Régulation de l’épissage alternatif de l’exon 10 de tau
par la température
Thèse
Franck Petry
Doctorat en neurobiologie
Philosophiae doctor (Ph. D.)
Québec, Canada
© Franck Petry, 2017
Régulation de l’épissage alternatif de l’exon 10 de tau
par la température
Thèse
Franck Petry
Sous la direction de :
Résumé
La protéine tau est une protéine neuronale associée aux microtubules. L’exon 10 code pour un domaine de liaison aux microtubules et son épissage alternatif définit deux types d’isoformes ayant une fonction biologique distincte. En effet, quand l’exon 10 est exclu, les isoformes de tau ont trois domaines de liaison aux microtubules (Tau3R) alors que les isoformes en possèdent quatre lorsque l’exon 10 est inclus (Tau4R). Ainsi, les isoformes Tau4R sont connues pour avoir une meilleure affinité pour les microtubules, et permettent de mieux les stabiliser au sein de l’axone des neurones. Les tauopathies sont des maladies neurodégénératives qui se caractérisent par la présence d’agrégats de la protéine tau sous forme hyperphosphorylée. Parmi ces agrégats, certains sont composés des isoformes Tau3R et Tau4R, alors que d’autres sont essentiellement composés soit des isoformes Tau3R, soit des isoformes Tau4R. Ces données montrent qu’un défaut d’épissage alternatif de l’exon 10 de tau peut conduire à une pathologie. Cependant, la régulation de l’épissage alternatif de l’exon 10 est encore mal connue à la fois dans un contexte physiologique et pathologique et l’absence de données sur la physio-pathologie des tauopathies et de modèles d’études intégrés rendent l’avancement des connaissances et le développement de stratégies thérapeutiques nébuleux. De manière intéressante, il existe un changement d’épissage alternatif de l’exon 10 au cours du développement du cerveau chez la souris. En effet, les isoformes Tau3R sont majoritaires dans les premiers stades du développement et ne sont plus du tout exprimées à l’âge adulte. En revanche, le cerveau humain à l’âge adulte exprime autant d’isoformes Tau3R que d’isoformes Tau4R.
Nous avons remarqué que deux évènements ont lieu simultanément au cours du développement du cerveau chez la souris : le changement d’expression des isoformes de tau et la mise en place de la thermogénèse. Plusieurs études ont montré que la température influence le niveau de phosphorylation de la protéine tau, mais aucune n’a mis en évidence l’impact de la température sur l’épissage alternatif de tau. Ainsi, notre hypothèse de départ est que la température représente un nouveau régulateur de l’expression des isoformes de tau, en modulant l’épissage alternatif de l’exon 10. Les principaux objectifs de cette thèse étaient d’analyser l’impact de la température sur l’épissage alternatif de l’exon 10 de tau et les mécanismes cellulaires associés aux changements d’épissage alternatif de l’exon 10.
Dans un premier temps, nous avons utilisé le développement du cerveau chez la souris comme modèle pour faire une caractérisation plus précise de l’expression des isoformes de tau. Ensuite, nous avons utilisé des cultures de neurones primaires de souris, dans le but d’évaluer l’impact de
changements directs de température sur l’expression des isoformes de tau. Dans un deuxième temps, nous avons utilisé une approche in vitro, pour caractériser les changements d’épissage alternatif de l’exon 10 de tau au niveau de l’ARNm et protéique. Dans un troisième temps, nous avons analysé les mécanismes cellulaires responsables de ces changements d’expression.
Nos résultats montrent que la température affecte directement l’épissage alternatif de l’exon 10 au niveau de l’ARNm mais également des protéines synthétisées. De plus, nous avons vu que l’hypothermie favorise l’exclusion de l’exon 10, ce qui conforte notre observation de départ en lien avec le développement du cerveau chez la souris, tandis que l’hyperthermie favorise l’inclusion de l’exon 10 dans tous les modèles analysés. Nous avons également montré que la température affecte l’épissage alternatif de l’exon 10 humain. Nos résultats montrent également que la température affecte le patron développemental d’expression des isoformes de tau. De plus, nos résultats ciblent le facteur d’épissage Muscle blind-like (MBNL), comme mécanisme cellulaire responsable des changements d’expression des isoformes de tau induits par la température.
