le TDAH, et démontraient des symptômes du type TOC. Au niveau cellulaire, la perte de fonction de Lmx1a/b a induit une réduction de l’arborisation dendritique et de la fréquence des courants postsynaptiques excitateurs miniatures spontanés (mEPSCs) dans les neurones dopaminergiques. Le profil d'expression des gènes chez les souris Lmx1a / b cKO a révélé que Lmx1a/b contrôle l'expression de Slitrk2 et Slitrk5, deux membres de la famille des protéines Slit et Trk (Slitrk). Le gain et la perte de fonction de Slitrk2 et Slitrk5 dans des cultures de neurones dopaminergiques ont montré que Slitrk2 régule positivement et Slitrk5 régulent négativement la croissance dendritique. Également, le gain et la perte de fonction de Slitrk2 ont induit une variation de la densité des punctas synaptiques excitateurs (PSD95 et VGLUT). En conséquence, la perte de fonction de Slitrk2 a réduit la fréquence des mEPSCs, tandis que l'augmentation de l'expression de Slitrk2 a augmenté la fréquence des mEPSCs, sans changement d'amplitude ou dans la fréquence ou de l'amplitude des mIPSCs. Ces données suggèrent un rôle pour Slitrk2 dans la formation de synapses excitatrices fonctionnelles. À l'inverse, le gain et la perte de fonction de Slitrk5 ont induit une modification de la densité des punctas synaptiques inhibiteurs (géphyrine et VGAT). La perte
iv
d’expression de Slitrk5 a réduit la fréquence des mIPSCs tandis que l'augmentation de l'expression de Slitrk5 a augmenté la fréquence des mIPSCs, sans changement dans l'amplitude ou de la fréquence et de l'amplitude des mEPSCs. Ces données suggèrent un rôle pour Slitrk5 dans la formation de synapses fonctionnelles inhibitrices. Nous avons également étudié les conséquences sur le comportement de Slitrk2 et Slitrk5 dans les neurones mDA. Les souris, dans lesquelles Slitrk2 a été invalidé dans la VTA, démontrent un changement significatif dans l'activité locomotrice et montrent de l’hyperactivité. À l'inverse, les souris avec une expression réduite de Slitrk5 présentent une activité locomotrice réduite et un comportement analogue à un TOC.
Ces changements de comportement peuvent être causés par une modification de l'activité des neurones dopaminergiques. L'inhibition chronique des neurones de la VTA, en utilisant une approche pharmacogénétique, pendant le développement postnatal à induit une activité motrice augmentée, similaire au TDAH, et un comportement analogue à un TOC. Ceci évoque certains aspects du comportement des souris Lmx1a/b cKO. Une inhibition aiguë a entraîné une diminution de l'activité locomotrice, alors que l'inhibition chronique chez des animaux plus âgés n'a eu aucun effet. Ensemble, ces résultats indiquent que Lmx1a/b, Slitrk2, et Slitrk5 sont des acteurs clés du développement des neurones dopaminergique et de la formation des synapses, ce qui peut avoir un impact sur le développement de TDAH et de TOC.
v
Abstract
Midbrain dopaminergic (mDA) neurons are critically involved in various key functions of the brain, including voluntary movement, reward, attention, and learning. The proper specification of dopaminergic neurons, as well as the establishment of dopaminergic circuits are necessary to a good functioning of the brain. Dopaminergic circuitry dysfunctions are linked to the development of neuropsychiatric disorders, including attention deficit hyperactivity disorder (ADHD), obsessive-compulsive disorder (OCD) and OCD-like disorders, such as Gilles de la Tourette’s syndrome.
The LIM-homeodomain transcriptional factors Lmx1a and Lmx1b are early determinants of the dopaminergic fate and are essential for each step of mDA progenitor differentiation. Previously, it has been demonstrated that Lmx1a/b cKO mice show increased locomotor activity. Further characterization of Lmx1a/b cKO mice revealed that these mice had ADHD- and OCD-like behaviour. The loss of function of Lmx1a/b reduced dendritic morphology and frequency of spontaneous miniature excitatory postsynaptic currents (mEPSCs) in mDA neurons. Gene expression profiling in Lmx1a/b cKO mice revealed that Lmx1a/b controls the expression of Slitrk2 and Slitrk5, two members of the Slit and Trk-like (Slitrk) protein family.
Gain and loss of function of Slitrk2 and Slitrk5 in mDA neuron cultures showed that Slitrk2 positively regulates and Slitrk5 negatively regulate dendritic growth. Additionally, gain and loss of function of Slitrk2 induced a change in the density of excitatory synaptic puncta (PSD95 and VGLUT). Accordingly, Slitrk2 knockdown reduced the frequency of mEPSCs while increased Slitrk2 expression increased the frequency of mEPSCs, with no change in amplitude or in mIPSCs frequency or amplitude. These data suggest a role for Slitrk2 in the formation of functional excitatory synapses. Inversely, gain and loss of function of Slitrk5 induced a modification in the density of inhibitory synaptic puncta (gephyrin and VGAT). Slitrk5 knockdown reduced the frequency of mIPSCs while increased Slitrk5 expression increased the frequency of mIPSCs, with no change in amplitude or in mEPSCs frequency or amplitude. These data suggest a role for Slitrk5 in the formation of functional inhibitory synapses. We also investigated the consequences on behaviour of Slitrk2 and Slitrk5 reduced expression in mDA neurons. Mice, in which Slitrk2 was knocked down in the VTA, display significant change in locomotor activity and show ADHD. Inversely, mice with reduced expression of Slitrk5 exhibit lower activity and OCD-like behaviour.
vi
These behavioural changes might be caused by a change in mDA neuron firing activity. Chronic inhibition of mDA neurons during postnatal development using a pharmacogenetic approach induced ADHD and OCD-like behaviour and mimic some aspects of the Lmx1a/b cKO mice. Acute inhibition resulted in decreased locomotor activity, while chronic inhibition in older animals had no effect. Altogether, these results indicate that Lmx1a/b and Slitrk2/5 are key players of mDA neuron development and synapse formation, which may have an impact on ADHD and OCD-like disorders.
