RACHEL THEORET-GOSSELIN
EFFETS DES SOINS MATERNELS, DU
DÉVELOPPEMENT MOTEUR ET DES CONDITIONS
ENVIRONNEMENTALES SUR LA SURVIE DES
JEUNES CHÈVRES DE MONTAGNE
Mémoire présenté
à la Faculté des études supérieures et postdoctorales de l'Université Laval dans le cadre du programme de maîtrise en Biologie
pour l'obtention du grade de maître es sciences (M.Sc.)
DEPARTEMENT DE BIOLOGIE FACULTÉ DES SCIENCES ET DE GÉNIE
UNIVERSITÉ LAVAL QUÉBEC
2012
RESUME
Selon la théorie des composantes biodémographiques, les soins maternels devraient augmenter les chances de survie des jeunes. La plupart des études ont par contre étudié les effets des conditions environnementales et des traits maternels sur la survie des jeunes. Nous avons évalué l'effet des soins maternels et du développement du jeune sur la survie des chevreaux d'une population marquée de chèvres de montagne, Oreamnos americanus. Nous avons confirmé l'influence des conditions environnementales et des traits maternels sur la survie des jeunes. Or, les soins maternels et le développement du jeune étaient les déterminants les plus influents de leur survie. Les soins prodigués par la mère augmentaient les probabilités de survie des jeunes en hiver tandis que les jeunes jouant fréquemment augmentaient leurs chances de survie jusqu'au sevrage. Nous concluons que la qualité des soins parentaux et les comportements de jeu devraient être considérés dans les études de dynamique de population.
ABSTRACT
Life history theory predicts that maternal care should increase offspring survival. However, population dynamic studies have mainly focused on the effects of environmental conditions and maternal traits on offspring survival. Here, we evaluated the influence of maternal care and offspring development on kid survival in a long-term study of marked mountain goats, Oreamnos americanus. We confirmed the influence of environmental conditions and maternal traits on offspring survival. However, maternal care and play behaviour were the strongest determinants of kid survival. Maternal care enhanced offspring survival after weaning while kids exhibiting frequent play behaviours increased their survival chances before weaning. We conclude that maternal care and play are crucial factors influencing juvenile survival and ultimately population dynamics.
Ill
AVANT-PROPOS
Ma maîtrise est présentée avec insertion d'un article scientifique en anglais mis en contexte et interprété par une introduction et une conclusion en français. Je suis l'auteure principale de ce mémoire et de cet article. La contribution de chaque auteur de l'article va comme suit :
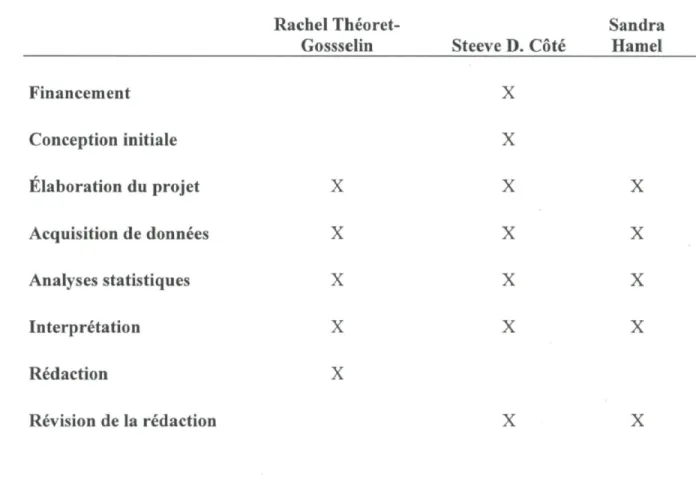
TABLEAU i. Contribution relative des auteurs de l'article scientifique.
Rachel Théoret- Sandra Gossselin Steeve D. Côté Hamel
X X X X X X X X X X X X X X Financement Conception initiale Elaboration du projet Acquisition de données Analyses statistiques Interprétation Rédaction Révision de la rédaction X X
Au cours des dernières années, j'ai eu la chance de côtoyer plusieurs personnes qui ont participé de près ou de loin à ce mémoire et je tiens à prendre le temps de les remercier. D'abord, le premier merci va à mon directeur de maîtrise, Steeve Côté, pour m'avoir fait assez confiance à l'hiver 2008 et me donner une chance de faire mes preuves sur la montagne à l'été suivant. Sa passion des chèvres, le soutien logistique, moral et financier que Steeve m'a fourni ont été sans contredit le moteur de ce projet. Merci d'avoir été à mes
IV
côtés! Une autre personne a été plus qu'essentielle à la réalisation de mon projet tel qu'il est, Sandra Hamel. Sa passion pure du terrain ne pouvait que me faire mourir d'envie de faire une maîtrise sur les biquettes. Les premières tempêtes, les premières identifications et les premières observations cibles se sont faites à ses côtés, sous son oeil vigilant et patient. Sandra a été comme une mère pour moi; exigeante et motivante, mais seulement parce qu'elle savait jusqu'où elle pouvait me mener. Elle m'a appris beaucoup, tant en science, en statistique que sur la vie. Je ne te remercierai jamais assez pour tout le soutien et l'aide précieuse que tu m'as fournis aux moments où j'en avais besoin. Il y a aussi plusieurs autres chercheurs qui ont pris le temps de lire ou de discuter avec moi à propos de mon projet et qui ont amené des idées et des pistes de réflexion intéressantes. Parmi ceux-ci, je compte Gwenael Beauplet, Julien Béguin, Marco Festa-Bianchet, Gilles Gauthier, Julien Martin, Kathreen Ruckstuhl, Bill Shipley et Jean-Pierre Tremblay.
Un projet comme le mien nécessitait plusieurs mois sur le terrain, loin de territoires connus et confortables de la maison. La présence de Kirby Smith et de Dave Hobson, non loin de Grande Cache, toujours prêts à intervenir lorsqu'ils pouvaient nous faciliter la vie a été essentielle. Je tiens également à remercier la famille Ewald et particulièrement Mike. Ça a toujours été un plaisir de voir des visages familiers et de se sentir les bienvenus à n'importe quel moment où nous voulions socialiser un peu. En plus du soutien moral, Mike a toujours été là pour l'organisation et la « logistique » du projet à Grande Cache et même aux alentours. Now I try to use the expression: "Is that right?" as much as I can, thanks a lot! Merci particulier également à tout le personnel de Fish and Wildlife Service de Grande Cache, Penny Cannon, Karen Guillemette et Shane Ramstead, pour l'accès à la photocopieuse, les carnets de notes Report a Poacher, les mises à jour de la météo et les beaux sourires. Je dois remercier aussi le personnel de SRD de Grande Prairie, spécialement Dave Stepnisky pour la réparation rapide du pont après le déluge de l'été 2009 et pour l'aide sur une multitude d'autres points. Un merci également à Karen Stroebel pour son aide et la facilité de communication qu'elle nous a permis avec la mine pour assurer notre arrivée à bon port lors de certains déplacements dangereux. Je tiens à remercier spécialement toutes les personnes qui sont passées sur la montagne et qui ont permis un suivi des biquettes tellement précieux, non seulement pour mon étude, mais pour
le projet en entier. Un merci spécial à ceux qui ont passé plusieurs heures les fesse gelées et le cou cassé pour observer ces chevreaux : Chantai Beaudoin, Yannick Gendreau, Steeve Côté, Geneviève Simard, Sandra Hamel, Mélina Houle, Etienne Cardinal, Cassandra Cameron, Alexandre Veillette et Mark O'Connor. Particulièrement, j'ai eu la chance de travailler avec des gens qui ont fait de mes séjours à Caw Ridge des moments inoubliables : Alexandre Veillette, Mark O'Connor, Karine Dancose, Alexandre Auger, Aaron Shafer, Olivier Simard et Etienne Cardinal. Avant le départ et de retour du terrain, l'aide apportée par Sonia de Bellefeuille et Caroline Hins pour l'organisation matérielle, bureaucratique et le soutien moral a été plus qu'appréciée, merci beaucoup!
Évidemment, je n'ai pas passé ces trois années sur la montagne; j'ai également rencontré des gens très importants qui m'ont entourée de retour à la civilisation. Pour des discussions stimulantes, de l'aide informatique, des conseils, ou simplement une bonne jasette, merci à tous ceux du labo qui ont été à mes côtés plus ou moins longtemps : Martin Barrette, Etienne Cardinal, William Crosmary, François Lebel, Léon L'Italien, Ariane Massé, Anouk Simard, Joëlle Taillon, Antoine St-Louis, Karine Pigeon, Vanessa Viera, Mael Le Corre, Marianne Bachand, Alexandre Veillette, Julie Yolande Faure-Lacroix, Glenn Yannic, Gaëlle Darmon, Albert Hidding, Alice-Anne Simard, Amélie Drolet, Emilie Champagne, Janick Gingras, Julie Lambert, Julien Hénault Richard, Sébastien Lavoie, Valérie Saucier, Alexandre Rasiulis, Karine Dancose, Sabrina Courant et Geneviève Doufour-Tremblay. Merci à Joëlle, Gaëlle, Glenn et Karine pour vos corrections et commentaires sur divers textes et présentations qui ont découlés de cette maîtrise. Merci également à Katherine Charland pour l'aide dans la saisie de données. Un remerciement particulier à Yolande et Mael, pour votre soutien en fin de soirée de 5 à 7 et pour votre amitié. Finalement, mille et un merci à Karine qui ne m'a qu'encouragée, conseillée et motivée tout au long de ma maîtrise. Je suis vraiment heureuse d'avoir eu la chance de compter des bleuets, résoudre des problèmes complexes de physique diesel et faire des shootings photo avec toi dès le début. Ton amitié et ta présence chaleureuse m'ont permis de relever ce défi que je m'étais lancé en voyant combien on pouvait avoir du plaisir en été 2008. Merci vraiment beaucoup d'avoir été le meilleur boss au monde!