De manière préliminaire, nos travaux de recherche montrent ainsi un nouveau rôle de la température dans la régulation de l’expression des isoformes de tau. Les prochaines étapes seraient d’évaluer l’impact fonctionnel de ces changements d’expression de tau sur le cerveau et de tester les changements de température comme nouvelle avenue thérapeutique pour le traitement des tauopathies présentant un défaut d’épissage alternatif de l’exon 10 de tau.
Abstract
Tauopathies are a group of neurodegenerative disorders characterized by the presence of aggregates of hyperphosphorylated tau protein. These aggregates are either constituted of the six tau isoforms, or Tau4R isoforms or Tau3R isoforms, suggesting that altered tau exon 10 alternative splicing can lead to neurodegeneration. This was further supported by the discovery of mutations on matp gene mainly responsible for fronto-temporal dementia. The regulation of tau exon 10 alternative splicing is not fully understood in both physiological and pathological conditions. Indeed, the lack of data on the development of sporadic tauopathies (in absence of tau mutations) and models to study them make therapeutics strategies compromised.
Interestingly, changes of tau isoforms expression has been reported during the mouse brain development. Indeed, Tau3R isoforms are dominant in the first developmental stages (embryonic and early post-natal) and are absent in adulthood. To the opposite, human adul brain expresses both Tau3R and Tau4R to equal amount. This inter-species fundamental difference of expression of tau isoforms is not understood.
We noctided that some events are concomitant during the mouse brain development: shift of tau isoforms, shift of tau phosphorylation state and pups thermoregulation efficiency. It has been reported that temperature can influence the phosphorylation of tau protein and especially that hypothermia increases it. To date, no study has shown the impact of temperature on tau exon 10 alternative splicing. Thus, our hypothesis is that temperature is a new regulator of the expression of tau isoforms by modulating the alternative splicing of tau exon 10. The major goals of this thesis were to analyze the impact of temperature on the alternative splicing of tau exon 10 overall, and especially during the mouse brain development. On another hand, we want to analyze the mechanisms responsible for temperature-mediated tau exon 10 alternative splicing changes.
First, we used the mouse brain development to characterize the expression of tau isoforms. Thus, we used mouse primary neuronal cells in the aim of analyzing the impact of direct changes of temperature on tau isoforms expression. Second, we used in vitro approaches to characterize the impact of temperature on tau exon 10 alternative splicing at both mRNA and proteins levels. Third, we have analyzed mechanisms involved in these changes.
Our results show that temperature directly affects tau exon 10 alternative splicing at both mRNA and protein levels. For the first time, we show that hypothermia induces tau exon 10 exclusion
whereas hyperthermia favors exon 10 exclusion. Moreover, we show that temperature is able to modulate the alternative splicing of human tau exon 10. Our results with primary neuronal cells show that temperature can influence the developmental pattern of expression of tau isoforms, suggesting that temperature is a strong modulator of tau exon 10 alternative splicing. Eventually, our results highlight the role of Muscle blind-like proteins (MBNL) as potential mechanisms involved in the regulation of tau exon 10 alternative splicing by the temperature.
Interestingly, our work put in relief a new role of the temperature in the regulation of tau isoforms expression. As perspectives, it should be important to evaluate the functional impact of temperature-mediated changes of tau isoforms expression on brain.