vii
Table of contents
Résumé... iii
Abstract ... v
Table of contents ... vii
List of figures ... xiii
List of tables ... xiv
List of abbreviations ... xv
Remerciements ... xviii
Preface ... xxi
1. Introduction ... 1
1.1. The dopamine system ... 1
1.1.1. The substancia nigra pars compacta ... 2
1.1.2. The ventral tegmental area ... 2
1.2. Dopamine... 3
1.2.1. Roles and functions... 3
1.2.2. The dopamine transporter ... 4
1.2.3. Receptors ... 4
1.2.3.1. D1-like receptors ... 6
1.2.3.2. D2-like receptors ... 6
1.2.3.3. Expression of dopamine receptors ... 7
1.3. Main dopaminergic pathways ... 7
1.3.1. Nigrostriatal pathway ... 8
1.3.2. Mesolimbic pathway ... 8
1.3.3. Mesocortical pathway ... 8
1.3.4. Inputs on dopaminergic neurons ... 8
1.3.4.1. Inputs to the VTA ... 8
1.3.4.2. Inputs to the SNpc ... 10
1.4. Dopaminergic neurons ... 11
1.4.1. Origin and development ... 11
1.4.2. Transcription factors of mDA ... 13
1.4.2.1. Otx2... 14 1.4.2.2. En1/2 ... 14 1.4.2.3. FOXA1/2 ... 14 1.4.2.4. Lmx1a/b... 14 1.4.2.5. Msx1/2 ... 16 1.4.2.6. Mash1 and Ngn2 ... 16 1.4.2.7. Nurr1 ... 16
1.4.3. Heterogeneity in neurotransmitters in mDA neurons ... 16
1.4.3.1. Dopaminergic neurons co-release glutamate... 17
1.4.3.2. Dopaminergic neurons co-release GABA ... 17
viii
1.4.4.1. Electrophysiological properties of dopaminergic neurons of the SNpc... 18
1.4.4.2. Electrophysiological properties of dopaminergic neurons of the VTA ... 18
1.4.5. Morphology of dopaminergic neurons ... 18
1.4.6. Synapses and dopamine ... 19
1.5. Synaptic adhesion molecules ... 20
1.5.1. Adhesion domains ... 20
1.5.2. Neuroligins and Neurexins ... 21
1.5.3. Cadherins ... 21
1.5.4. SynCAM... 21
1.5.5. Nectin ... 22
1.5.6. Receptor proteine-tyrosine phosphatases ... 22
1.5.7. Tropomyosin receptor kinase ... 22
1.5.8. Leucine-rich repeat proteins ... 22
1.5.9. Slitrk proteins ... 23
1.5.9.1. Distribution and expression ... 23
1.5.9.2. Protein structure ... 27 1.5.9.3. Slitrk1 ... 28 1.5.9.4. Slitrk2 ... 29 1.5.9.5. Slitrk3 ... 30 1.5.9.6. Slitrk4 ... 30 1.5.9.7. Slitrk5 ... 30 1.5.9.8. Slitrk6 ... 31
1.6. Attention deficit/hyperactivity disorder ... 32
1.6.1. Dopamine and ADHD ... 32
1.6.2. Treatment of ADHD ... 33
1.6.3. Genetics of ADHD ... 33
1.6.4. Brain circuits and regions implicated in ADHD ... 33
1.6.5. Animal models of ADHD ... 34
1.6.5.1. Genetic models of ADHD ... 34
1.6.5.2. Induced animal models of ADHD... 35
1.6.6. Comorbitity with ADHD ... 36
1.6.6.1. Obsessive-compulsive disorders ... 36
1.6.6.2. Tourette’s syndrome ... 37
1.6.6.3. Bipolar disorder... 37
1.6.6.4. Schizophrenia ... 37
1.7. Aim of the project ... 38
1.7.1. Objectives ... 39
1.7.1.1. Characterize the behavioural and physiological phenotypes associated with Lmx1a/b inactivation in midbrain dopaminergic neurons. ... 39
1.7.1.2. Determine the roles of Slitrk2 and Slitrk5 in midbrain dopaminergic neurons dendritic growth and synapse formation. ... 39
ix
1.7.1.3. Determine the in vivo roles of Slitrk2 and Slitrk5 in midbrain dopaminergic circuit
formation and related behaviour... 39
1.7.1.4. Determine the role of synaptic input on dopaminergic neurons and circuit formation in hyperactivity related behaviours ... 40
1.7.2. Importance of this work ... 40
2. Opposite control of excitatory and inhibitory synapse formation by Slitrk2 and Slitrk5 on dopaminergic neurons modulates hyperactivity ... 41
2.1. Résumé ... 41
2.2. Summary... 42
2.3. Significance statement ... 42
2.4. Introduction ... 43
2.5. Results ... 44
2.5.1. Loss of Function of Lmx1a/b Leads to Hyperactive Behaviour, Altered Dendritic Arborization and Altered Synaptic Input on mDA Neurons ... 44
2.5.2. Members of the Slitrk Family Are Regulated by Lmx1a/b and Are Expressed in mDA Neurons ... 47
2.5.3. Slitrk2 and Slitrk5 Have Opposite Effects on the Regulation of mDA Neuron Dendritic Morphology... 50
2.5.4. Slitrk2 and Slitrk5 Respectively Promote Excitatory and Inhibitory mDA Synaptic Inputs on mDA Neurons ... 52
2.5.5. Loss of Function of Slitrk2 in the VTA Leads to Hyperactivity ... 54
2.5.6. Chronic Pharmacogenetic Inhibition of mDA Neurons During Development Leads to Hyperactivity ... 56
2.6. Discussion ... 59
2.7. Materials and Methods ... 61
2.7.1. Mouse Lines... 61
2.7.2. qRT-PCR Analysis ... 61
2.7.3. Multiplex Fluorescent In Situ Hybridization... 62
2.7.4. Primary Cell Cultures of mDA Neurons and Plasmid Transfections ... 62
2.7.5. Immunofluorescence ... 63
2.7.6. Sholl and Synaptic Density Analyses ... 64
2.7.7. Electrophysiological Recordings ... 64
2.7.8. Neonatal Stereotaxic Brain Injections ... 65
2.7.9. Behavioural Analysis ... 65
2.7.10. Pharmacogenetic Inhibition of mDA Neurons in the VTA ... 66
2.7.11. Statistical Analysis ... 66 2.8. Acknowledgments ... 66 2.9. Author Contributions ... 66 2.10. References ... 67 2.11. Supplementary Figures ... 73 3. Discussion ... 84
3.1. Lmx1a/b cKO, hyperactivity, and OCD ... 84
x
3.3. Input on dopaminergic neurons... 86
3.4. Lmx1a/b regulate the expression of Slitrk2 and Slitrk5 ... 88
3.5. Polarizing roles for Slitrk2 and Slitrk5 in dendritic growth... 88
3.6. What about the axon? ... 89
3.7. Slitrk2 at excitatory synapses in dopaminergic neurons ... 90
3.8. Slitrk5 at inhibitory synapses in dopaminergic neurons ... 91
3.9. Importance of excitatory-inhibitory balance ... 92
3.10. Activity-dependent dendritic growth ... 93
3.11. What about other members of the Slitrk family and dopaminergic neurons? ... 95
3.12. Slitrk2 as a potential marker for hyperactivity ... 96
3.13. Role of the VTA in hyperactivity ... 96
3.14. What about the role of the SNpc in hyperactivity? ... 98
3.15. Slitrk5 and OCD ... 98
3.16. Dopamine and OCD ... 98
4. Conclusion ... 100
5. Bibliography ... 102
6. Appendix 1: Gold Nanoparticle-Assisted all optical localized stimulation and monitoring of Ca2+ signalling in neurons... 128
6.1. Abstract ... 128
6.2. Introduction ... 128
6.3. Results ... 129
6.3.1. Nanoparticle-assisted localized optical stimulation with bare gold nanoparticles ... 129
6.3.2. NALOS with functionalized gold nanoparticles ... 130
6.3.3. Electrophysiological recordings during NALOS ... 131
6.3.4. NALOS to investigate localized Ca2+ signaling ... 132
6.4. Discussion ... 132
6.5. Methods ... 134
6.5.1. Rat hippocampal cultures, plasmids, and transfection ... 134
6.5.2. Confocal imaging and laser-induced stimulation ... 134
6.5.3. Functionalization of the gold nanoparticles ... 135
6.5.4. Gold nanoparticle incubation ... 135
6.5.5. Electrophysiological recording ... 135
6.5.6. Calcium imaging analysis ... 135
6.6. References ... 136
6.7. Figures ... 138
7. Appendix 2: βCaMKII is necessary for the activity-dependent trapping of AMPA receptors at synapses ... 142
7.1. Abstract ... 142
7.2. Significance Statement ... 143
7.3. Introduction ... 143
7.4. Materials and Methods ... 144
xi
7.4.2. Live imaging and rapid quantum dot labeling ... 144
7.4.3. Single molecule analysis ... 145
7.4.4. Fixation and immunocytochemistry ... 145
7.4.5. Confocal imaging ... 145
7.4.6. Electrophysiology ... 145
7.4.7. Experimental Design and Statistical Analysis ... 145
7.5. Results ... 146
7.5.1. βCaMKII cannot substitute for αCaMKII in trapping AMPA receptors at synapses under basal neuronal activity... 146
7.5.2. βCaMKII is required for the immobilization of AMPARs at synapses after cLTP induction ... 146
7.5.3. shRNA knockdown of βCaMKII decreases the frequency of mEPSCs... 147
7.6. Discussion ... 147
7.7. Reference ... 149
7.8. Figures : ... 151
8. Appendix 3: Role of calpain in synaptic plasticity. ... 159
