Pépite | Les conséquences génétiques de la colonisation d’un milieu pollué par las métaux, des approches de génétique des populations et génétique quantitative
198
0
0
Texte intégral
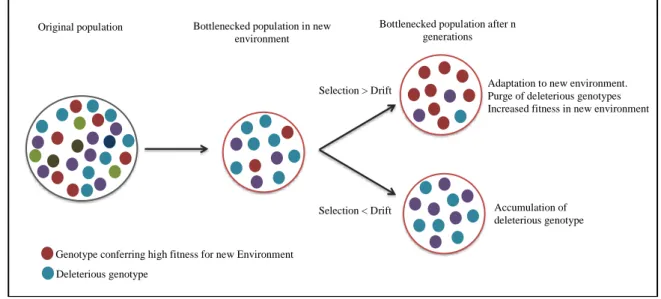
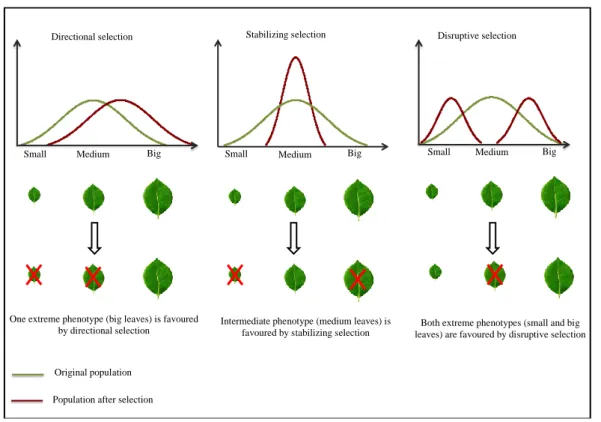
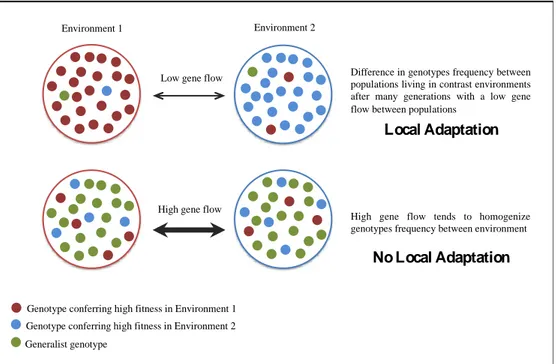
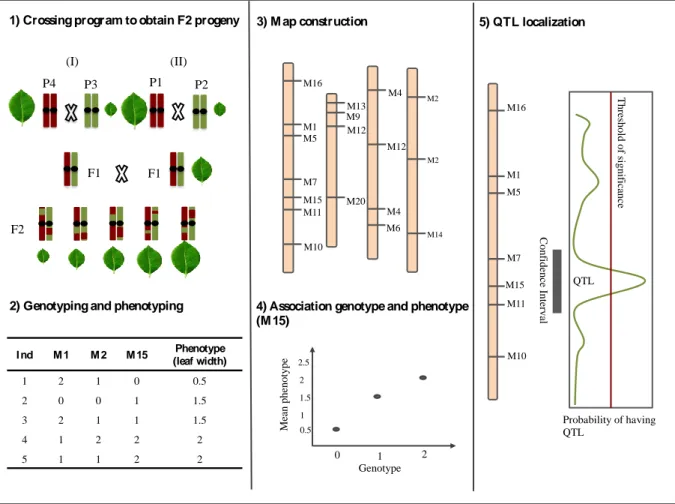
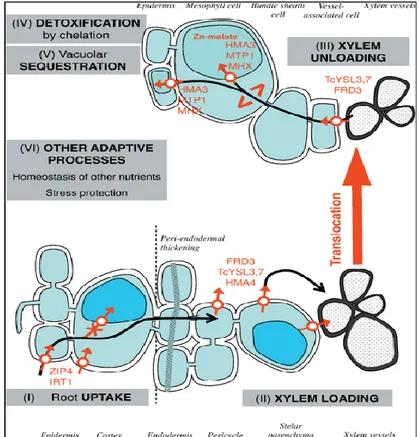
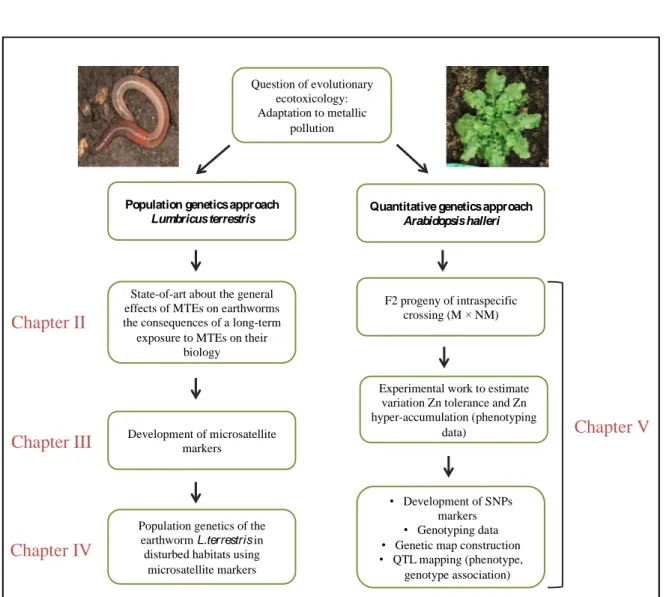
Figure
+7




Documents relatifs