Table des matières
RESUME ... III
ABSTRACT ... V
TABLE DES MATIERES ... VII
LISTE DES TABLEAUX ... X
LISTE DES FIGURES ... XI
LISTE DES ABREVIATIONS ... XII
REMERCIEMENTS ... XV
AVANT-‐PROPOS ... XVII
CHAPITRE I ... 1
INTRODUCTION ... 1
1 : LA PROTEINE TAU, EXPRESSION ET FONCTIONS ... 2
I. LE GENE MAPT ... 2
I. A. Localisation chromosomique et structure du gène mapt ... 2
I. B. Le promoteur ... 3
I. C. Le polymorphisme ... 5
II. L’EPISSAGE ALTERNATIF ... 5
II. A. Généralités sur l’épissage et l’épissage alternatif ... 5
II. A. 1. Généralités ... 5
II. A. 2. Les différents types d’épissage alternatif ... 7
II. B. Epissage alternatif de tau ... 8
II. B. 1. Epissage alternatif dans le système nerveux central ... 9
II. B. 2. Epissage alternatif dans le système nerveux périphérique ... 10
II. B. 3. L’exon 6 ... 11
III. LA PROTEINE TAU ... 12
III. A. Structure ... 13
III. A. 1. Le domaine de projection ... 14
III. A. 2. Le domaine de liaison aux microtubules ... 14
III. B. Modifications post-‐traductionnelles ... 14
III. B. 1. Phosphorylation ... 15
III. B. 2. Autres modifications post-‐traductionnelles ... 15
III. C. Fonctions de la protéine tau ... 17
III. C. 1. Liaison de tau aux microtubules ... 17
III. C. 2. Polarité cellulaire ... 19
III. C. 3. Transport synaptique ... 20
III. C. 4. Liaison à la membrane cytoplasmique ... 20
III. C. 5. Tau dans le noyau ... 21
III. D. Régulation des fonctions de la protéine tau ... 22
III. D. 2. Régulation par épissage alternatif ... 23
IV. TAU PENDANT LE DÉVELOPPEMENT ... 24
IV. A. Expression spécifique des isoforms de tau ... 24
IV. B. Hyperphosphorylation transitoire de la protéine tau ... 26
2 : MÉCANISMES CELLULAIRES IMPLIQUÉS DANS LA RÉGULATION DE L’EPISSAGE ALTERNATIF DE L’EXON 10 DE TAU ... 27
I. L’EPISSAGE ALTERNATIF ... 27
I. A. Mécanisme détaillé de l’épissage ... 27
II. REGULATION EN CIS ... 30
II. A. Les séquences régulatrices exoniques ... 31
II. B. Les séquences régulatrices introniques ... 33
II. C. Le motif tige-‐boucle ... 34
III. REGULATION EN TRANS ... 36
III. A. Les facteurs d’épissage ... 36
III. A. 1. Les protéines SR ... 36
III. A. 2. Les protéines hnRNPs ... 39
III. A. 3. Les autres facteurs d’épissage ... 40
III. B. Régulation des facteurs d’épissage : phosphorylation et localisation intracellulaire ... 42
3 : DE LA PROTEINE TAU A LA PATHOLOGIE ... 45
I. LES TAUOPATHIES : AGREGATION PATHOLOGIQUE DE TAU ... 45
I. A. Généralités ... 45
I. B. Composition biochimique des agrégats de tau ... 46
I. C. Distribution topographique et laminaire des agrégats de Tau ... 47
II. DEFAUT DE PHOSPHORYLATION: EXEMPLE DE LA MALADIE D’ALZHEIMER ... 50
II. A. Généralités ... 50
II. B. Lésions neuro-‐histo-‐pathologiques ... 51
II. B. 1. Pathologie amyloïde ... 52
II. B. 2. Pathologie Tau ... 54
III. DEFAUTS FONCTIONNELS ET D’EPISSAGE ALTERNATIF DE L’EXON 10 DE TAU ... 57
III. A. Dérégulation en cis ... 58
III. A. 1. Mutations sur le gène mapt ... 58
III. A. 1. 1. Liaison de tau aux microtubules ... 59
III. A. 1. 2. Epissage alternatif de l’exon 10 ... 62
III. A. 2. Haplotype ... 65
III. B. Tauopathies sporadiques ... 66
HYPOTHESES ET OBJECTIFS ... 68
CHAPITRE II ... 71
TEMPERATURE REGULATES TAU ISOFORMS EXPRESSION BY MODULATING TAU EXON 10 ALTERNATIVE SPLICING ... 71
CHAPITRE III ... 109
REGULATION DE L’EPISSAGE ALTERNATIF DE L’EXON 10 ... 125
CONCLUSION GENERALE ... 128
REFERENCES ... 129
Liste des tableaux
Tableau 1. Résumé des études qui ont analysé l’épissage alternatif de l’exon 10 de tau au cours du
développement chez l’humain et les rongeurs ... 25
Tableau 2. Classification des séquences cis régulatrices en fonction de la localisation sur le pré-ARNm (intron ou exon) et de l’effet sur l’épissage alternatif. ... 29
Tableau 3. Propriétés des protéines SR et impact sur l’épissage alternatif de l’exon 10 de tau. ... 153