VI
Je remercie également ma famille et mes amis qui ont su faire preuve de compréhension et de patience lors des moments plus difficiles. Vos encouragements et votre présence tout au long de mon projet ont été essentiels, je n'aurais jamais réussi sans vous. Maman, Papa, Rémi, Marie, Sophie, Annie-Claude, Christine et Mark, merci! Finalement, merci à Olivier. C'est avec un sourire et de beaux souvenirs que je raconte que c'est la montagne qui nous a mis sur le chemin l'un de l'autre. Merci de partager ma vie!
Finalement, je remercie tous nos partenaires financiers qui ont rendu possible cette étude : l'Alberta Conservation Association (ACA), le Conseil de Recherche en Sciences Naturelles et en Génie du Canada, l'Alberta Fish and Wildlife Division, le Fonds Québécois de la Recherche sur la Nature et les technologies, l'Université Laval, le Centre d'Études Nordiques, l'Université de Sherbrooke, La fondation Rocky Mountain Goat, la fondation de l'Alberta Sports, Recreation, Parks, and Wildlife, Les Offices Jeunesse Internationaux du Québec, l'Association des Chercheurs et Chercheuses Étudiants en Biologie de l'Université Laval, l'Association des Étudiantes et Étudiants de Laval inscrits aux Études Supérieures.
Vil
TABLE DES MATIERES
RÉSUMÉ i ABSTRACT ii AVANT-PROPOS iii
TABLE DES MATIÈRES vii LISTE DES TABLEAUX ix LISTE DES FIGURES x LISTE DES ANNEXES xi CHAPITRE 1 - Introduction générale 1
CADRE THÉORIQUE 3 La masse corporelle 4 Les soins maternels 5
Le sexe 7 Les facteurs environnementaux 7
OBJECTIFS GÉNÉRAUX ET SPÉCIFIQUES 9
LA CHÈVRE DE MONTAGNE. 9
L'AIRE D'ÉTUDE 11 Capture et marquage 11 LES OBSERVATIONS COMPORTEMENTALES 12
CHAPITRE 2 - Why care? The role of maternal behaviour and offspring
development in the survival of mountain goat kids 15
RÉSUMÉ 17 ABSTRACT. 18
INTRODUCTION 19 METHODS 21
Study area and captures 21
Survival 22 Environmental conditions 22
Masses 23 Behavioural observations 23
Statistical analyses 27 First step: Variables selection 27
Second step: Confirmatory multilevel path analysis '. 29
RESULTS 31 Variables selection 31
Path analysis for summer survival 31 Path analysis for winter survival 34
DISCUSSION 35 Kid development 36
vin Maternal care 37 Maternal traits 38 Environmental conditions 39 ACKNOWLEDGEMENTS 40 CHAPITRE 3 - Conclusions 41
Sommaire des résultats 43 Les comportements de jeu 44 Les soins maternels 45 Contributions 46 BIBLIOGRAPHIE 49
ANNEXE A 63 ANNEXE B 67
IX
LISTE DES TABLEAUX
TABLEAU i
TABLE 1
TABLE 2
Contribution relative des auteurs de l'article
scientifique iii Descriptions of the behavioural variables representing maternal care
and kid developmental processes for mountain goats at Caw Ridge,
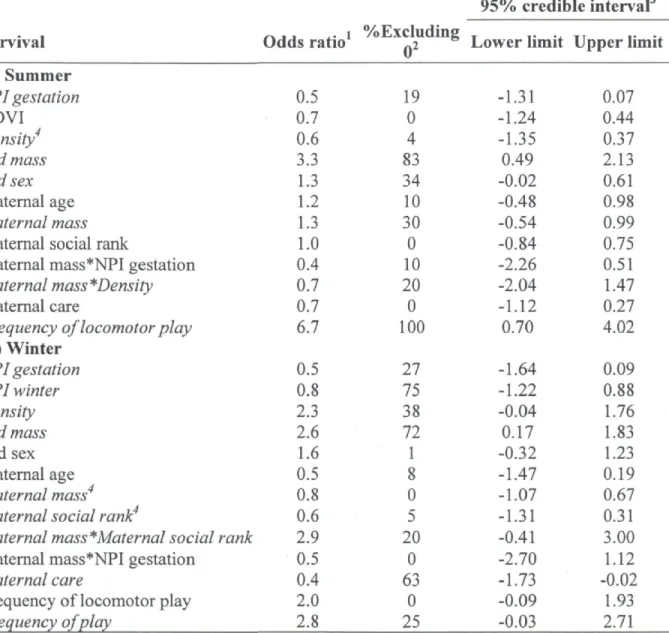
Alberta, Canada (1995-2010) 26 Summary of the generalized linear mixed models describing the
influence of maternal characteristics, maternal behaviours, environmental conditions, offspring characteristics, and kid development on a) summer and b) winter survival probabilities of mountain goat kids, at Caw Ridge, Alberta Canada
LISTE DES FIGURES
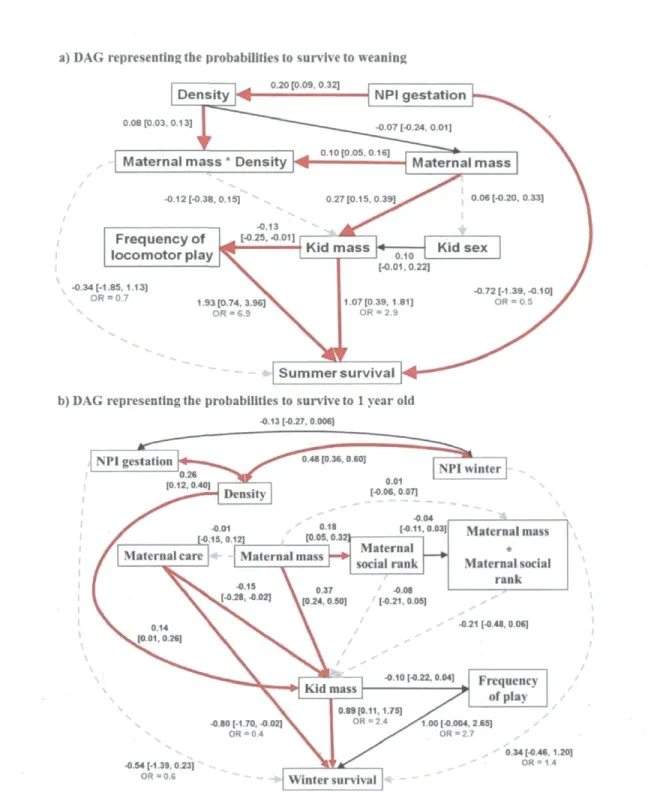
FIGURE 1 Path analysis models of the direct and indirect influence of environmental conditions, maternal characteristics, and kid's characteristics on a) summer survival and b) winter survival of mountain goat (Oreamnos americanus) kids at Caw Ridge, Alberta, Canada (1995-2010). Standardized partial-regression coefficients with their associated 95% credible intervals (CI) are presented above each path. Odds ratios (OR) are presented in gray for the variables having a direct effect on survival. Thick and red solid lines represent paths with good evidence of an effect (95% CI not overlapping zero), thin solid black lines indicate weak evidence for an effect (95% CI including marginally zero; < 0.05), and thin gray dotted lines stand for unclear relationships (95% CI including zero). Double-headed lines stand for correlations among independent variables that cannot be explain by causation. Female ID was included as a random