8.1. Abstract ... 159
8.2. Figure... 160
9. Appendix 4: Transcriptional repression of Plxnc1 by Lmx1a and Lmx1b direct topographic dopaminergic circuit formation ... 161
9.1. Abstract ... 161
9.2. Introduction ... 162
9.3. Results ... 162
9.3.1. Lmx1a/b are required for appropriate mDA axon projections ... 162
9.3.2. Lmx1a and Lmx1b regulate Plxnc1 expression ... 164
9.3.3. Sema7a/Plxnc1 interaction mediates repulsion of VTA neurons ... 164
9.3.4. mDA projections in the striatum is altered in Sema7a KO mice ... 165
9.3.5. Forced expression of Plxnc1 in mDA neurons recapitulates the Lmx1a/b cKO phenotype ... 165
9.3.6. Otx2 and Lmx1a/b regulation of Plxnc1 expression in mDA neurons ... 166
9.4. Discussion ... 166
9.5. Methods ... 168
9.5.1. Animals ... 168
9.5.2. Tissue analysis ... 169
9.5.3. Optical density measurements on sections ... 169
9.5.4. In situ hybridization ... 169
9.5.5. Quick TH staining and LCM ... 169
9.5.6. cDNA library production... 170
9.5.7. RNA sequencing ... 170
9.5.8. RT-qPCR ... 170
9.5.9. Brain injections ... 171
xii
9.5.11. Western blot... 171
9.5.12. In vitro mDA neurons and explant cultures ... 171
9.5.13. Stripe assay ... 172 9.5.14. iDISCO ... 172 9.5.15. Light-sheet imaging ... 172 9.5.16. Electrophysiological recordings ... 173 9.5.17. Statistical analysis ... 173 9.5.18. Microscopes... 173 9.5.19. Data availability ... 173 9.6. Acknowledgements ... 174 9.7. Author Contributions ... 174 9.8. Figures ... 174 9.9. References ... 189
10. Appendix 5: MT3-MMP promotes excitatory synapse formation by promoting Nogo-66 receptor ectodomain shedding ... 192
10.1. Abstract ... 192
10.2. Significance ... 192
10.3. Results ... 192
11. Appendix 6: Fast two-photon imaging of subcellular voltage dynamics in neuronal tissue with genetically encoded indicators ... 194
11.1. Abstract ... 194
12. Appendix 7: Optical approach for the measurement of electrical activity in neurons with voltage-sensitive dye ANNINE-6plus ... 195
12.1. Abstract ... 195
12.2. Introduction ... 195
12.3. Methods ... 196
12.3.1. Slice preparation ... 196
12.3.2. Electrophysiology and Annine-6plus labeling ... 197
12.3.3. Optophysiological recordings ... 197
12.3.4. Data analysis ... 197
12.4. Results ... 198
12.4.1. Single cell labelling and imaging ... 198
12.4.2. Evaluation of phototoxicity ... 198
12.4.3. Measurement of electrical activity in rat hippocampal slices ... 200
12.4.4. Synaptic events recordings ... 201
12.5. Discussion ... 202
xiii List of figures
Figure 1-1: Midbrain section in adult mice. ... 2
Figure 1-2: Biosynthesis pathway for dopamine. ... 3
Figure 1-3: Schematic representation of dopaminergic synapse ... 4
Figure 1-4: Schematic of dopaminergic receptors ... 5
Figure 1-5: Dopaminergic pathways ... 7
Figure 1-6: Inputs to ventral tegmental area neurons ... 9
Figure 1-7: Neurulation in the mammalian embryo ... 11
Figure 1-8: Transcriptional control of ventral midbrain dopaminergic neurons ... 13
Figure 1-9: Genomic organization of SLITRKs ... 24
Figure 1-10: Structure Homology of Slitrk proteins to Trk and Slit ... 27
Figure 1-11: Structure of Slitrk proteins ... 28
Figure 2-1: Loss of Lmx1a/b results in altered motor function, dendritic growth and synaptic input. 46 Figure 2-2: Lmx1a/b transcription factors regulate Slitrk2 and Slitrk5 expression in mDA neurons. 49 Figure 2-3: Slitrk2 and Slitrk5 differentially regulate dendritic development. ... 51
Figure 2-4: Slitrk2 and Slitrk5 have opposite effects on the formation of inhibitory and excitatory synapses. ... 53
Figure 2-5: Knockdown of Slitrk2 and Slitrk5 in the VTA results in altered motor behaviour. ... 55
Figure 2-6: Selective inhibition of dopaminergic neurons in the VTA during development results in increased motor activity. ... 57
Figure 2-7 (Supplementary): Loss of Lmx1a/b results in altered motor function. ... 73
Figure 2-8 (Supplementary): Loss of Lmx1a/b results in altered dendritic arborization in acute slices. ... 74
Figure 2-9 (Supplementary): Slitrk2 and Slitrk5 have opposing roles at excitatory and inhibitory synapses. ... 75
Figure 2-10 (Supplementary): Knockdown of Slitrk2 or Slitrk5 with an alternate shRNA. ... 76
Figure 2-11 (Supplementary): shRNA constructs of Slitrk2 and Slitrk5 effectively reduce mRNA expression. ... 77
Figure 2-12 (Supplementary): Knockdown of Slitrk2 or Slitrk5 results in altered motor behaviours. 78 Figure 2-13 (Supplementary): Dat-IRES-Cre mice display normal motor behaviours compared to their control littermates. ... 79
Figure 2-14 (Supplementary): Saline injection in mice expressing an inhibitory DREADD does not alter motor behaviours. ... 80
Figure 2-15 (Supplementary): Hyperpolarization of cultured mDA neurons expressing an inhibitory DREADD in the presence of CNO. ... 81
Figure 2-16 (Supplementary): Persistence of hyperactivity after chronic CNO injections. ... 82
Figure 4-1: Lmx1a/b regulates Slitrk2 and Slitrk5 expression, which exert opposite roles in dendritic growth and synapse formation ... 101
xiv List of tables
Table 1-1 : Expression and distribution of Slitrk in the brain at different stages of development in mice ... 25
xv
List of abbreviations
ADHD: Attention deficit hyperactivity disorder
AMPAR: α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor Aldh1a: aldehyde dehydrogenase 1a1
bHLH: Basic helix-loop-helix family BNST: Bed nucleus of the stria terminalis Ca2+: Calcium
CaMKII: Ca2+/calmodulin-dependent protein kinase II
cLTP: Chemical long-term potentiation CNO: Clozapine-n-oxide
DA: Dopamine
DAT: Dopamine transporter
DREADD: designer Receptor Exclusively Activated by Designer Drugs DRN: Dorsal raphe nucleus
EPSC: Excitatory post synaptic current GABA: γ-aminobutyric acid
GAD: Glutamate decarboxylase
GAD67: Glutamate decarboxylase with amolecular weight of 67 kDa HCN: Hyperpolarizing cation current dependent on cyclic nucleotide IPSC: Inhibitory post synaptic current
L-DOPA: L-3,4-dihydroxyphenylalanine LDTg: Laterodorsal tegmentum
LHb: Lateral habenula LHT: Laterhal hypothalamus
LINGO: Leucine rich repeat and Immunoglobin-like domain-containing protein Lmx1: LIM homeobox transcription factor 1
LRR: Leucine-rich repeat LTP: Long term potentiation
MAP2: Microtubule-associated protein 2 Mash1: Mouse Achaete-Schute homolog 1 mDA: Midbrain dopaminergic neuron mEPSC: Miniature EPSC
xvi
MSN: medium spinal neurons
MT3-MMP: Membrane-type-3 matrix metalloproteinase mPFC: medium prefrontal cortex
NAc: Nucleus accumbens NGF: Neuronal growth factor NGL: Netrin-G ligands Ngn2: Meuroligin 2
NMDAR: N-methyl-D-aspartate receptor Nurr1: Nuclear receptor related 1 protein PAG: Periaqueductal grey
RMTg: Rostromedial mesopontine tegmental nucleus SAM: Synaptic adhesion molecules
SALM: Synaptic adhesion-like molecules Slitrk: SLIT and NTRK-like
SNpc: Substancia nigra pars compacta TH: Tyrosine hydroxylase
TTX: Tetrodotoxin
VGAT: Vesicular GABA transporter VGLUT: Vesicular glutamate transporter VMAT2: Vesicular monoamine transporter 2 VP: Ventral pallidum
xvii
A scientist in his laboratory is not a mere technician: he is also a child confronting natural phenomena that impress him as though they were fairy tales. -Marie Curie
Nothing is impossible, the word itself says I’m possible! -Audrey Hepburn
Above all, don't fear difficult moments. The best comes from them. -Rita Levi-Montalcini
xviii
Remerciements
Je tiens à adresser mes remerciements aux nombreuses personnes qui ont contribué, de près ou de loin, à la réalisation de cette thèse. Sans vous, je ne serai pas où j’en suis aujourd’hui.