Tableau 4. Distribution topographique et laminaire, composition des isoformes de tau dans les inclusions ainsi que leur profil électrophorétique pour les tauopathies majeures ... 49
Tableau 5. Principales mutations de tau qui affectent la liaison aux microtubules ... 60
Tableau 6. Principales mutations de tau qui affectent l’épissage alternatif de l’exon 10 ... 62
Tableau 7. List of antibodies used ... 106
Tableau 8. List of primers used ... 107
Tableau 9. Comparison of tau isoforms shit between development and primary neurons ... 108
Tableau 10. Correction des mutations affectant l’épissage alternatif de l’exon 10 ... 123
Liste des figures
Figure 1. Expression de la protéine tau dans le SNC : gène, ARN et protéines ... 3
Figure 2. Promoteur de l’expression du gène mapt, îlots CpG et polymorphisme ... 4
Figure 3. Schéma représentant l’épissage d’un pré-ARNm en ARNm ... 6
Figure 4. Schéma représentant l’épissage alternatif ... 7
Figure 5. Différents types d’épissages alternatifs ... 9
Figure 6. Expression de la protéine tau dans le SNP ... 11
Figure 7. Expression des isoformes de tau selon l’épissage alternatif de l’exon 6 ... 12
Figure 8. Représentation schématique structurelle de la protéine tau ... 13
Figure 9. Sites putatifs de phosphorylation de la protéine tau ... 16
Figure 10. Schéma représentant la formation des microtubules ... 18
Figure 11. Fixation de la protéine tau au niveau des microtubules ... 19
Figure 12. Transport axonal antérograde et rétrograde ... 21
Figure 13. La phosphorylation de tau diminue la fonction de liaison aux microtubules ... 23
Figure 14. Mécanisme général d’épissage ... 29
Figure 15. Séquences cis régulatrices de l’exon et de l’intron 10 de tau ... 31
Figure 16. Interaction entre les séquences cis régulatrices pour l’épissage de l’exon 10 ... 34
Figure 17. Motif tige-boucle au site d’épissage 5’ de l’exon 10 ... 35
Figure 18. Interaction des facteurs trans pour l’épissage de l’exon 10 ... 42
Figure 19. Code barre des tauopathies ... 47
Figure 20. Organisation laminaire du cortex cérébral ... 48
Figure 21. Photographies de l’équipe du Dr. Alois Alzheimer et d’Auguste Deter ... 50
Figure 22. Cerveaux d’individu sain et atteint de la maladie d’Alzheimer ... 51
Figure 23. Voies de clivage de la protéine précurseur de l’amyloïde (APP) ... 52
Figure 24. Marquage des plaques amyloïdes dans la maladie d’Alzheimer ... 53
Figure 25. Évolution de la pathologie amyloïde au cours de la maladie d’Alzheimer ... 54
Figure 26. Enchevêtrements neurofibrillaires dans la maladie d’Alzheimer ... 55
Figure 27. Évolution de la pathologie tau au cours de la maladie d’Alzheimer ... 56
Figure 28. Mutations sur le gène mapt ... 59
Figure 29. Types d’agrégats de la protéine tau ... 67
Figure 30. Température des nouveaux-nés au cours du développement post-natal chez le rat. ... 69
Figure 31. Analysis of tau exon 10 alternative splicing during mouse brain development. ... 94
Figure 32. Temperature modifies tau exon 10 alternative splicing and MBNL expression during the differentiation of mouse primary neurons. ... 96
Figure 33. Impact of temperature on tau exon 10 alternative splicing in N2a cells. ... 97
Figure 34. Temperature directly affects tau exon 10 alternative splicing at mRNA and protein levels in N2a cells. ... 100
Figure 35. Hypothermia induces re-expression of Tau3R isoforms. ... 102
Figure 36. Temperature affects the alternative splicing of human tau exon 10. ... 104
Figure 37. Complex regulation of microtubules dynamic by the temperature. ... 105
Figure 38. Correction de l’épissage alternatif par oligonucléotides antisens ... 118
Figure 39. Correction de l’épissage alternatif par trans-splicing ... 119
Liste des abréviations
aa Acide aminé
Aβ Peptide amyloïde β
Aβ40 Peptide amyloïde β 40
Aβ42 Peptide amyloïde β 42
ACE A/C-rich enhancer