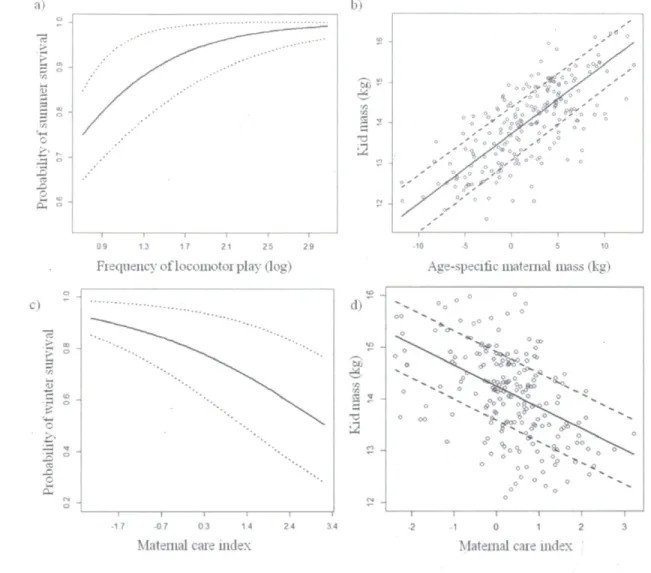
effect 33 FIGURE 2 Direct and indirect effects emphasised by the best DAG on survival
(left panels) and kid mass (right panels) of mountain goat (Oreamnos americanus) kids at Caw Ridge, Alberta, Canada (1995-2010). Influence of the frequency of locomotor play on kid summer survival (a), of maternal mass (corrected for maternal age) on kid mass (b), of maternal care index (length and frequency of suckling bout, frequency of suckling rejection, and mother-kid distances) on winter survival (c), and on kid mass (d). Continuous lines represent the predicted values from the mixed models, dotted lines are the 95% credible intervals, and dots are the observed
ANNEXE A
XI
LISTE DES ANNEXES
TABLE A.l Complete list of a priori generalized linear mixed models tested with their AICc for summer survival of mountain goat kids (n=267 kids), at Caw Ridge, Alberta, Canada
(1995-2010) 65 TABLE A.2 Complete list of a priori generalized linear mixed models
tested with their AICc for winter survival of mountain goat kids (n=267 kids), at Caw Ridge, Alberta, Canada
(1995-2010) 66
ANNEXE B
FIGURE B.lFIGURE B.2
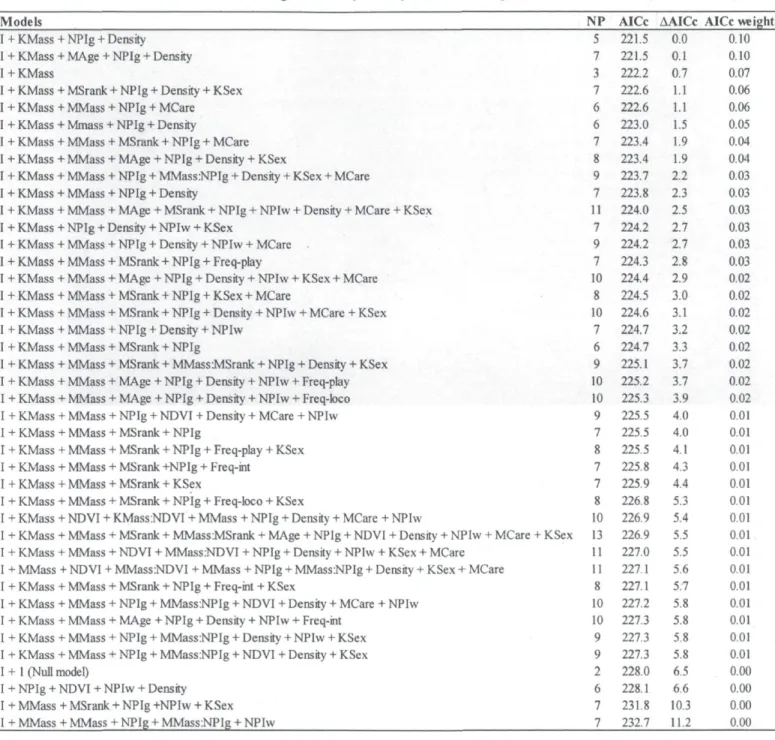
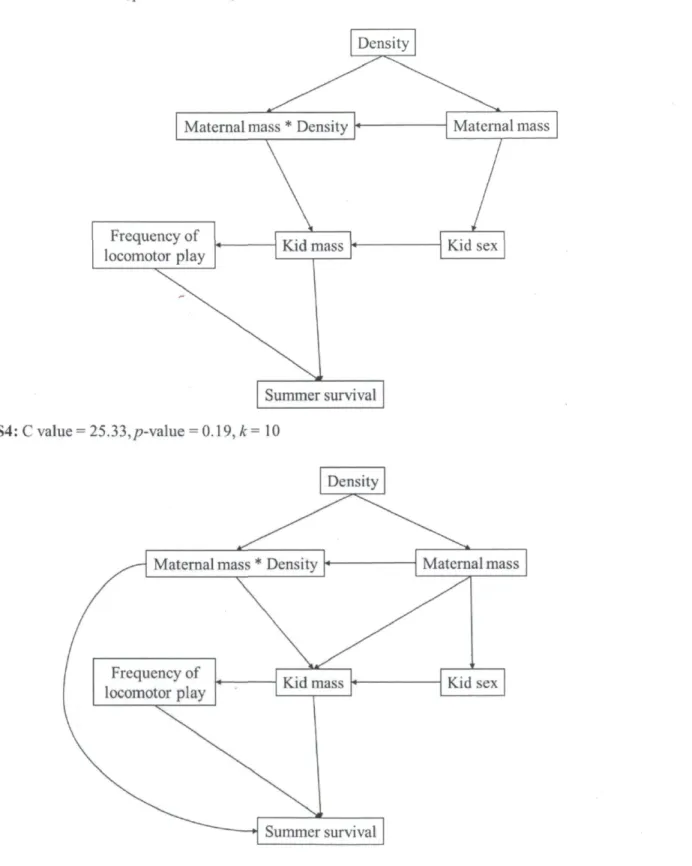
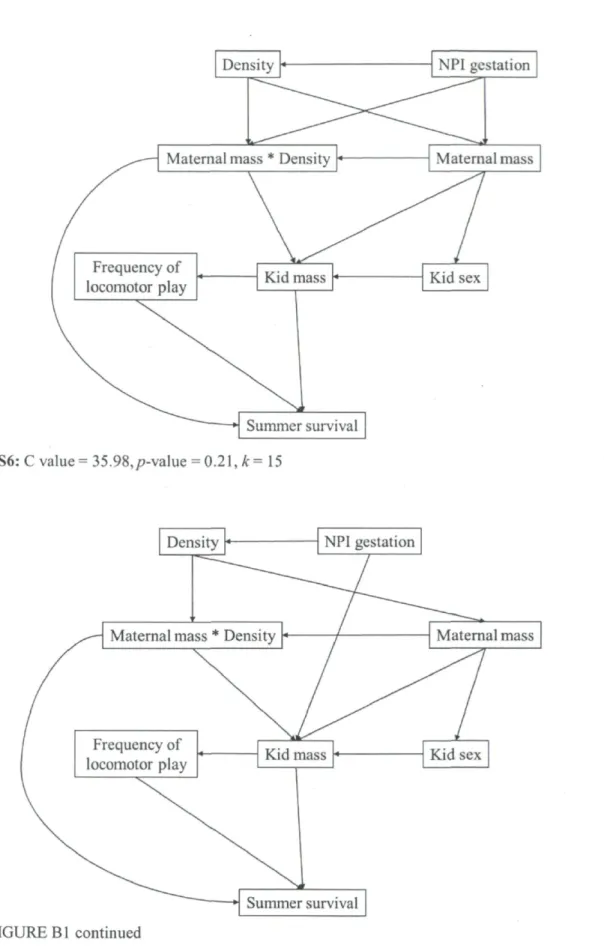
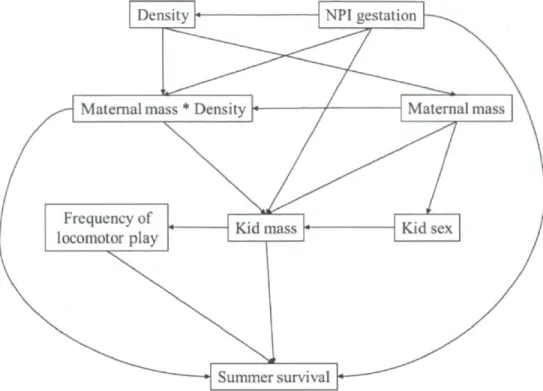
Causal models (S1-S9) tested to determine summer survival for mountain goat kids on Caw Ridge, Alberta, Canada (1995-2010). C values are the calculated combined values using Shipley's method (2002). P-value is obtained by comparing probabilities of the corresponding C value with k (number of d-sep claims) as degrees of freedom. Values of the model presenting the best fit are in bold. NPI= North Pacific Index. The figure spreads on the next four
pages 69 Causal models (WI- W19) tested to determine winter survival
for mountain goat kids on Caw Ridge, Alberta, Canada (1995-2010). C values are the calculated combined values using Shipley's method (2002). P-value is obtained by comparing probabilities of the corresponding C value with k (number of d-sep claims) as degrees of freedom. Values of the model presenting the best fit are in bold. NPI=North Pacific Index. The figure spreads on the next nine
CHAPITRE 1
CADRE THEORIQUE
Depuis la publication de «L'origine des espèces» de Darwin en 1859, les scientifiques se sont intéressés aux causes et aux conséquences des variations phénotypiques et génétiques entre les organismes. La biologie évolutive vise à comprendre l'importance des pressions de sélection sur l'évolution du phénotype amenant des différences dans les composantes biodémographiques caractérisant une espèce, une population ou même un individu (Thomas et al. 2010). Ces composantes biodémographiques, aussi appelées traits d'histoire de vie, sont le résultat de diverses contraintes auxquelles font face quotidiennement les organismes vivants (Stearns 1992). Ces contraintes, qui peuvent être de nature physiologique, génétique ou environnementale, surviennent principalement parce que les ressources disponibles sont limitées. Des compromis s'installent alors entre les différentes composantes biodémographiques car l'allocation d'énergie dans un trait se fait au dépend de l'énergie disponible pour un autre trait (Thomas et al. 2010). Les organismes sont ainsi amenés à évoluer vers diverses stratégies d'allocation d'énergie dépendamment des contraintes auxquelles ils sont soumis. Parmi ces contraintes, le compromis entre la valeur reproductive présente et les possibilités de reproduction futures est à la base de nombreuses stratégies évolutives telles le nombre et la taille de la progéniture produite, l'âge à la première reproduction ou la qualité des soins parentaux prodigués (Williams 1966). Une stratégie reproductive est donc sélectionnée en fonction des contraintes locales auxquelles les organismes font face. L'étude des causes et des conséquences de cette sélection sur les composantes biodémographiques permet une meilleure compréhension des stratégies évolutives d'une espèce ou d'une population.