Je tiens tout d’abord à remercier mon directeur thèse Paul De Koninck. Sans le savoir, ou peut-être le savais tu un peu, tu m’as ramassé à la petite cuillère. Après avoir passé un moment difficile, ce fut comme une bouffée de fraicheur d’arriver dans ton laboratoire. Je tiens à te remercier pour tout ton soutien, ta confiance et tes encouragements (parfois sous forme de « fist bump »). Je tiens également à te remercier pour tout le temps que tu m’as accordé, car parfois tu n’en avais pas, mais tu as toujours trouvé des moyens pour en trouver pour moi. Les multiples discussions, qui souvent te gardais à ton bureau jusqu’au soir, m’ont permis de m’épanouir en tant que scientifique : tu m’as montré à réfléchir. Merci d’avoir cru en moi.
Merci également à Martin Lévesque, mon codirecteur de recherche, de m’avoir accueilli dans ton laboratoire avec tes idées et projets de grandeur. Tu m’as poussé à me dépasser. Avec toi, j’ai pu développer mes compétences dans un grand nombre de nouvelles techniques. Avoir été dans ton laboratoire m’a permis de vivre des expériences vraiment enrichissantes à l’étranger, en allant présenter mes travaux dans divers congrès scientifiques. Je te remercie pour toutes les opportunités que tu m’as offertes.
J’aimerais remercier les membres du laboratoire de Paul, spécialement à Francine. Merci de m’avoir enseigné la culture cellulaire avec une rigueur incomparée. Tu m’as montré ce qu’est un travail bien fait et tu as réponse à tout. Un gros merci. Merci à Laurence, avec qui j’ai la chance de travailler dans le laboratoire, chaque jeudi. Avec ton incapacité de n’avoir rien à faire, tu m’as montrée ce qu’est être dédié à son travail. Merci à Simon, pour les multiples discussions, tant de science et que de potins, et de m’avoir fait confiance en m’incluant dans un projet qui te tenait à cœur. Merci à Flavie, pour nos collaborations passées et futures, pour tous tes conseilles, et pour être devenue une référence et une amie Merci à Olivier, pour m’avoir initié à l’imagerie. Merci également à Mado, Christian, Theresa, Kapil, Valérie, Gabriel et Pierre-Luc.
J’aimerais remercier les membres du laboratoire de Martin, spécialement à Véronique. La liste de choses que tu fais qui rend ma vie, et celle de tous les étudiants, plus facile est trop
xix
longue. Je vais donc te remercier pour tout ton aide et ton soutien. Reviens vite! Thank you Marishka for all your help, I’ll never forget, and the elevator is coming back your way! Julien, merci de m’avoir initié au projet (et de m’avoir présenté (sort of) Guillaume). Hélène, merci pour ta collaboration et les bons commentaires sur mon manuscrit. Merci à Audrey pour tout ton aide. Merci à Axelle pour ton éternelle bonne humeur. Merci également à Marcos, Cléo, Guillaume (voir plus bas), Catherine, François, Caroline et Sarah.
Ma première vraie expérience en recherche, en tant qu’étudiante au Bac en biologie à l’UQAC où j’ai travaillé dans le laboratoire de recherche, m’a permis de découvrir que j’aimais vraiment la recherche universitaire. Je tiens donc à remercier Catherine Laprise de m’avoir donné l’opportunité de mieux connaitre le milieu de la recherche. Les deux ans et demi passés dans ton labo m’ont montré que pour réussir, il faut se dépasser.
L’accomplissement d’un doctorat ne se fait pas du jour au lendemain, et on oublie parfait comment tout ça a commencé. Pour moi, c’est mon cours d’initiation à la recherche au Cégep qui a été l’élément déclencheur. Ce cours peut sembler anodin pour plusieurs, mais il a été mon premier contact avec la possibilité d’une carrière en recherche. Mes enseignants m’ont initié à cette carrière (et créer un monstre); ils nous ont laissés faire la recherche que l’on voulait, moi et mon équipe, tout en nous guidant et en nous montrant les bases de la recherche. Je tiens donc à remercier très sincèrement Martin Bertrand et Jean-François Lambert.
Merci à mes amis au centre. Alesya et Simon, vous avez été là dès mon arrivée au centre. Même si vous êtes partie (vers un monde meilleur?), je n’oublierai jamais les bons moments (et les mauvais) passés ensemble. Merci d’avoir été mes amis et d’avoir partagé avec moi certaines embuches. Avec Yanina et Dino, vous avez été mon laboratoire d’adoption, où vous m’avez donné le titre de membre honorifique. I think the fridge is still broken. Thanks Dino, for being a great friend, and for all the late nights and cocktails at HQ. Merci à tous les autres que je côtoie au quotidien (Cyril, Mario, Louis, Marc, Lou-Lou, Karine, Annie, et tous les autres, car je ne peux tous vous nommer). Merci au personnel de soutien du centre de recherche CERVO. Merci à Isabelle et Catherine de l’animalerie pour votre immense aide technique et sans qui toutes ces expériences ne pourraient avoir lieu. Merci à Sonya, pour tout ton soutient et ton aide.
xx
Merci à mes amis. Même si les occasions de se voir ont été rares, elles ont été appréciées. Merci spécialement à Marie-Ève, d’avoir été une amie hors pair depuis tant d’années.
Merci à ma famille. Merci à ma belle-sœur Mélanie, mon beau-frère Martin, mon « filleul » Alexandre et mes beaux-parents Louiselle et Marc, et ma belle-sœur Joanie pour votre soutien et vos encouragements.
Merci à mon frère Andrew, qui me rappelle souvent qu’il n’y a pas que l’école dans la vie et que je devrais passer moins de temps dans mes livres.
Merci à mes parents, Nancy et Pierre, d’avoir toujours cru en moi. De m’avoir montré que je suis capable de tout. De m’avoir encouragé, épaulé et poussé tous les jours, et ce, dès mes premiers à la petite école. La confiance en soi que vous m’avez apportée a fait de moi qui je suis aujourd’hui. Merci pour tout.
Guillaume (encore), ton soutien et tes encouragements m’ont permis de passer à travers les bons moments jusqu’aux plus difficiles. Tu as su comprendre mes embuches comme personne. Les mots ne suffisent pas pour te dire merci.
xxi
Preface
This thesis is part of the final steps in the doctoral program in neurobiology at Université Laval. I performed all my research in Dr Paul De Koninck’s and Dr Martin Lévesque’s labs in the CERVO Brain Research Center.
In my first chapter, I introduced my article. The second chapter is the core of this thesis. It is the main article I have worked on during my doctoral studies and has recently been submitted to Nature Medicine. For this, I contributed to the planning of the research and experimental design. I performed and analyzed electrophysiological recordings, stereotaxic injections, immunohistochemistry, cell cultures, behaviour experiments. I also I wrote the manuscript.
The next chapter discusses why this article is of great impact, and what are the next steps in this project. The last chapter is a conclusion to my thesis.
I have also included many appendices, which are some of the many collaborations I was fortunate to participate in.
List of contributions:
1. Lavoie-CardinalF, Salesse C, Bergeron E, Meunier M, and Koninck P. Gold
nanoparticle-assisted all optical localized stimulation and monitoring of Ca2+ signalling in neurons. Scientific Reports. 2016 Feb 09
2. Lavoie-Cardinal F, Salesse C, Ayotte-Nadeau PL and De Koninck P. Nanoparticle-assisted
localized optical stimulation of cultured neurons. Nanoparticles in neuroscience. F.
Santamaria and X. Peralta, Springer. In press.
3. Chamberland S, Yang HH, Pan M, Evans SW, Guan S, Chavarha M, Yang Y, Salesse C, Wu H, Wu JC, Clandinin TR, Toth K, Lin MZ, St-Pierre F. Fast two-photon imaging of
subcellular voltage dynamics in neuronal tissue with genetically encoded indicators. eLife.