ADN/DNA Acide désoxyribonucléique/desoxyribonucleic acid AICD Amyloid precursor protein intracellular domain
APP Amyloïde precursor protein
ARN/RNA Acide ribonucléique/ribonucleic acid
ARNm ARN messager
ASF Alternative splicing factor 1
ASO Anti-sens oligonucléotide
AVC Accident vasculaire cérébral
BHE Barrière hémato-encéphalique
CA1 Cornus ammonis 1
CDK Cyclin-dependent kinase
cDNA Complementary DNA
CaMKII Calcium/calmodulin-dependent protein kinase
CLK Clk/Sty protein kinase
CTX Cortex
DCB/CBD Dégénrérescence corticobasale/corticobasal degeneration
DFTP-17/FTDP-17 Démence fronto-temporale avec parkinsonisme liée au chromosome 17/fronto-temporal dementia with parkinsonisme linked to chromosome 17
DIV Days in vitro
dmpk Dystrophia myotonica protein kinase
DNF Dégénérescence neurofibrillaire
dscam Down syndrome cell adhesion molecule
Dyrk Dual specificity tyrosine-phosphorylation-regulated kinase
E2/E3 Exon 2/Exon 3
E10 Exon 10
E18 Embryonic 18 days
ECL Enhanced chemiluminescence
ESE Exonic splicing enhancer
ESM Exonic splicing modulator
ESS Exonic splicing silencer
GAPDH Glyceraldehyde 3’-phosphate dehydrogenase
GDP Guanosine-5’-diphosphate
GSK3 Glycogen synthase kinase 3
GTP Guanosine-5’-triphosphate
H1 Haplotype 1
H2 Haplotype 2
HC Hippocampus
HEK293 Human embryonic kidney 293
hnRNP Heterogeneous nuclear ribonucleoprotein particle
ISS Intronic splicing silencer
kb Kilo base
kDa Kilo dalton
MA Maladie d’Alzheimer
MAPK Mitogen-activated protein kinase
mapt Microtuble-associated protein tau
mb104 Monoclonal antibody 104
MBD Microtubule-binding domain
MBNL Muscleblind-like protein
MCI Mild cognitive impairment
MP Maladie de Parkinson
CELF CUG-BP- and ETR-3-like factor
MTs Microtubules N2a Neuro2a NFT Neurofibrillary tangles NMDA N-méthyl-D-aspartate nt Nucléotide P Post-natal pb Paire de bases
PBS Phosphate buffer saline
PCR Polymerase chain reaction
PHF Paired-helical filaments
PiD Pick’s disease
PKA Protéine kinase A
PP Protein phosphatase
PPE Polypurine ehancer
pré-ARNm Pré-ARN messager
protéine SR Protéine riches en sérines et arginines
PSF Polypyrimidine tract-binding protein-associated splicing factor PSP/PSP Paralysie supranucléaire progressive/progressive supranuclear palsy
PTM Pre-trans-splicing
R1/R2/R3/R4 Repeat 1/Repeat 2/Repeat 3/Repeat 4
RBM RNA binding motif protein
RIPA Radioimmunoprecipitation assay buffer
RRM RNA recognition domain
RS Domaine RS
RT-PCR Reverse transcription-PCR
sAPP Soluble amyloïd precursor protein
SC35 Splicing component 35 kDa
SD/DS Syndrome de Down/Down syndrome
SDS-PAGE Sodium dodecyl sulphatase-polyacrylamide gel electrophoresis
Ser Sérine
SF2 Splicing factor 2
SMaRT Spliceosome-mediated RNA trans-splicing
SNC Système nerveux central
SNP Système nerveux périphérique
SNPs Single-nucleotide polymorphisms
snRNA Small nuclear ribonucleic acid snRNP Small nuclear ribonucleoprotein SRSF Serine/arginine-rich splicing factor
Tau3R Tau three repeats
Tau4R Tau four repeats
Thr Thréonine
TMF Tau mouse forward
TMR Tau mouse reverse
TNRC Trinucleotide repeat containing protein
Tra2β Transformer protein 2β
TRF Tau rat forward
TRR Tau rat reverse
Tyr Tyrosine
UTR Untranslated region
VIH Virus de l’immunodéficience humaine
WB Western blot
WT Wild-type
Remerciements
Je voudrais remercier dans un premier temps tous les gens qui ont contribué à l’avancement de ma thèse et qui ont été présents tout au long de ces cinq années.
Merci aux membres du jury d’évaluer mon travail de thèse.
Un grand merci à mon directeur de thèse, le Dr. Emmanuel Planel, de m’avoir soutenu tout au long du doctorat (et même pendant une partie de ma maitrise). Je voudrais te remercier plus particulièrement pour ta présence, ton accueil à Québec et dans le laboratoire, les nombreuses conférences scientifiques aussi bien locales qu’internationales, les articles obtenus en collaboration et également pour ta confiance.