Une des composantes biodémographiques directement liées à la stratégie reproductive est l'investissement dans la progéniture par les soins post-partum prodigués aux jeunes. L'apport de l'énergie nécessaire à leur croissance ainsi que la protection contre les prédateurs sont des soins ayant comme but ultime d'assurer la survie de cette progéniture (Clutton-Brock 1991). D'autres soins ayant des effets plus subtils tels l'inclusion sociale ou l'apprentissage des meilleures aires d'alimentation peuvent aussi
influencer la survie des jeunes (Bernardo 1996). Les jeunes qui survivent jusqu'à l'âge de la maturité peuvent ainsi se reproduire eux-mêmes et permettre la transmission des gènes parentaux dans la population, assurant par le fait même la valeur phénotypique ou adaptative (aussi appelée fitness) des parents (Clutton-Brock 1988). La quantité et la qualité des soins parentaux ayant un effet positif sur la survie des juvéniles seraient donc le résultat d'une sélection sous les différentes contraintes physiologiques, génétiques et environnementales auxquelles sont confrontés les parents et les jeunes (Trivers 1974, Clutton-Brock 1991). Une étude approfondie des facteurs influençant la probabilité de survie des jeunes peut ainsi amener une meilleure compréhension de l'évolution des soins parentaux. Leur efficacité à augmenter la probabilité de survie des jeunes révèle également leur potentiel adaptatif pour une espèce et même entre les différentes populations de cette même espèce faisant face à des contraintes différentes (Badyaev et Uller 2009).
La masse corporelle
Les études sur la survie, adulte ou juvénile, montrent souvent que les facteurs améliorant la probabilité de survie d'un individu agissent indirectement en favorisant l'augmentation de la masse corporelle (Guinness et al. 1978, Wauters et al. 1993, Festa-Bianchet et al. 1997, Boltnev et al. 1998). Par exemple, Telia et ses collègues (2000) ont montré chez les crécerelles d'Amérique (Falco sparverius) que les individus de masse corporelle supérieure présentaient une réponse immunologique aux infections plus efficace et rapide, ce qui les rendait plus résistants aux maladies et augmentaient ainsi leur chance de survie. Pour les espèces de milieux tempérés, des réserves de graisse suffisantes sont essentielles puisque les individus sont soumis à des périodes hivernales prolongées où l'accès à la nourriture est limité et les demandes énergétiques liées à la thermorégulation et aux déplacements sont accrues (Stevenson et Woods 2006, Taillon et al. 2006). Par conséquent, la sélection devrait favoriser une augmentation de la masse corporelle (revu dans Blanckenhorn 2000). Cependant, une masse corporelle trop élevée diminuera l'agilité des organismes face à un prédateur, par exemple, ou conduira à un taux de métabolisme basai plus élevé par une augmentation des demandes énergétiques. Ces coûts limiteront alors l'augmentation de la masse (Blanckenhorn 2000). La masse corporelle serait donc un
facteur ayant une forte influence sur les chances de survie (e.g., Ribble 1992, McMahon et al. 2000, Hall et al. 2001, Pettorelli et Durant 2007).
Les soins maternels
Puisque les chances de survie d'un individu sont en partie le résultat de l'interaction de plusieurs facteurs environnementaux, génétiques et phénotypiques influençant sa masse, l'exploration de ces facteurs devient également fondamentale afin étudier les déterminants de la survie juvénile (Hadley et al. 2007). Chez la plupart des mammifères et particulièrement chez les ongulés, les soins parentaux sont des comportements maternels qui ont un effet direct sur la masse et donc la survie des jeunes (Bernardo 1996). Étant donné que l'énergie nécessaire à la croissance du jeune jusqu'au sevrage vient presqu'exclusivement de la mère via la lactation, la quantité et la qualité du lait maternel sont alors parmi les déterminants principaux de la masse corporelle des jeunes (Jasper et Weary 2002).
En plus de l'allaitement, la mère prodigue aussi d'autres soins qui peuvent influencer plus subtilement la croissance et la survie de son jeune. Le lien unissant un nouveau-né à sa mère est essentiel à sa survie chez les mammifères (voir Nowak et al. 2000) et le maintien de ce lien est le résultat d'un équilibre entre les intérêts de la mère et ceux du jeune (Trivers 1974). Malgré la dépense énergétique encourue par la mère, sa protection et sa proximité permettent une meilleure défense contre les prédateurs et une réduction des interactions agressives provenant des autres femelles adultes (Green 1992a, Bernardo 1996). De plus, si son jeune est à proximité, la mère peut diminuer sa vigilance, donc elle profitera mieux des périodes d'approvisionnement tout en offrant une meilleure protection contre les prédateurs à son jeune (Barrett et al. 1995, May et al. 2008).
La quantité et la qualité des soins maternels nécessaires au bon développement du jeune seraient avant tout influencées par les caractéristiques maternelles (Green 1986,
affectent le coût de la reproduction (Hamel et al. 2009a) et la variation de ces coûts due à l'hétérogénéité des femelles devrait alors se refléter dans la qualité et la quantité de soins alloués aux jeunes. Les jeunes femelles ont un coût reproductif plus grand que les vieilles femelles puisqu'elles n'ont pas encore atteint leur masse asymptotique et doivent donc répartir leur énergie entre la reproduction et la croissance (Côté et Festa-Bianchet 2001a, Loison et al. 2004, Hamel et Côté 2009). D'un autre côté, les femelles âgées pourraient augmenter l'investissement accordé à la croissance de leur jeune puisque leur valeur reproductive résiduelle diminue avec l'âge (i.e. elles ont moins d'opportunités de reproduction futures; Williams 1966, Stearns 1992). Ainsi, l'âge de la mère et son rang social influencent les comportements d'allaitement chez le bison, Bison bison (Green 1986, 1990), chez l'arui, Ammotragus lervia (Cassinello 2001) et chez le mouflon de Corse, Ovis gmelini musimon (Réale et Boussès 1995). Green (1986, 1990) a notamment montré chez le bison que l'âge de la mère influençait positivement la durée totale des allaitements. Par contre, chez les autres espèces, un jeune né d'une mère âgée ou subordonnée avait moins de périodes d'allaitement; celles-ci étaient de plus courte durée et il consacrait une plus grande proportion de son budget d'activité à s'approvisionner (Réale et Boussès 1995, Cassinello 2001). Un meilleur accès aux lieux d'approvisionnement chez les femelles âgées et de rang social élevé devrait aussi leur permettre de sevrer des jeunes de plus grande masse et ayant une plus forte probabilité de survie (Curio 1983, Hadley et al. 2007, Solberg et al. 2007, Hamel et al. 2009b). Les relations sociales du jeune avec le reste du groupe aux sites d'alimentation peuvent être influencées par le rang social de sa mère (Weeks et al. 2000, Cassinello 2002). Les agneaux nés d'une brebis arui ayant un rang social plus élevé avaient un meilleur accès aux ressources et étaient moins dérangés par les autres adultes lors de l'approvisionnement (Cassinello 2002). En plus de l'expérience maternelle antérieure et du rang social, l'effort alloué à la lactation par les femelles varie également en fonction de leur condition corporelle. Étant donné qu'une femelle adulte de masse supérieure a plus de réserves énergétiques, elle peut produire du lait de meilleure qualité et en plus grande quantité (Landete-Castillejos et al. 2005). En retour, cela augmente les apports nutritifs de son jeune et ainsi sa croissance jusqu'au sevrage (Clutton-Brock et al. 1996, Bowen et al. 2001). Dans l'ensemble, la compréhension des contraintes et des effets des soins maternels est essentielle à l'étude de la variabilité de la masse et de la survie des jeunes.
Le sexe
Chez les espèces présentant un dimorphisme sexuel important, le sexe est un facteur clé dans la détermination des patrons de croissance. Chez les ongulés, les mâles ont généralement évolué vers une croissance plus rapide et plus importante que celle des femelles puisque leur succès reproducteur est souvent relié à leur masse et à leurs habiletés au combat (Trivers et Willard 1973, Festa-Bianchet et al. 1996, Birgersson et Ekvall 1997, Cassinello 2001). En considérant que la masse corporelle a plus d'effet sur le succès reproducteur d'un mâle que sur celui d'une femelle chez les espèces polygynes avec dimorphisme sexuel, une mère augmenterait sa valeur adaptative en investissant plus d'énergie dans la croissance de ses fils que celle de ses filles (Trivers et Willard 1973). Le sexe du jeune peut donc influencer les soins maternels que le jeune reçoit, affectant ainsi ses apports et ses dépenses énergétiques, et de fait sa croissance et sa survie. Chez certaines espèces comme la chèvre marronne de la Nouvelle-Zélande, Capra hircus (Alley et al. 1995) et l'antilope d'Amérique, Antilocapra americana (Byers et Moodie 1990), les fréquences d'allaitement et les budgets d'activité des jeunes varient selon leur sexe. Le sexe peut également influencer les comportements du jeune et ainsi modifier le développement de sa motricité et ultimement sa survie. Chez le cheval sauvage, Equus caballus, les mâles passent une plus grande proportion de leur temps actifs que les femelles, augmentant alors leurs demandes énergétiques et leur vulnérabilité aux blessures ou aux prédateurs (Duncan et al. 1984). En résumé, un dimorphisme sexuel marqué est généralement caractérisé par un patron de croissance et de survie différent entre les sexes.