In press.
4. Chabrat A, Brisson G, Doucet-Beaupré H, Salesse C, Schaan Profes M, Dovonou A, Akitegetse C, Charest J, Lemstra S, Côté D, Pasterkamp RJ, Abrudan MI, Metzakopian E, Ang SL and Lévesque M. Transcriptional repression of Plxnc1 by Lmx1a and Lmx1b directs
topographic dopaminergic circuit formation. Nature Communications. Accepted
5. Labrecque S, Salesse C, and De Koninck P. βCaMKII is necessary for the
xxii
6. Sanz R, Ferraro G, Kacervosky B, Salesse C, Gowing E, Beaubien F, Rambaldi I, Holmbeck K, Cloutier JF, Lévesque M, Murai K, and Fournier A. MT3-MMP promotes excitatory synapse formation by promoting Nogo-66 receptor ectodomain shedding. J Neuro. Under revision
7. Salesse C, Charest J, Doucet-Beaupré H, De Koninck P, and Levesque M. Slitrk2 and
Slitrk5 differentially control excitatory and inhibitory synapse formation on midbrain dopaminergic neurons and modulate hyperactivity-related behaviour. Nat Med. Submitted
8. Dupont-Therrien O*, Salesse C*, Labrecque S, Cote D, and De Koninck P.
Optophysiological recordings of neuronal and synaptic activities from single neurons in rat hippocampal slice with the voltage-sensitive dye Annine-6plus. In preparation (*equal
contribution)
My expertise in electrophysiology made me an ideal candidate to collaborate on many projects. The first project was in Paul De Koninck’s lab along with Michel Meunier from Polytechnique Montréal. This project aimed to characterize and validate gold nanoparticles as a new tool for optical stimulation of neurons. To do this, calcium imaging combined with whole-cell patch-clamp recordings were performed on cultured hippocampal neurons. The results of this project were published in Scientific Reports. I have included the article in Appendix 1: Gold Nanoparticle-Assisted all optical localized stimulation and monitoring of Ca2+ signalling in neurons. For this article, I planned, performed and analyzed the electrophysiological recordings, as well as helped Flavie Lavoie-Cardinal with writing part of the manuscript that delt with electrophysiological recordings. A book chapter, for which I contributed, was also written following the publication (in press).
Another project in Paul De Koninck’s lab aimed to define the role of βCaMKII in synaptic plasticity and compare it to αCaMKII. To do this, quantum dot imaging was used to image AMPAR mobility using a cLTP protocol in different overexpression/knockdown of α and βCaMKII. This needed to be physiologically investigated and this is where I was able to validate, using whole-cell patch-clamp recordings, the findings observed in imaging experiments. The results of this project were submitted to the Journal of Neuroscience. I have included the paper in Appendix 2: βCaMKII is necessary for the activity-dependent trapping of AMPA receptors at synapses. For this article, I performed and analyzed the electrophysiological recordings, as well helped Simon Labrecque with writing part of the manuscript that delt with my results. The role of calpain in synaptic plasticity is an additional project I participated in Paul’s lab. It is in collaboration with Kapil Seghal, a PhD student and Valérie Clavet-Fournier, a MSc student. For this project, I have, and will, confirm results
xxiii
obtained by imaging (Kapil) and biochemistry (Valerie). As this project is still in the works, I have not included a manuscript, but rather a figure and a brief explanation of my contribution so far in Appendix 3: Role of calpain in synaptic plasticity.
In Martin Lévesque's lab, my expertise allowed me to collaborate on a project about the role of PlexinC1 in axon guidance. In this project, we found that Lmx1a and Lmx1b control the expression of PlexinC1. We show that the interactions of PlexinC1 and its ligand, Semaphorin7a, are responsible for the segregation of nigrostriatal and mesolimbic dopaminergic pathways. For this project, I was asked to show the physiological impact of loss of dopamine innervation in the dorsal striatum. For this, I planned, performed and analyzed electrophysiological recordings in the ventral and dorsal striatum. I also helped write my part of the manuscript. The article has been accepted for publication in Nature Communications. I have included a copy of the manuscript in Appendix 4: Transcriptional repression of Plxnc1 by Lmx1a and Lmx1b direct topographic dopaminergic circuit formation. I was also asked to collaborate on a project led by the group of Alyson Fournier at McGill University. I was asked to confirm, by performing electrophysiological recordings, that MT3-MMP promotes excitatory synapse formation. The abstract, a significance statement as well as my contribution to the paper is in Appendix 5: MT3-MMP promotes excitatory synapse formation by promoting Nogo-66 receptor ectodomain shedding.
During my PhD, I optimized a method to culture and bulk electroporate organotypic slices. This technique was used in two projects. The first one is in collaboration with researchers from many universities, as well as Katalin Toth from Laval University. It aims to characterize a genetically encoded protein and its efficiency for reporting changes in membrane potential. It has recently been accepted for publication in eLife. Seeing as I have not contributed to the figures, I have not added a copy of the manuscript, but rather an abstract of the findings in Appendix 6: Fast two-photon imaging of subcellular voltage dynamics in neuronal tissue with genetically encoded indicators. A second project was made possible using organotypic slice cultures and bulk electroporation. This project is still in the works and aims to develop the use of a voltage-sensitive dye for imaging changes in potential in small compartments, mainly spines. For this project, I planned and designed the experiments, performed and analyzed electrophysiological recordings and wrote the manuscript with Olivier Dupont-Thérrien. I have added a copy of the manuscript in Appendix 7: Optical approach for the measurement of electrical activity in neurons with voltage-sensitive dye ANNINE-6plus.
1
1. Introduction
The brain is the most complex organ in the body. It controls, among many things, the other organs of the body, language, movement, sleep and wakefulness, memory, emotions and thoughts. It is an astonishing structure, whether looked at in molecular, cellular, systemic, behavioural, or cognitive terms (Purves, 2004).
Neuronal circuits, ensembles of neurons and glial cells of a specific anatomical location, process specific types of information. These systems have three main functions, sensory, motor and association. The sensory system represents information about the organism and its environment. The motor system organizes and generates actions. The associative systems link the sensory and motor systems, providing the basis for complex functions such as perception, attention, cognition, emotions, rational thinking, and many other intricate brain functions.
Much remains to be learned on the detailed functions that the many neuronal circuits of the brain enable. This knowledge is essential to improve our capacity to treat or cure brain-related neurological and psychiatric disorders. My thesis is focused on the dopaminergic system, on its genetically regulated development and circuit connectivity, and on its functions associated with hyperactivity.
1.1. The dopamine system
The dopaminerergic system is comprised of neurons which secrete a specific neurotransmitter: dopamine. Most cells of the dopaminergic system are part of the midbrain, or mesencephalon. The midbrain’s main roles include motor control, vision, hearing, sleep/wake, reward, learning and motivation. The two main structures of the midbrain containing dopaminergic neurons are the substancia nigra pars compacta (SNpc) and the ventral tegmental area (VTA) (Figure 1-1: Midbrain section in adult mice.).
2 Figure 1-1: Midbrain section in adult mice.
Schematic representation of the midbrain in adult mice showing the location of the VTA (red) and the SNpc (green) in a coronal section of a mouse.
1.1.1. The substancia nigra pars compacta
The substancia nigra, Latin for “black substance”, has been named so because of its dark structure, explained by the expression of neuromelanin in dopaminergic neurons (Rabey
and Hefti, 1990). The main function of the substancia nigra pars compacta (SNpc) is motor
control. This is an indirect role as electrical stimulation of the SNpc does not result in movement (Hodge and Butcher, 1980). Degeneration of SNpc dopaminergic neurons is what underlies Parkinson’s disease. The SNpc is also involved in learned responses to stimuli. Dopaminergic neurons of the SNpc principally innervate the dorsal part of the striatum (Bjorklund and Dunnett, 2007; Roeper, 2013) and form the nigrostriatal pathway.