J’exprime également mes remerciements à tous les membres du laboratoire (tau team) et plus particulièrement à Maud Gratuze et à Françoise Morin pour leur soutien, leur aide technique mais également pour leur amitié. Cinq années de doctorat c’est long et on a le temps de voir passer des personnes dans le laboratoire : Alexis Bretteville, Carl Julien, Noura El-khouri, Maya Dickler, François Bezeau, Anastasia Noel, Geoffrey Truchetti, Jacynthe Julien, Isabelle Guisle, Andréanne Turgeon, Amy Bouchard, Laura Eyoum Jong, ainsi que tous les stagiaires et les étudiants. Je voudrais également remercier toutes les personnes que j’ai pu fréquenter tout au long de mon doctorat faisant partie des équipes adjacentes et avec qui j’ai pu développer des liens privilégiés, notamment Mathilde Henry, Nicolas Josset et Melody Mazon.
Je voudrais également remercier le Dr. Nicolas Sergeant, le Dr. Luc Buée et le Dr. Valérie Vingtdeux pour les deux stages de collaboration effectués au sein de leur laboratoire à Lille. À ce sujet, je remercie également l’Université de Lille 2 pour la bourse de mobilité internationale CABRI que j’ai obtenu lors de mon deuxième stage. Ces stages ont notamment permis de développer les cultures de neurones primaires en collaboration avec l’équipe du Dr. Valérie Vingtdeux. Je remercie donc tous les membres de son équipe pour leur aide, aussi bien technique que morale : Sébastien Didier, Claudia Marinangeli (mi piacconno le albicocche) et Manon Domise. Je voudrais également remercier tous les membres de l’équipe du Dr. Nicolas Sergeant : Sabiha Eddarkaoui pour son aide précieuse et son écoute, ainsi que Caroline, Hélène, Mathilde, Anthony, Alexandre et Floriant pour les bons moments passés dans et à l’extérieur du laboratoire. Je tiens aussi à remercier tous les
autres membres qui constituent l’équipe 1 du Dr. Luc Buée et qui ont rendu mon stage à Lille très agréable. Un merci tout spécial à mes deux amies de goûter Mégane Homa et Laura Morant et pour ces moments délicieux passés en votre compagnie.
Je souhaite remercier chaleuresement tous les membres de ma famille pour leur amour, leur encouragement et leur soutien tout au long de ma vie et notamment pendant ces cinq années de ma thèse. Je voudrais tout particulièrement remercier mes parents, mes grands-parents Papou et Mamou, Jessica c(p+m), Ludivine cp2 et bien sûr mon frère jumeau Renaud.
Je tiens également à remercier mes amies intimes: Coralie Kilburg (cocopower) et Isabelle Poitras pour tous les bons moments qu’on a passé ensemble, et qu’il nous reste à encore vivre.
Je remercie également ma nouvelle gérante Marie-Pier et tous les employés de Lush, de m’avoir accepté nouvellement au sein de leur équipe et pour tous les beaux moments passés avec vous.
Je voudrais également remercier mon idôle Serena Williams, meilleure joueuse de tennis au monde, notamment pour avoir été un modèle de persévérence, d’humilité et de combativité.
Finalement, je souhaite remercier tous les organismes subventionnaires, sans lesquels ce travail de doctorat n’aurait pas été possible : le CRSNG pour la subvention de recherche, la Société Alzheimer du Canada pour ma bourse de doctorat, l’Université de Lille 2 pour la bourse de mobilité internationale CABRI et le Centre Thématique de Recherche en Neurosciences pour une autre bourse de doctorat, ainsi que l’AFIRMAQ et le RQRV pour des bourses de voyage.