Les facteurs environnementaux
La survie des jeunes d'une même cohorte peut être affectée par différents effets environnementaux, tels la présence de prédateurs ou la densité de la population (Albon et al. 1987, Couturier et al. 2009). La densité de la population qui influence la qualité de l'habitat affecte ultimement la croissance des jeunes en modifiant la qualité et la quantité de nourriture disponible (Albon et al. 1987, Singer et al. 1997, Festa-Bianchet et al. 1998).
8
D'autre part, la présence de prédateurs ou d'activité anthropique peuvent changer les patrons de mouvements et d'utilisation des habitats, ce qui peut entraîner des dépenses énergétiques additionnelles (Cari et Robbins 1988, Lima 1998, Mullner et al. 2004, Rode et al. 2007). Les facteurs environnementaux influençant la masse et la survie des jeunes résultent également du climat (Gaillard et al. 1997, Couturier et al. 2009). Les conditions climatiques peuvent grandement affecter les apports énergétiques de la mère et du nouveau-né, ce qui aura une grande influence sur les chances de survie de ce dernier (Loison et al. 2004, Pettorelli et al. 2007). Lors de la croissance, un habitat de meilleure qualité suite à un climat favorable augmente généralement le gain en masse des jeunes herbivores en leur donnant accès à une source de nourriture plus abondante et de valeur nutritive supérieure (Langvatn et al. 1996, Saether et al. 1996, Pettorelli et al. 2007, Couturier et al. 2009). Les bénéfices d'une bonne qualité d'habitat se répercuteront également de façon positive sur la condition corporelle de la mère qui pourrait alors produire plus de lait et de meilleure qualité (Landete-Castillejos et al. 2005). Un climat défavorable, par contre, peut entrainer une augmentation des dépenses énergétiques et une diminution des ressources disponibles, ce qui, chez la femelle gestante, pourrait amener une diminution de l'énergie disponible à la croissance du foetus et à la production de lait par la suite (Saether et al. 1996, Weladji et Holand 2003). Puisque ces contraintes augmentent les coûts de reproduction chez les espèces adoptant une stratégie conservatrice, la femelle favorisera sa propre survie et diminuera l'effort investi à la croissance et à la survie de son jeune (Festa-Bianchet et Jorgenson 1998). Les périodes d'allaitement sont de plus courte durée et moins fréquentes lors des années où la densité de la population de mouflons, Ovis musimon, est élevée et la qualité de l'habitat se détériore (Réale et Boussès 1995, Réale et al. 1999). Chez les bouquetins, Capra ibex, les femelles prolongent la période de lactation de leur jeune et modifient leurs comportements maternels durant les années de sécheresse (Grignolio et al. 2007). Une influence directe des conditions environnementales sur la survie des jeunes peut alors être en combinaison avec des changements comportementaux de la mère qui influencent les patrons de croissance et de survie des jeunes.
OBJECTIFS GENERAUX ET SPÉCIFIQUES
Dans le cadre général de l'évolution des soins parentaux et de leur efficacité à augmenter les probabilités de survie des jeunes, mon projet de maîtrise visait à identifier les facteurs influençant la masse corporelle et la survie des jeunes chez la chèvre de montagne, Oreamnos americanus. À l'aide de données longitudinales sur une population naturelle marquée, mon travail avait pour but de déterminer l'influence relative i- des conditions environnementales, ii- des caractéristiques du jeune, iii- des caractéristiques de la mère, iv-des soins maternels, et v- du développement du jeune sur la survie iv-des chevreaux jusqu'au sevrage (estivale) et jusqu'à un an (hivernale). Parmi les déterminants les plus importants de chacune des périodes, nous avons ensuite examiné les relations causales entre ces facteurs (effets directs ou indirects) à l'aide de modèles de pistes.
LA CHÈVRE DE MONTAGNE
La chèvre de montagne est un ongulé des milieux alpins. Elle présente plusieurs caractéristiques permettant d'évaluer les facteurs influençant la probabilité de survie des jeunes ainsi que l'effet des soins maternels sur la survie. L'habitat montagnard des chèvres se prête particulièrement à l'étude de l'effet des conditions environnementales sur la croissance et la survie des jeunes puisque ce milieu est caractérisé par une saison végétative courte, suivie d'une saison hivernale longue et rigoureuse. De plus, les chèvres de montagne sont présentes dans des habitats peu productifs et relativement isolés d'une population à l'autre, alors la variabilité annuelle des taux de survie des chevreaux pourrait aussi être en partie expliquée par la densité de la population qui influencerait la qualité de l'habitat.
Durant la lactation qui s'étend entre mai et septembre, la mère est le seul parent en charge de l'apport régulier d'énergie au chevreau. Les soins parentaux alloués au jeune lors de sa croissance sont donc strictement maternels. En plus, puisque les femelles adultes ne produisent qu'un seul chevreau par année et qu'il n'y a pas d'allaitement alloparental,
10
l'ensemble des soins maternels est donc entièrement alloué au seul chevreau. Côté et Festa-Bianchet (2001a, 2001b) ont démontré une influence positive de la masse maternelle sur la masse et la survie du jeune pour la même population de chèvres. Par contre, ils n'ont détecté aucun effet direct du rang social et de l'âge de la mère sur la masse corporelle et la probabilité de survie des chevreaux. Étant donné que les caractéristiques maternelles ont le potentiel d'interagir avec les facteurs environnementaux, les soins maternels et les caractéristiques du jeune, affectant ainsi la probabilité de survie des jeunes, il est essentiel d'évaluer à nouveau leur influence en combinaison avec ces autres déterminants. En effet, puisque les coûts engendrés par la lactation peuvent varier grandement chez les chèvres de montagne selon l'âge, le rang social et la condition corporelle de la femelle adulte (Hamel et al. 2009b), ces caractéristiques maternelles pourraient en retour modifier les comportements de la femelle envers son jeune. De plus, la chèvre de montagne semble suivre les prémisses de l'hypothèse de Trivers et Willard (1973) sur l'allocation des soins parentaux : condition maternelle positivement corrélée avec celle du jeune au sevrage (Côté et Festa-Bianchet 2001c), caractéristiques juvéniles récurrentes à l'âge adulte (Gendreau et al. 2005), et une petite différence de masse a un effet plus marquée sur le succès reproducteur des mâles que sur celui des femelles (Hewison et Gaillard 1999, Mainguy et al. 2008, Mainguy et al. 2009). Effectivement, la proportion supérieure de chevreaux femelles produite par les jeunes femelles (70% comparativement à 25% pour les femelles plus âgées; Côté et Festa-Bianchet 2001c), suppose une différence dans l'allocation des ressources selon le sexe du chevreau et les caractéristiques de la mère. Finalement, la naissance des chevreaux est très synchronisée à la mi-mai et 80% des jeunes naissent à l'intérieur d'une période de deux semaines (Côté et Festa-Bianchet 2001a), ce qui permet une comparaison valable des comportements maternels et des patrons de croissance des jeunes.
11
L'AIRE D'ETUDE
Nous avons étudié la survie des chevreaux d'une population de chèvres de montagne, située en Alberta, Canada, sur une période de 15 ans. L'aire d'étude se trouve en milieux alpin et subalpin aux piémonts de la chaîne des montagnes Rocheuses. Cette population de chèvres occupe une aire d'environ 28 km2 variant en altitude de 1750 à 2170 mètres. Au bas de ce
gradient altitudinal, à 1900 mètres, le milieu est caractérisé par de la forêt subalpine majoritairement composée d'épinette d'Engelmann (Picea engelmanniï). Les sommets de toundra alpine offrent une végétation comprenant principalement des graminées, des cypéracées et différentes plantes herbacées. En plus de ces habitats, les chèvres utilisent les falaises et les éboulements rocheux en guise de terrains de fuite pour échapper aux prédateurs (e.g., ours grizzly, Ursus arctos, le loup, Canis lupus et le couguar, Puma concolor; Festa-Bianchet et al. 1994). La chasse sportive à la chèvre de montagne y est interdite depuis 1969. Par contre, l'accès à l'aire d'étude pour fins récréatives n'y est pas restreint. Le climat montagnard peut amener des températures sous zéro degré Celsius ainsi que des précipitations de neige tout au long de l'année. Les hivers sont longs et rigoureux, les étés sont courts et frais. Les chèvres disposent donc d'une saison végétative très courte et la période où l'alimentation est de très bonne qualité est encore plus brève.
Capture et marquage
L'étude de cette population a commencé en 1989. Depuis 1993, au moins 98% des individus d'un an et plus sont marqués à l'aide de colliers et d'étiquettes apposées aux oreilles (Festa-Bianchet et Côté 2008). Les chèvres sont appâtées au lieu de capture du mois de juin jusqu'à la fin du mois de septembre à l'aide de blocs de sel. Nous utilisons des trappes de type Clover et Stephenson afin de capturer les individus non-marqués (Côté et al. 1998). La manipulation des chèvres âgées de plus de deux ans nécessite une immobilisation chimique à l'aide de Xylazine® suivie d'une injection d'Idazoxan®, utilisée pour contrer les effets du tranquillisant à la fin de la capture (Haviernick et al.