1.1.2. The ventral tegmental area
Dopaminergic neurons of the ventral tegmental area (VTA) have been shown to play roles in positive and negative reinforcement, decision-making, working memory, and aversion
(Bromberg-Martin et al., 2010). The VTA is also the major focus of neuropsychiatric
disorders.
About 50 to 80% of neurons in the VTA are reported to be dopaminergic (Johnson and North,
1992; Margolis et al., 2006b). Dopaminergic neurons of the VTA principally innervate the
dorsal part of the striatum and the prefrontal cortex (Bjorklund and Dunnett, 2007; Roeper,
3
1.2. Dopamine
Dopamine is one of the earliest neurotransmitters expressed in the developing brain. The presence of dopaminergic projections in the developing striatum and cortex, as well as precursor cells expressing dopamine receptors, places dopamine as an important modulator of neuronal development. This includes cell proliferation, differentiation and migration, all of which can lead to dysfunctional synapses and altered connectivity, which in turn could contribute to striatal and cortical dysfunction in neuropsychiatric disorders (Money and
Stanwood, 2013). Dopamine acts on the neuronal circuit by slowly modulating rapid synaptic
transmission mediated by γ-aminobutiric acid (GABA) and glutamate (Beaulieu and
Gainetdinov, 2011).
Tyrosine hydroxylase (TH) is the enzyme responsible for converting the amino acid L-tyrosine in L-3,4-dihydroxyphenylalanine (L-DOPA), which is then converted to dopamine by the DOPA decarboxylase (Figure 1-2: Biosynthesis pathway for dopamine.) (Kaufman,
1995; Nagatsu, 1995).
Figure 1-2: Biosynthesis pathway for dopamine.
Dopamine is synthesized from tyrosine through two enzymatic reactions. The Tyrosine hydroxylase by hydroxylation of the aromatic cycle of the tyrosine leads to L-DOPA synthesis. Then the L-Dopa is transformed into dopamine by decarboxylation via DOPA decarboxylase. [From (Hare and Loer,
2004)]
1.2.1. Roles and functions
Once in the synapse, dopamine binds to dopamine receptors (Beaulieu and Gainetdinov,
2011). These can be postsynaptic dopamine receptors (D1 and D2), which are located on
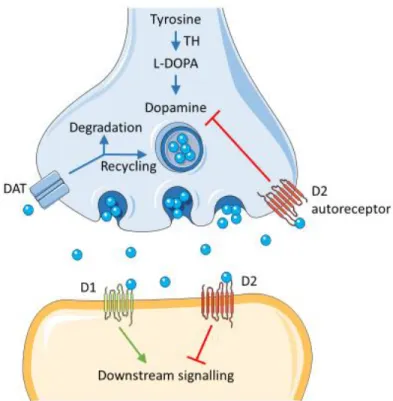
dendrites, or presynaptic D2-autoreceptors (Beaulieu and Gainetdinov, 2011). After the postsynaptic neuron evokes an action potential, dopamine quickly becomes unbound from its receptors. It is then absorbed back into the presynaptic cell, via reuptake mediated mostly by the dopaminergic transporter (DAT) (Figure 1-3: Schematic representation of dopaminergic synapse). Once back in the cytosol, dopamine can either be broken down by
4
a monoamine oxidase or repackaged into vesicles, making it available for eventual re-release (Eiden et al., 2004).
Figure 1-3: Schematic representation of dopaminergic synapse
Dopamine in the cleft can bind presynaptically to D2 autoreceptors or the DAT, or postsynaptically to D1- and D2-class receptors. The DAT is predominately located perisynaptically.
1.2.2. The dopamine transporter
The dopamine transporter (DAT) is a protein at the cell membrane that is responsible for pumping dopamine out of the synaptic cleft back into the cytosol. In the cytosol, other transporters are responsible for sequestering dopamine into vesicles for storage for eventual re-release. DAT function requires the binding and co-transport of two Na+ ions and one Cl−
ion with dopamine. The driving force for DAT-mediated dopamine reuptake is the ion concentration gradient generated by the membrane-bond Na+/K+ ATPase (Torres et al.,
2003).
1.2.3. Receptors
Following its release by presynaptic terminals, dopamine activates G-protein coupled receptors (Beaulieu and Gainetdinov, 2011). These receptors are divided in 2 main groups:
5
D1-like (D1 and D5) and D2-like (D2, D3, and D4) (Andersen et al., 1990; Sibley and
Monsma, 1992; Missale et al., 1998; Beaulieu and Gainetdinov, 2011) (Figure 1-4:
Schematic of dopaminergic receptors). These receptors are already present in the striatum, even before dopaminergic axons reach the structure (Schambra et al., 1994; Araki et al.,
2007; Sillivan and Konradi, 2011). They have different effects on intracellular cascades in
post synaptic cells.
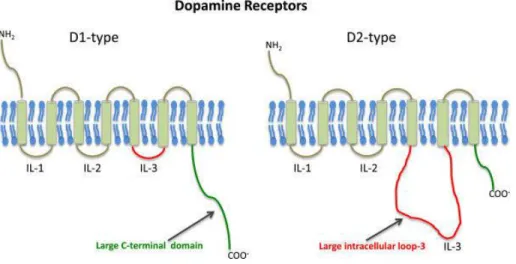
Figure 1-4: Schematic of dopaminergic receptors
Schematic of structures highlighting comparative general differences between D1- and D2-like dopamine receptors [From (Pandey et al., 2013)].
Binding of dopamine to its receptors can lead to many cellular processes, which can impact cellular morphology. D1-like receptor stimulation leads to increased neurite localization of cytoskeletal elements needed for cell motility, whereas D2-like receptor stimulation increased cell body localization of these cytoskeletal elements (Crandall et al., 2007). D1-like receptor stimulation increases neurite outgrowth in the striatum in vitro (Schmidt et al.,
1996; Schmidt et al., 1998) and reduces neurite outgrowth in frontal cortical neuronal
cultures (Reinoso et al., 1996; Song et al., 2002; Li et al., 2013). Unpublished data from Money and Stanwood suggests that D1 receptor null mice had increased basal neurite outgrowth in cortical cultures (Money and Stanwood, 2013). Cortical primary cultures where D2-like receptor was activated showed increased neurite growth (Todd, 1992; Reinoso et
al., 1996). D3 receptors, can guide adult neurogenesis as well (Kim et al., 2010; Egeland et al., 2012; Lao et al., 2013). The roles for dopamine receptors in cellular morphology seem
to be cell type specific and requires more studies to elucidate the mechanisms of each receptor in each cell type.
6
1.2.3.1. D1-like receptors
Receptors of class D1 (D1 and D5) activate the type Gαs/olf family of G proteins to stimulate
the production of AMPc by adenylyl cyclase. They are found mainly on postsynaptic sites of dopamine-sensitive cells, such as medium spiny neurons of the striatum.
The activation of D1 receptors inhibits the evoked activity at hyperpolarizing membrane potentials (Calabresi et al., 1987) and facilitates action potential activation when the cell is at a depolarizing potential (Hernandez-Lopez et al., 1997). However, in vivo experiments show that tonic activation of D1 receptors increases membrane excitability (West and Grace,
2002).
Alterations in D1 receptor signalling during the maturation of the cortex in adolescence could play a significant role in the development of neuropsychiatric diseases (Money and
Stanwood, 2013). For example, a decrease in dopaminergic axons in layers of medial
prefrontal cortex and decreased D1 receptor binding occurs in schizophrenic patients
(Okubo et al., 1997; Akil et al., 1999). Additionally, it has been shown that D1 receptor
activation enhances cortically drug seeking behaviours (Piazza et al., 1991; Kalivas et al.,
2005). Clearly, D1 receptors could be targeted in treatment of such disorders.
1.2.3.2. D2-like receptors
Receptors of class D2 (D2, D3 and D4) activate the type Gαi/o family of G-protein which
inhibits adenylyl cyclase (Beaulieu and Gainetdinov, 2011). D2 and D3 receptors are expressed both on the postsynaptic cell and on the presynaptic side of the dopaminergic cell (Sokoloff et al., 2006; Rondou et al., 2010).
Activation of D2 receptors reduces the action potential firing when the cell is depolarized
(Hernandez-Lopez et al., 2000). However, in vivo experiments show that tonic activation of
D2 receptors decreases membrane excitability in striatal neurons (West and Grace, 2002).