Avant-propos
Au cours de mon doctorat, j’ai eu la chance de pouvoir participer à des plusieurs projets qui ont fait l’objet d’une publication scientifique :
- Dexmedetomidine increases tau phosphorylation under normothermic conditions in vivo and in vitro, dans Neurobiology of Aging (Whittington, Virag et al. 2015)
- Hypothermia-induced hyperphosphorylation: a new model to study tau kinase inhibitors, dans Scientific Reports (Bretteville, Marcouiller et al. 2012)
- Deregulation of protein phosphatase 2A and hyperphosphorylation of tau protein following onset of diabetes in NOD mice, dans Diabetes (Papon, El Khoury et al. 2013)
- ERK (MAPK) does not phosphorylate tau under physiological conditions in vivo or in vitro, dans Neurobiology of Aging (Noel, Poitras et al. 2015)
- Hypothermia mediates age-dependent increase of tau phosphorylation in db/db mice, dans Neurobiology of Disease (El Khoury, Gratuze et al. 2016)
- Insulin deprivation induces PP2A inhibition and tau hyperphosphorylation in hTau mice, a model of Alzheimer’s disease-like tau pathology, dans Scientific Reports (Gratuze, Julien et al. 2017)
J’ai également eu la chance de participer à un projet de caractérisation des anticorps de tau, dont je suis le premier auteur et qui est publié dans PlosOne : specificity of anti-tau antibodies when analyzing mice models of Alzheimer's disease: problems and solutions (Petry, Pelletier et al. 2014). Dans cet article, j’ai participé au désign expérimental (avec l’aide de mon directeur de recherche Emmanuel Planel et d’un étudiant au post-doctorat Alexis Bretteville), aux sacrifices des souris, à la préparation des échantillons et à la majorité des expériences (avec l’aide de Jérôme Pelletier et de Françoise Morin). J’ai également réalisé les figures et écrit la première version de l’article, qui a été corrigée par Frédéric Calon, Sébastien Hébert et Robert Whittington.
Au cours de la dernière année de mon doctorat, j’ai également participé à un projet de collaboration entre les laboratoires du Dr. Emmanuel Planel et du Dr. Frédéric Bretzner. Ce projet a pour but d’analyser la phosphorylation de la protéine tau dans une souris modèle de pathologie tau après une lésion de la moëlle épinière. Les expériences et l’écriture de l’article sont présentement en cours. J’ai participé au design expérimental (avec l’aide d’Emmanuel Planel, de Frédéric Bretzner et d’un étudiant au doctorat dans l’équipe du Dr. Bretzner Nicolas Josset). Les lésions de la moëlle épinière chez les souris ont été realisées dans le laboratoire du Dr. Bretzner. Au sein de notre laboratoire, j’ai
pris en charge la préparation des échantillons. Les expériences ont été réalisées en grande partie par moi-même, avec l’aide de Françoise Morin, Maud Gratuze et Amy Bouchard. J’ai procédé aux analyses statistiques, à la réalisation des figures et à l’écriture de certaines sections de l’article, avec l’aide d’Amy Bouchard.
Enfin, j’ai également eu le privilège de travailler sur l’épissage alternatif de l’exon 10, qui est le sujet principal de mon doctorat. En effet, la thématique de recherche dominante du laboratoire du Dr. Emmanuel Planel est l’étude de la phosphorylation de la protéine tau. Ainsi, j’ai pu développer au sein du laboratoire une nouvelle thématique de recherche. Dans ce contexte, mes travaux de recherche m’ont permis d’écrire un article dont je suis le premier auteur en cours de préparation (non soumis), s’intitulant « Temperature regulates tau isoforms expression by modulating exon 10 alternative splicing » et faisant l’objet du chapitre 2. Dans cet article, nous montrons que l’épissage alternatif de l’exon 10 de tau répond à des variations de température dans de nombreux modèles in vitro et in vivo. De plus, une partie du désign expérimental et des expériences a été réalisée en collaboration avec le laboratoire du Dr. Nicolas Sergeant et du Dr. Valérie Vingtdeux (Lille. France). Ainsi, j’ai participé au désign expérimental, à la préparation des échantillons, à la majorité des expériences (cellules, cultures primaires et souris), aux analyses statistiques, aux interprétations, à la réalisation des figures et à l’écriture de l’article. La partie mini-gènes et facteurs MBNL de mon projet ont été développé dans le laboratoire du Dr. Nicolas Sergeant. La partie culture primaire a été faite dans le laboratoire du Dr. Valérie Vingtdeux. La partie culture cellulaire et in vivo a été réalisée dans le laboratoire du Dr. Emmanuel Planel.
Chapitre I
1 : LA PROTEINE TAU, EXPRESSION ET FONCTIONS
La protéine tau a été découverte pour la première fois en 1975 par Weingarten et ses collaborateurs, sous la forme d’un facteur essentiel à la polymérisation des dimères de tubuline (Weingarten, Lockwood et al. 1975). Depuis cette découverte, la protéine tau a été largement étudiée, puis a été associée à différentes pathologies neurodégénératives regroupées sous le nom de tauopathies (pour revue (Buee, Bussiere et al. 2000)).