12
par la mère suivant la capture du jeune (Côté et al 1998). Les jeunes ayant été capturés puis abandonnés ont été retirés des analyses puisque le sevrage précoce peut influencer leur probabilité de survie. Une fois capturée, plusieurs mesures morphologiques sont prises sur la chèvre telle la masse, la longueur des pattes, la longueur et la largeur de la base des cornes ainsi que la circonférence du torse. L'âge des individus est évaluée en comptant les anneaux de croissance sur les cornes, une méthode considérée fiable jusqu'à l'âge d'environ sept ans (Stevens et Houston 1989). Afin d'effectuer des observations comportementales, les femelles, à partir de leur deuxième année, sont munies d'un collier visuel permettant leur reconnaissance à distance. Les mâles sont équipés de colliers émetteurs de type VHF et des étiquettes de couleur leur sont apposées aux oreilles. Des échantillons de peau et de poil sont recueillis afin de confirmer génétiquement la parenté des juvéniles âgés d'un an. En plus des trappes, le site de capture est pourvu de trois balances électroniques à plateau permettant une lecture à distance. Une grande quantité de masses a donc été recueillie depuis 2001 sans que les individus ne soient manipulés.
LES OBSERVATIONS COMPORTEMENTALES
Les observations comportementales sont réalisées à distance à l'aide de jumelles ou de télescopes (15x-45x). La taille, l'emplacement sur l'aire d'étude et l'identification des individus composant les groupes de chèvres sont notés quotidiennement. Le statut reproducteur de chaque femelle adulte est évalué selon la présence d'un chevreau avec elle. Le sexe du chevreau est jugé par sa position lorsqu'il urine ou par l'observation de la région anogénitale lors des allaitements. Des observations cibles de deux heures sont effectuées sur les dyades chevreau-mère. Durant ces observations, l'activité de la mère et du jeune est notée à intervalle régulier d'une minute. Les classes d'activité retenues pour cette étude sont l'approvisionnement, le déplacement, les comportements sociaux, le repos, le sommeil/rumination et les indéfinis. À chaque deux minutes, la distance entre les deux membres de la dyade ainsi que leur mouvement respectif par rapport à l'autre individu (immobile, suit, est suivi) sont enregistrés. Les possibilités d'interactions sociales sont notées dès qu'un individu se trouve dans un rayon de quatre mètres du jeune; ce qui est considéré comme une opportunité d'interaction (Côté 2000). Lorsqu'une interaction se
13
produit, l'individu responsable de l'initiation et celui qui remporte l'interaction sont enregistrés. Cela permet d'obtenir la fréquence des interactions agressives dirigées vers le chevreau pendant la période échantillonnée. Finalement, le rang social des femelles adultes est évalué par des observations répétées d'interactions agressives récoltées de façon ad libitum (Altmann 1974). Les interactions sont enregistrées de la même façon que lors des observations cibles des chevreaux, c'est-à-dire que l'initiateur et le gagnant de l'interaction sont notés. Une matrice de dominance est ainsi élaborée en combinant le nombre d'interactions gagnées, le nombre d'interactions perdues et l'identité des individus impliqués dans ces interactions. La hiérarchie sociale peut alors être établie en ordonnant les femelles en fonction du nombre d'interactions remportées. Finalement, la durée des tétées et l'individu responsable de l'initiation et de la terminaison de celles-ci sont enregistrés lors des observations cibles. La durée d'allaitement est calculée en secondes à partir du moment où le museau du chevreau entre en contact avec le pis de la mère jusqu'à la fin de ce contact. Un allaitement de moins de trois secondes où la mère s'éloigne ou empêche le jeune de téter est enregistré comme une tentative (allaitement refusé). Cela permet d'obtenir la fréquence des allaitements rejetés pour la période de l'observation. Nous utiliserons les comportements d'allaitement comme indicateurs de satisfaction du jeune plutôt que comme mesure indirecte de la quantité réelle de lait ingérée par le jeune,
(Mendl et Paul 1989, Cameron 1998, Therrien et al. 2008). Un transfert de lait inefficace ou de moindre qualité forcerait alors un jeune à téter plus longtemps et plus souvent afin d'atteindre la satiété, et se manifesterait par une diminution de la croissance (Therrien et al. 2008).
Des données sur ces diverses variables ont été récoltées à tous les étés entre 1995 et 2010. Au total, 267 chevreaux ont été suivis en observation cible pour une durée d'environ 120 minutes. Nous avons également enregistré la masse corporelle de 118 de ces chevreaux. Pour les 15 années de suivi, un total de 756 heures d'observations cibles a été amassé. Finalement, grâce au suivi quotidien des individus marqués et aux analyses génétiques confirmant l'identité de la mère, une donnée de survie est disponible pour tous les chevreaux étudiés depuis 1995.
CHAPITRE 2
Why care? The role of maternal behaviour and offspring
development in the survival of mountain goat kids
Rachel Théoret-Gosselin ' , Sandra Hamel and Steeve D. Côté
1. Department of biology, Université Laval, Québec, Canada 2. Centre d'études nordiques, Québec, Canada
RESUME
Les études de la survie juvénile mettent généralement l'emphase sur les effets des conditions environnementales et des caractéristiques maternelles. Par contre, la capacité des parents à donner des soins ainsi que le développement locomoteur des jeunes pourraient jouer un rôle déterminant dans la survie de la progéniture. Nous avons étudié l'influence
/-des conditions environnementales, //-/-des caractéristiques du jeune, iii- /-des traits maternels, iv- des soins maternels, et v- du développement du jeune sur les chances de survie jusqu'au sevrage et jusqu'à un an chez la chèvre de montagne (Oreamnos americanus). Le développement locomoteur sous forme de jeux et les soins maternels influençaient directement la survie des jeunes, et ce de façon plus importante que les conditions environnementales et les traits de la mère. La fréquence de jeu du jeune était corrélée positivement à ses chances de survie jusqu'au sevrage. Les soins maternels augmentaient la probabilité de survie à l'hiver, directement mais aussi indirectement par un effet sur la masse du jeune. La masse du chevreau était également un des deux déterminants influençant le plus la survie à l'été et à l'hiver. Les conditions environnementales influençaient directement la survie estivale tandis que les caractéristiques maternelles affectaient indirectement la survie hivernale par un effet sur la masse du jeune. Les adaptations comportementales des soins parentaux et du développement juvénile aux pressions de sélection locales peuvent amener des adaptations locales et pourraient avoir de plus grandes implications dans la dynamique des populations qu'anticipé.
ABSTRACT
Studies on juvenile survival have mainly focused on the effects of environmental conditions and maternal traits. However, growing evidence indicates that the ability of parents to care for their young and the offspring developmental behaviours could be key determinants of their survival. We examined the relative influence of i- environmental conditions, ii- offspring characteristics, iii- maternal characteristics, iv- maternal care behaviours, and v- offspring developmental behaviours on kid survival to weaning and to 1 year old in mountain goats (Oreamnos americanus). Offspring development and maternal care affected directly offspring survival and this, more importantly than environmental conditions and maternal traits. Frequency of play strongly increased survival before weaning. Greater maternal care increased offspring survival during winter, directly and indirectly through kid mass. Kid mass was also a major determinant of both summer and winter survival. Environmental conditions mainly influenced directly summer survival while maternal characteristics affected indirectly winter survival through an effect on kid mass. Behavioural adaptation of maternal care and offspring development to local selective pressures can lead to local adaptations and have greater implications in population dynamic studies than previously believed.
19
INTRODUCTION
Variation in parental allocation can profoundly influence fitness. Although the ability of parents to care could be a key determinant of offspring survival (Green 1990, Bernardo 1996, Mousseau and Fox 1998, Réale et al. 2000, Therrien et al. 2008), the influence of reproductive allocation on offspring survival is often neglected in life-history studies. In most polygynous mammals, parental care is given strictly by females, and lactation is the most important caring behaviour providing the main source of energy for offspring early in the development (Oftedal 1985). In addition, maternal efforts to maintain proximity with her young and the presence of an adequate social environment for locomotor development can decrease prédation risk and concomitantly enhance offspring survival (Nowak et al. 2000). Offspring social and locomotor development may also strengthen the mother-young bond and provide juveniles with abilities to cope with stressful situations (Spinka et al. 2001, Fagen and Fagen 2004). Regardless of the increasing evidence that maternal reproductive effort may affect juvenile life-history traits (Bernardo 1996, Mousseau and Fox 1998, Marshall and Uller 2007, Festa-Bianchet and Côté 2008, Mousseau et al. 2009), the ultimate impacts of maternal care and offspring development on juvenile survival remain overlooked. Indeed, most studies assessing the determinants of offspring survival have focused on environmental factors, offspring characteristics, and maternal traits. However, the relationships between maternal traits, offspring condition and survival are likely explained by maternal care, which thus be considered in such studies (Andersen et al. 2000). Researches focusing on the relationships between maternal care and offspring development are required to disentangle the direct and indirect determinants of juvenile survival, a fundamental component for understanding population fluctuations (Gaillard et al. 1998, Gaillard étal. 2000).
Several studies have revealed the influence of environmental conditions and offspring characteristics on juvenile survival. In large herbivores, environmental conditions associated with access to high quality forage, such as a gradual green-up of vegetation in temperate ecosystems, high habitat productivity, or low population density, can enhance
20
juvenile growth and survival (Guinness et al. 1978, Festa-Bianchet 1988, Coulson et al. 2001, Pettorelli et al. 2007, Couturier et al. 2009). Other environmental factors, like prédation risk or snow depth, influence habitat selection and movement patterns, and can therefore affect juvenile growth through impacts on energy expenditure and forage availability (Frid and Dill 2002). In general, favourable environmental conditions increase juvenile survival through an indirect positive influence on offspring traits such as enhanced body growth, fat reserves, and mass (Gaillard et al. 1997, Loison et al. 1999); larger offspring are thus more likely to survive than lighter ones (e.g. McMahon et al. 2000, Taillon et al. 2006). Moreover, direct and indirect effects of environmental conditions on offspring traits and juvenile survival can vary with sex in dimorphic species, as demonstrated in red deer (Cervus elephus) where males had to be larger to achieve the same survival probabilities as females during winter (Loison et al. 1999). Environmental conditions can also have an indirect impact on offspring survival through their influence on maternal condition (Festa-Bianchet and Jorgenson 1998).