Increased D2 receptor binding of dopamine in the striatum has been seen in patients with attention deficit/hyperactivity disorder (ADHD) and depression (D'Haenen and Bossuyt,
1994; Shah et al., 1997), which suggests that targeting D2 receptors could provide treatment
7
1.2.3.3. Expression of dopamine receptors
Dopamine receptors are expressed at different levels in the brain, with some structures expressing more D1- or D2-like receptors. In the nucleus accumbens and the dorsal striatum, patterns of expression of D1 and D2 receptors are similar. The shell of the nucleus accumbens has high expression of D3 receptors (Le Moine and Bloch, 1995). Dopamine receptor expression is lower in the frontal cortex, where D1 receptors seem to be the predominant form. In this structure, both D1 and D2 receptor are found in glutamatergic projecting neurons of layer V/VI and parvalbumin expressing GABAergic interneurons (Le
Moine and Gaspar, 1998; Muly et al., 1998). D4 and D5 receptors are expressed at low
levels in the frontal cortex (Beaulieu and Gainetdinov, 2011).
1.3. Main dopaminergic pathways
Dopaminergic neurons are known to project to far away target regions. These pathways are critically involved in various key functions of the mammalian brain, including voluntary movement, affection, reward, attention, and learning (Grace et al., 2007; Redgrave et al.,
2010). Dopaminergic circuitry dysfunctions are linked to the development of
neurodegenerative and neuropsychiatric disorders, including ADHD, Parkinson’s disease, schizophrenia and disorders of the obsessive-compulsive spectrum. There are 3 main dopaminergic pathways: the nigrostriatal the mesocortical and mesolimbic pathways (Figure 1-5: Dopaminergic pathways).
Figure 1-5: Dopaminergic pathways
Schematic representation of the midbrain in adult mice showing the location of the VTA (red) and the SNpc (green). [From: (Money and Stanwood, 2013)]
8
1.3.1. Nigrostriatal pathway
The nigrostriatal pathway encompasses dopaminergic neurons from the SNpc innervating the dorsal striatum (caudate nucleus and putamen) (Money and Stanwood, 2013). This pathway is mostly involved in motor behaviour (Bjorklund and Dunnett, 2007; Van den
Heuvel and Pasterkamp, 2008). Loss of SNpc dopaminergic neurons is the main
pathological findings in Parkinson’s disease, which lead to a concomitant loss of dopaminergic input in the striatum (Davie, 2008).
1.3.2. Mesolimbic pathway
The mesolimbic pathway is comprised of dopaminergic neurons from the VTA projecting to the ventral part of the striatum, and the nucleus accumbens (NA). This pathway plays an important role in the reward system (Money and Stanwood, 2013), motivated behaviour and reinforcement learning (Ikemoto and Panksepp, 1999; Schultz et al., 2000). Dysregulations in this pathway can be one of the causes of addiction (Dreyer, 2010). This pathway is also implicated in schizophrenia and depression (Laviolette, 2007; Van den Heuvel and
Pasterkamp, 2008), as well as ADHD (Viggiano et al., 2002; Engert and Pruessner, 2008; Nestler et al., 2009; Volkow et al., 2009).
1.3.3. Mesocortical pathway
The mesocortical pathway is comprised of dopaminergic neurons from the VTA projecting to the prefrontal cortex (PFC) regulating cognitive, learning, memory, and emotional functions (Gorelova et al., 2012). This pathway is also implicated in schizophrenia
(Abi-Dargham and Moore, 2003), as well as ADHD (Viggiano et al., 2003; Engert and Pruessner, 2008; Nestler et al., 2009; Volkow et al., 2009).
1.3.4. Inputs on dopaminergic neurons
The SNpc and the VTA dopaminergic neurons receive different types of inputs from many different regions of the brain.
1.3.4.1. Inputs to the VTA
Recent methodological advances, such as rabies-based methods, have helped in the mapping of brain structures that project to dopaminergic neurons of the VTA neurons
9
These studies suggest that VTA dopamine receive inputs from various brain regions and that there are some qualitative, but not quantitative, differences in their input patterns
(Watabe-Uchida et al., 2012; Ogawa et al., 2014; Beier et al., 2015; Faget et al., 2016). The
main excitatory input on dopaminergic neurons in the VTA is from the medial prefrontal cortex (mPFC) (Carr and Sesack, 2000). The lateral habenula (LHb) also projects to both glutamatergic and GABAergic neurons to the VTA. This pathway has a potential role in motivation, shown using a conditioned place aversion test (Lammel et al., 2012). The excitatory inputs to the VTA also comes from the laterodorsal tegmentum (LDTg). This pathway has a role in motivation, as demonstrated by stimulation or inhibition of LDTg fibers in a conditioned place preference test (Omelchenko and Sesack, 2005; Lammel et al., 2012;
Steidl et al., 2017). A major glutamatergic input to the VTA arises from the dorsal raphe
nucleus (DRN). This pathway activates AMPA receptors from vesicular glutamate transporter type 3 (VGLUT3) inputs which enhances DA levels in the nAcc. This input plays a role in self-stimulation (Qi et al., 2014). The lateral hypothalamus (Watabe-Uchida et al.,
2012), pedunculopontine tegmentum (PPTg) (Charara et al., 1996), amygdala (Morales and Margolis, 2017), periaqueductal grey (PAG) (Omelchenko and Sesack, 2010) bed nucleus
of the stria terminalis (BNST) (Georges and Aston-Jones, 2001) and the VTA itself (Dobi et
al., 2010; Wang et al., 2015) all send glutamatergic projections to the VTA (Figure 1-6: Inputs
to ventral tegmental area neurons).
Figure 1-6: Inputs to ventral tegmental area neurons
The schematics summarize the synaptic connections that have been detected either anatomically (using electron microscopy) or functionally (using optogenetics in conjunction with ex vivo electrophysiology). [From (Morales and Margolis, 2017)]
10
Anatomical findings show GABAergic projections from nucleus accumbens (NAc) are the main inhibitory inputs to the VTA (Watabe-Uchida et al., 2012; Beier et al., 2015). The rostromedial mesopontine tegmental nucleus (RMTg) (Jhou et al., 2009; Kaufling et al.,
2010), the PAG (Omelchenko and Sesack, 2010), the DRN (Beier et al., 2015), the lateral
hypothalamus (LHT) (Nieh et al., 2015), the ventral palladium (VP) (Hjelmstad et al., 2013) and the VTA itself (Omelchenko and Sesack, 2009; Dobi et al., 2010; Tan et al., 2012; van
Zessen et al., 2012) have all been shown to project to the VTA.
The VTA also receives modulatory inputs. It has been demonstrated that neurons located in the LDTg make cholinergic input on dopaminergic neurons (Russo and Nestler, 2013). The VTA also receives orexinergic input from the lateral hypothalamus (Russo and Nestler,
2013). Serotoninergic input to the VTA comes mainly from the DRN (Qi et al., 2014).
1.3.4.2. Inputs to the SNpc
Little is known about the neuronal inputs to the SNpc. The PPTg neurons send glutamatergic projections to the SNpc (Jackson and Crossman, 1983; Futami et al., 1995; Oakman et al.,
1995; Parent et al., 1999; Zahm et al., 2011; Watabe-Uchida et al., 2012). Glutamatergic
input can also come from the somatosensory and motor cortex, which seems to be the main input to the SNpc (Watabe-Uchida et al., 2012). The subthalamic nucleus also provides excitatory inputs to the SNpc (Kita and Kitai, 1987; Groenewegen and Berendse, 1990;
Smith et al., 1990; Watabe-Uchida et al., 2012).
Inhibitory input can come from the central nucleus of the amygdala, globuls pallidus, entopeduncular nucleus and the dorsal raphe nucleus (Watabe-Uchida et al., 2012). The striatum also sends GABAergic input to the SNpc, specifically from compartments called striosomes (Squire, 2013).