Maternal traits can determine offspring mass gain and survival (Bernardo 1996, Solberg et al. 2007, Landete-Castillejos et al. 2009). Females in poor nutritional condition and with low body fat reserves generally give birth to light young and are potentially unable to satisfy their progeny's nutritional needs because they produce less or low quality milk (Landete-Castillejos et al. 2005, Lomas and Bender 2007). Offspring cared by older females may have higher survival because older mothers are likely to provide greater allocation than younger mothers due to their increased reproductive experience or reduced reproductive value (Pianka and Parker 1975, Daunt et al. 2007, Solberg et al. 2007, Meijer et al. 2011). Other maternal traits, such as social rank or home range quality, can also enhance juvenile survival through access to high quality forage and reduced vulnerability to prédation (Clutton-Brock et al. 1984, Van Moorter et al. 2009). Because offspring depend mostly on maternal care before weaning (Nowak et al. 2000), maternal traits could also indirectly affect juvenile survival through their effects on the quantity and quality of care provided.
21 We assessed the summer and winter survival probabilities of mountain goat kids (Oreamnos americanus) using 15 years of longitudinal data from marked individuals. Our objectives were to determine the relative influence of /- environmental conditions, //-offspring characteristics, iii- maternal characteristics, i maternal care behaviours, and v-offspring developmental behaviours on kid survival to weaning (summer survival) and to one year old (winter survival). Using the most influential factors revealed in a first set of regression analyses, we further examined with path analyses the causal patterns, i.e. direct or indirect effects, of these variables and how they interacted among each other to influence kid survival. We expected environmental conditions and maternal characteristics to affect kid survival, but only indirectly through their influence on kid mass. We hypothesized that greater maternal care would enhance summer and winter survival, both indirectly, through increased energy allocation enhancing offspring mass, and directly, through reduced prédation risk associated with the maintenance of proximity. We expected heavy, old, and dominant females to provide greater maternal care than light, young, and subordinate mothers. In addition, we expected offspring locomotor development to influence survival positively over winter because of improved muscular development and capacity to cope with stressful situations (Spinka et al. 2001).
METHODS
Study area and captures
We studied mountain goats at Caw Ridge (54°N, 119°W), west central Alberta, Canada. Goats used an area of approximately 28 km of alpine tundra and open subalpine forest of Engelmann spruce (Picea engelmanii) at 1,750 to 2,170 m in altitude. The climate is subarctic-arctic, characterised by short, cool summers followed by long, harsh winters. The main predators of mountain goats in the study area are wolves (Canis lupus), grizzly bears (Ursus arctos), and cougars (Puma concolor). Black bears (U. americanus), coyotes (C. latrans), wolverines (Gulo gulo), and golden eagles (Aquila chrysaetos) may also predate on mountain goats (Festa-Bianchet and Côté 2008).
22
We used longitudinal data from marked females with a kid at heel from 1995 to 2010. Since 1988, we have captured goats using remotely-controlled box traps and self-tripping Clover traps baited with salt, and marked them with coloured ear tags and visual collars. Since 1993, 98% of goats aged 1 year or older have been marked. For adults not marked as yearlings, we determined their age by counting the number of horn annuli, a method reliable up to 7 years of age (Stevens and Houston 1989). Since 1998, we have not captured any adult female with a kid at heel, or any kid, to avoid kid abandonment (Côté et al. 1998). We excluded data from all mothers and kids captured in earlier years to avoid potential bias.
Survival
From mid-May to mid-September, we observed goats almost daily with spotting scopes (15x-45x). Mother identity of the kids was determined from observation of nursing behaviours and later confirmed using genetic analyses. We determined kid sex by the urination posture and the observation of the black vulvar patch for females (Côté and Festa-Bianchet 2001b). No kid was ever missing for more than one week and subsequently re-sighted, thus we considered kids that disappeared for seven days as dead. Kidding period was highly synchronised and occurred mainly between 20 May and 3 June (Côté and Festa-Bianchet 2001b). We defined kid summer survival as survival until weaning, September 15 (Côté and Festa-Bianchet 2001b). We considered that a kid survived over the winter when it was seen after June 1st the following year. We determined the identity of kids that
survived over the winter from observations of close associations with their mothers in the spring (Gendreau et al. 2005) and from DNA analyses of skin samples collected during marking (Mainguy 2008).
Environmental conditions
To characterize winter conditions, we used the November to March anomalies of the North Pacific Index (NPI; Trenberth and Hurrell 1994). For our study site, high values of NPI
23 represent snowy and cold winters and low values correspond to less snowy and mild winters (Hamel et al. 2009b). We used two indices: one representing winter conditions during the pre-natal period (NPI gestation) and the other describing winter conditions during the first winter of the kid (NPI winter). To characterize annual variation in vegetation availability in the spring, we used the sum of the bimonthly Normalized Difference Vegetation Index (NDVI; Pettorelli et al. 2005) of June. At Caw Ridge, high NDVI values correspond to rapid spring green-up, representing early access to vegetation (Hamel et al. 2009c). We used the total number of goats in the population on June 1st as a
measure of population density.
Masses
We recorded masses (± 0.5 kg) with three remotely-controlled electronic platform scales placed at the trap site and baited with salt. We could therefore weigh many individuals, especially kids, without handling them. Because we recorded several masses throughout the summer for the same individual, we adjusted mother masses to July 15l using the average
daily summer mass gain for five age classes of females (3, 4, 5, 6 and >7 years old; see Festa-Bianchet et al. 1996, Hamel and Côté 2009, Hamel et al. 2010). Because mass increased with age until approximately 7 years old (Côté and Festa-Bianchet 2001a), we calculated age-specific masses as the residuals of the cubic regression of mass on age (Mainguy and Côté 2008). We also adjusted kid masses to July 15th using the average
summer mass gain for each sex (males: 217.0±9.3 g/day; females: 202.9±8.4 g/day), which were obtained from a linear mixed model with mother identity as a random intercept (Festa-Bianchet and Côté 2008). Further description of capture and marking techniques can be found in Côté et al (1998).
Behavioural observations
From 1995 to 2010, we recorded 483 focal observations on 267 mother-kid pairs (from 1 to 6 observations per pair; Altmann 1974). From the observed dyads, 209 kids survived to
24
weaning (78%, ranging from 55% to 100% each year). Out of the kids that survived over the summer, 162 survived the winter (78%, ranging from 50% to 100%). We followed each dyad for a minimum of 30 subsequent minutes to a maximum of 120 minutes, for a total of 756 hours of observations. Every two minutes, we recorded the estimated distance between the dyad members and their respective direction of movement according to the other's position: approaching (moving towards the other), leaving (moving away), or neutral (Hinde and Atkinson 1970). During the entire focal observation, we recorded the identity of any individual observed within 4 m of the kid (considered as an opportunity for interaction; Côté 2000) and noted when an aggressive interaction towards the kid occurred. Every time the kid's nuzzle touched its mother's udder, we recorded the duration of the contact in seconds with stopwatches (±ls). These behavioural observations were used to define eight variables for characterizing maternal care and kid development processes (Table 1). We log-transformed these variables because they all followed a Poisson distribution. We used frequency of locomotor play, frequency of play behaviour, and frequency of social interaction to evaluate offspring physical and social development. To define maternal care quantity and quality, we used an index that combined a mother's nursing efforts with her efforts to maintain proximity to her kid (hereafter called maternal care). We performed a principal component analysis (PCA) on the variables: dyad distance, contact-maintenance index, suckling frequency, total suckling duration, and frequency of suckling rejection. The first axis, explaining 41% of the variance, was the only influential axis of the PCA (as determined by the rapid decrease in consecutive eigenvalues; Cattel 1966). ). We therefore used the scores of the first axis as an index of maternal care, where positive scores represented kids that nursed frequently (eigenvector^.62), for a long total duration (0.63), but stayed at closer distance to their mother (-0.44) which regularly rejected their suckling attempts (0.22), whereas negative scores characterized kids that were nursed less frequently, for a shorter total duration, and stayed at longer distance to their mother which seldom rejecting their suckling attempts. The scores were negatively correlated with summer day (r = -0.63). To control for variation in maternal care over the summer, we adjusted these scores to July 15l using a regression equation (intercept [±SE] = 6.54 ±
0.37, slope = -1.63 ± 0.09) obtained from a linear model where the date was log-transformed.
25
We determined the social rank of each mother annually based on ad libitum observations of agonistic interactions recorded among adult females over summer. Because relationships were highly linear every year, we applied the method of de Vries (1998) using Matman 1.0 for Windows (Noldus Information Technology 1998) to order adult females in an annual hierarchy (see Côté 2000 for details). Since social rank was strongly correlated with age (r > 0.9; Côté 2000), we used the residuals of a polynomial regression of rank on age to obtain age-specific social ranks for each year.
26 o o ■> ( N JS i X c> OJ - N „ ^ ■s x x o rt _ c LO rr} U ' X a 2- t_ 'C CJ o X) LO Q < u . — i se W •a J Di CÛ ^ < ^ h- u - r t O
as
* § C < iS 3 O w rt -*—* o x - i (U JS BO C o rt O O o O « ^d c « _L X » *-■ 4 . > . X - ^ _3et
!§£
2 | c -ag-ë
S .9 g -o j ■*—' oo .2 S •-> .B rt rt <u 2. T" C «H S o Ë}§
H > 81'S
a j rt _c S Hr
a._t
rt X J O X J X X J X X o X X o O - C X J > ï X T 3 . 2 T3 .t_ «i.s
1 S u -a _g (IJ o c c 'rti
o rt +_ C O O X • 1-H 3 0 0-73 C SJ cl C ___ a "u w 0 Q 0 S (U > CJ en -a o - c X J -a (U o E <u ■5 c <D CJ > 0 u 0 X J X is
.2 ' X OJ CJ "eo jgg
CJ .o X J c u 4 J LO xi rt O Ë 9 >- u • — C > -a CJI
u. ■■s1
C N o o CM S .o o -4-1 ^ - " > f _ £> 1 -c E G CJ C — '— C1
. OJ o o o CS ' C J <o •S >% -° t. u o -rt X -rt C*J.I
E E <- fe O o • _ TJ 5 o ■S S'1 'É
i l
'i 1
X ca O a iH o —, o • — ix o -y >-, -2 a. x x O >> CJ c o 3 cr CJ Ix U . o o o (N <o OJ X X J > ï X -rt «J -rt > • 1—1 - O ■a15
CJ X X J -rt c . o "3 o -O J3 rt t --^ w^ô
•O w 2_ .2 fe w JJ 0_ o i CJ8
.s f
0 O _5 •-111
Z s1
o s». o c 3 cr u. 3 o X OJ Cu C (U > (U G CJ 3 M U -O k> U X £ 3 c CJ 3 LO '*-O >. CJ c OJ 3 a-OJ 0 0 0 0 0 0 0 0 CN CN 1 — 4 53 X J rrS OJ C c (U OJ XÉ
o CJ CJ LO 0 0 .g13
CJ 3 LO C >. J 3 CTI
U x5 x -a CJ c o I -3 -a 130 _g13
CJ 3 LO OJ c o 3 - 3 00 _g13
CJ 3 LO "rt X J O l x 3 O X CJ C UJ C ^ O o (U 'ÔT' DXJ C13
CJ 3 LO 'xx O CJ X 3 Z c o ' X o a j 'âl? Ix 60 C rt x o 3 LO ' x -O > ï CJ c CJ 3 CT CJ27
Statistical analyses
The statistical analyses were carried out in two steps. First, we evaluated which of all the explanatory variables influenced kid survival over the summer and over the winter, using generalized linear mixed models (GLMMs). In a second step, we used path analyses to assess the structural or causal relationships among the predictors selected in the first step. We standardized all continuous variables to allow interpreting the influence of each predictor compared with one another (Schielzeth 2010). Because our models included a binary predictor (kid sex), we standardized variables by dividing the continuous variables by two standard deviations (Gelman 2008). We also centered independent variables to remove the collinearity between the main effects and the interactions, therefore allowing us to correctly execute model averaging (see below; Schielzeth 2010). We also tested for multicollinearity among the variables to avoid modeling problems. Furthermore, because our data set included some missing data in several explanatory variables, we used multiple imputations to avoid biases in the model selection process (Nakagawa and Freckleton 2008, 2011). The procedure uses bootstrapping and a model-based algorithm to provide data points for every missing data (Schafer 1999, Nakagawa and Freckleton 2011). We used the R function "amelia" (Honaker et al. 2011, R Development Core Team 2011) to produce 10 imputed data sets (as recommended by Rubin 1987). We thus performed all subsequent analyses included in the two analytical steps on the 10 imputed data sets.
First step: Variables selection
We used GLMMs to assess the effect of i- environmental conditions (population density, NDVI, NPI gestation, and NPI winter), ii- offspring characteristics (mass and sex), iii-maternal characteristics (age-specific mass, age-specific social rank, age, and iii-maternal care), iv- offspring development (frequency of locomotor play, frequency of play behaviour, and frequency of interaction), and v- relevant interactions on kid survival over the summer and over the winter (see App. A). The data set for summer survival (n=267 kids from 99 mothers) was slightly larger than for winter survival (n=209 kids from 90
28
mothers) because the latter excluded kids that died over the summer. We used the "glmer" function from the R package "lme4" (Bates et al. 2011) with a logit link, and included mother identity (ID) as a random intercept to account for the repeated measurements on mothers. Because the determination of the exact number of parameters is not clear in mixed models, and hence the degrees of freedom on which p-values are evaluated are approximated (Baayen et al. 2008), we wanted to avoid selecting the predictors using statistical significance methods. We thus determined whether a variable was an influent predictor of offspring survival using a model selection approach based on the Akaike's information criterion adapted for small samples (AICc; Burnham and Anderson 2002). We identified a list of models a priori for each dependent variable (43 models for summer survival and 41 for winter survival, see App. A), and computed for each model the average AAICc obtained from the 10 imputed data sets analysed. We considered all models with a AAICc of 4 or less as providing plausible support to the data (9 models for summer survival and 19 for winter survival; App. A; Burnham et al. 2011).
To present the estimate and the effect size of each variable included in the selected models, we performed model averaging. We avoided averaging the models using the raw estimates and the confidence intervals provided directly from the GLMMs because of the approximation problem of mixed models mentioned above. Instead, we performed Markov chain Monte Carlo (MCMC, 10 000 iterations) bootstraps set on posterior distribution of the parameters, and we calculated adjusted estimates and their 95% Bayesian credible intervals (CIs; see Baayen et al. 2008 for details). We used the function "pvals.fnc" in the "languageR" package to obtain the adjusted estimates and the CIs for linear mixed models (Baayen et al. 2008). Since this function is not yet implemented for GLMMs, we used a R script (obtained from Hamel et al. 2012) based on similar methods as the function "pvals.fnc", and that can calculate adjusted estimates and CIs for GLMMs. Because we had
10 imputed data sets, we had 10 estimates for each variable included in a model. We therefore averaged the estimates using the corresponding AICc weight for each model in a given data set. We calculated the median of the averaged estimates and the corresponding lower and upper boundaries of the CIs for all the variables included in the selected models. Finally, to determine the importance of each predictor, we computed the percentage of time
29
the CI of a variable excluded 0 out of all the estimates obtained for that variable (see Table 2).
Second step: Confirmatory multilevel path analysis
Once we had determined the most influential predictors, we assessed structural or causal relationships among them. Structural equation modeling (SEM) and path analysis are analogous statistical approaches to test these relationships, resembling the hypothético-déductive experimental method (Thomas et al. 2007). Because our models had hierarchical data structure (repeated measurements on mothers) and included a binary independent variable preventing multivariate normality, we were unable to perform SEM (Shipley 2000, 2009). We therefore used generalized multilevel path models, and followed Shipley's method of directional separation tests (d-sep tests; Shipley 2000, 2002). Path diagrams describe the causal relationships (arrows) between the variables (boxes) and the direction of these relationships (XI —» X2, or XI <— X2). Double-headed arrows represent correlation between two variables that cannot be explained by causation. Diagrams without feedback relationships are called directed acyclic graphs (DAGs; see Shipley 2002). The causal structure proposed by a DAG implies dependence and independence constraints between the variables (Shipley 2009). If the diagram describes adequately the causal process generating the data, the pattern of co-variations predicted by the DAG will then be mirrored in the data (non-significant Fisher's C test, see Shipley 2002). On the other hand, if the data show a significant lack of fit to the co-variations predicted (significantly different), the proposed DAG is rejected (Thomas et al. 2007).
DAGs - Because the results of the first selection of variables included several variables with a weak effect on survival, we only used variables and interactions with CI excluding zero for more than 10% of the models to prevent over-parameterization in the DAG models (see Table 2). However, we included "population density" in the DAGs for summer survival, which had only 4% of CIs excluding zero, because an interaction including this variable was selected (Table 2). For the same reason, we included "maternal mass" and
30
"maternal social rank" in the DAGs describing winter survival (0% and 5% of CIs excluding zero, respectively). Based on our knowledge of the population dynamics of goats at Caw Ridge (Côté and Festa-Bianchet 2001a, b, Festa-Bianchet and Côté 2008) and on the results obtained in our first analytical step, we constructed 9 causal models to describe the hypothesized relationships between the variables influencing summer survival (hypotheses SI to S9) and 19 for winter survival (hypotheses WI to W19). We only present the best DAG model (lowest C value with high p-value of the goodness-of-fit test indicates that the model provides a strong fit to the data, Shipley 2002) for each survival, but all DAGs tested are presented in Appendix B. Once the best causal model was found, we calculated the strength of each path as the coefficients estimated from the mixed models derived from the model (Shipley 2009). As described above, we obtained adjusted estimates and CIs for each path with MCMC bootstrapping methods. Because we obtained
10 estimates (i.e. from the 10 imputed data sets), we presented the median estimate and its corresponding CI. The selected causal models for summer and winter survival included a few variables with unclear relationships (path coefficients with 95% credible intervals including 0), but we kept these relationships because these models provided the best fit to the data (B. Shipley, pers. com.). To determine global effects, we calculated the estimates provided from simple GLMMs between each variable and survival (Shipley 2002).