Inputs to the SNpc can also come from the central nucleus of the amygdala, the superior colliculus, the parabrachial nuclei and the basal forbrain (Dommett et al., 2005; Matsumoto
11
1.4. Dopaminergic neurons
1.4.1. Origin and development
The first key steps in the formation of dopaminergic circuits are the primary events that will lead to the establishment of the ventral midbrain. During gastrulation, postero-anterior cell migration occurs while mesoderm, endoderm and ectoderm are formed (Arenas et al.,
2015). The secretion of the rostral side of the embryo suppresses posterior signals and
allows the implementation of the neural ectoderm, thus leading to the formation of the anterior neural tube (Takaoka et al., 2007). The dorsal ectoderm will be restricted to a neuronal destiny in response to signals, giving rise to the neural plate (Hemmati-Brivanlou
and Melton, 1997; Harland, 2000; Liu and Niswander, 2005). This will be subsequently
subdivided into restricted domains, guided by a gradient of different morphogens expressed along the anteroposterior and dorsoventral axis, while being closed on itself to form the neural tube (Simon et al., 1995; Puelles, 2001; Ulloa and Briscoe, 2007) (Figure 1-7: Neurulation in the mammalian embryo).
Figure 1-7: Neurulation in the mammalian embryo
On the left are dorsal views of the embryo at several different stages of early development; each boxed view on the right is a midline cross section through the embryo at the same stage. (A) During late gastrulation and early neurulation, the notochord forms by invagination of the mesoderm in the region of the primitive streak. The ectoderm overlying the notochord becomes defined as the neural plate. (B) The neural plate begins to fold at the midline, forming the neural groove and ultimately the neural tube. The neural plate immediately above the notochord differentiates into the floor plate, whereas the neural crest emerges at the lateral margins of the neural plate. (C) Once the edges of the neural plate meet in the midline, the neural tube is complete. The mesoderm adjacent to the tube
12
then thickens and subdivides into structures called somites—the precursors of the axial musculature and skeleton. (D) The neural tube, adjacent to the somites, becomes the rudimentary spinal cord, and the neural crest gives rise to sensory and autonomic ganglia. Finally, the anterior ends of the neural plate grow together at the midline and continue to expand, eventually giving rise to the brain. [From (Purves, 2004).]
As the formation of the neural tube continues, two important signal centres are formed: the isthmus organizer, defining the boundary between the midbrain and the rhombencephalon, and the floor plate (Wassarman et al., 1997; Rhinn et al., 1998; Joyner et al., 2000). The establishment of the floor plate is dependent on the secretion of the sonic hedgehog (Shh) signaling protein. Initially secreted by the notochord, Shh specifies the most ventral portion of the neural plate, the floor plate, by inducing expression of Foxa2 to E8 in mice (Ang et al.,
1993; Sasaki et al., 1997). At E8.5, the floor plate starts expressing Shh and becomes the
second ventral organ of the neural tube (Echelard et al., 1993; Hynes et al., 1995; Ho and
Scott, 2002). A Shh gradient from the floor plate then regulates ventro-dorsal placement:
ventral progenitors exposed to a higher concentration of Shh respond by expressing different transcription factors, as opposed to dorsal cells exposed to lower concentrations
(Marti et al., 1995; Roelink et al., 1995; Chiang et al., 1996; Ericson et al., 1996; Briscoe and Ericson, 1999). The floor plate keeps its ventral midbrain identity by stopping Shh signalling (Ribes et al., 2010; Metzakopian et al., 2012). During a specific time window at the induction
of the midbrain floor plate, the simultaneous expression of FGF8 from the isthmus organizer in the mid/hindbrain boundary and Shh signalling from the floor plate leads to the induction of mDA progenitors (Hynes et al., 1995).
Neurogenesis of dopaminergic midbrain neurons occurs at the ventricular zone of the floor plate. The differentiation process of dopaminergic neurons is regulated by several early transcription factors, including Otx2, Lmx1a/b, FoxA1/2, as well as En1/2 homeodomain transcription factors. These transcription factors remain expressed by postmitotic dopaminergic cells, and each is incapable of inducing a complete dopaminergic phenotype, suggesting that they work cooperatively (Hegarty et al., 2013). Dopaminergic precursors divide to generate immature dopaminergic neuron expressing the nuclear receptor related 1 protein (Nurr1) and Pitx3 (Arenas et al., 2015). In rodents, the first dopaminergic neurons appear at E10.5, whereas in humans, neurogenesis of dopaminergic neurons begins between the fifth and sixth week of embryonic development and ends before the end of the eleventh week (Freeman et al., 1991; Almqvist et al., 1996; Nelander et al., 2009).
13
1.4.2. Transcription factors of mDA
During the different stages of development, the combined action of transcription factors and morphogens from these signalling centres orchestrate multiple functions, including the definition of the regional identity of the ventral midbrain, specification and proliferation of dopaminergic progenitors, neurogenesis of mDA neurons, as well as their differentiation and survival (Arenas et al., 2015).
Dopaminergic progenitors exit the cell cycle and generate postmitotic immature dopaminergic neurons between E9.5 and E12.5 mice (Bayer et al., 1995). Immature neurons initiate expression of Nurr1 (Zetterstrom et al., 1997) and En1/2(Simon et al., 2001; Alberi
et al., 2004). From E11 onwards, immature dopaminergic neurons continue to migrate
radially on radial glia, further differentiating in dopaminergic neurons (Kawano et al., 1995). Mature dopaminergic neurons express Pitx3 and TH, in addition to the various markers expressed in immature dopaminergic neurons (Ang, 2006) (Figure 1-8: Transcriptional control of ventral midbrain dopaminergic neurons).
Figure 1-8: Transcriptional control of ventral midbrain dopaminergic neurons
(A) The sequential timing of transcription factor activation in mDA progenitors. The curved arrow indicates cycling cells. (B) The timing of expression of transcription factors in mDA progenitors (not in subsequent steps of the lineage) are shown. Dotted lines indicate that the precise timing of expression shutdown has not been determined [Adapted from: (Saucedo-Cardenas et al., 1998;
14
1.4.2.1. Otx2
Otx2 is a bicoid class homeodomain transcription factor that is widely expressed before gastrulation, but its expression becomes progressively exclusive to the anterior third of the mouse embryo after E7.5 (Simeone et al., 1993; Ang et al., 1994). Expression of Otx2 is restricted to the forebrain and the midbrain between E8.5 and E12.5. From E12.5 and onward, expression is also detected in the rhombencephalon (Mallamaci et al., 1996). Otx2 is required for the formation of the forebrain and the midbrain as a result of its role in the anterior visceral endoderm, where it functions to restrict the fate of the posterior embryo
(Mallamaci et al., 1996; Perea-Gomez et al., 2001). Loss of Otx2 expression from E10.5
results in loss of expression of Ngn2 and Mash1 in ventral dopaminergic progenitors (Vernay
et al., 2005). Otx2, by regulating Mash1 and Ngn2 expression, is required for the generation
of dopaminergic progenitors (Ang, 2006). Otx2 expression is limited to VTA neurons in the adult brain (Di Salvio et al., 2010). Otx2 regulates proliferation in part through regulation of Lmx1a and through regulating Wnt1 independently of Lmx1a and Lmx1b (Omodei et al.,
2008).
1.4.2.2. En1/2
In mammals, homeoprotein Engrailed exists in 2 forms: Engrailed-1 (En1) and (En2). They play critical roles in patterning and neurogenesis during central nervous system development (Simon et al., 2004). En1/2 help primarily to regulate development in the dorsal midbrain and anterior hindbrain (Sgaier et al., 2007). They are expressed by SNpc and VTA neurons from early development and throughout adulthood (Simon et al., 2001).
1.4.2.3. FOXA1/2
FOXA1 and FOXA2 transcription factors play a critical role in the generation of mDA neurons during early and late development, regulating both specification and differentiation in a dose-dependent manner (Ang, 2009). FOXA1/2 are necessary to maintain normal synthesis and transmission of dopamine as well as the burst activity of SNc neurons in adult mice (Pristera
et al., 2015).
1.4.2.4. Lmx1a/b
Lmx1a and Lmx1b belong to the Lmx group of LIM transcription factors (needed for the development of the pituitary gland and the specification of motoneurons), and share 64% of