Dans cette première partie de l’introduction, nous décrirons la protéine tau dans son ensemble, à commencer par son expression, sa régulation et ses fonctions principales.
I. LE GENE MAPT
I. A. Localisation chromosomique et structure du gène mapt
Comme toutes les protéines, la protéine tau suit un schéma d’expression traditionnel qui comprend la transcription du gène en pré-ARN messager (pré-ARNm), la maturation du pré-ARNm en ARN messager (ARNm) et la traduction de l’ARNm en protéine.
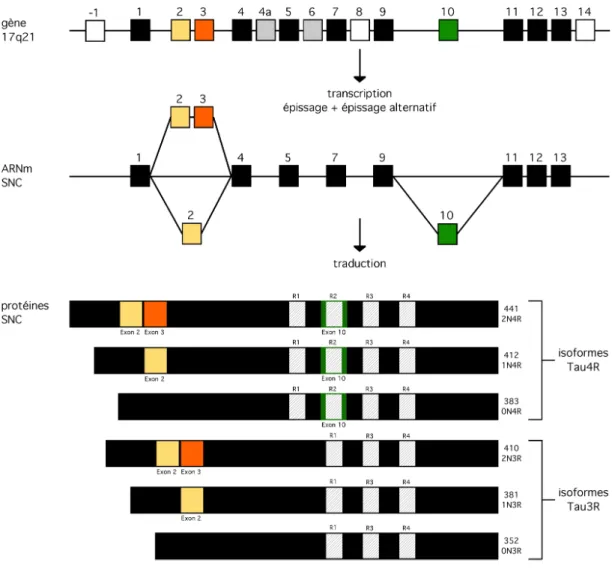
La protéine tau est encodée par un seul gène de 134 kb, le gène mapt (microtubule-associated protein tau), localisé sur le bras long du chromosome 17 en position 17q21.31 (Neve, Harris et al. 1986). Le gène mapt contient 16 exons, mais seulement 14 d’entre eux sont codants, donc seront utilisés pour la synthèse de la protéine tau (Figure 1). L’exon -1 ou 0 correspond à une partie du promoteur et de la région 5’ non traduite (5’UTR) (Andreadis, Brown et al. 1992) alors que l’exon 14 correspond à une partie de la région 3’ non traduite (3’UTR) (Goedert, Spillantini et al. 1989; Andreadis, Brown et al. 1992). Les exons 1, 4, 5, 7, 9, 11, 12 et 13 sont constitutifs alors que les exons 2, 3, 4a, 6, 8 et 10 subissent un épissage alternatif.
Le gène mapt peut coder pour trois transcrits différents de 2, 6 et 9 kb dépendamment de l’épissage alternatif de ces six exons non constitutifs, mais également du stade de différentiation neuronale ainsi que le type neuronal (Goedert, Spillantini et al. 1989; Goedert, Spillantini et al. 1989; Couchie, Mavilia et al. 1992; Goedert, Spillantini et al. 1992; Wang, Loomis et al. 1993; Nunez and Fischer 1997). En effet, le transcrit de 2 kb code pour la protéine tau nucléaire (Thurston, Pena et al. 1997), le transcrit de 6 kb pour les six isoformes axonales de tau trouvées dans le système nerveux central (SNC) et le transcrit de 9 kb permet l’expression d’une protéine tau spécifique de la rétine et des
Figure 1. Expression de la protéine tau dans le SNC : gène, ARN et protéines
Représentation schématique du gène mapt, de son ARNm et des isoformes de la protéine tau exprimées dans le SNC. Le gène mapt contient 16 exons. Les exons -1 (promoteur), 8 et 14 ne sont pas transcrits et donc ne sont pas présents dans le pré-ARNm (carrés blancs). Dans le SNC, les exons 4a et 6 ne sont pas inclus (carrés gris). Les exons 2, 3 et 10 sont alternatifs, conduisant à l’expression de six isoformes possibles (carrés jaune, orange et vert). Les autres exons sont constitutifs (carrés noirs). La taille en acides aminés et le symbole sont indiqués à côté de chaque isoforme.
I. B. Le promoteur
Le promoteur est une séquence cis régulatrice, le plus souvent retrouvé en amont du gène. Chez les eucaryotes, il est souvent constitué d’un motif TATA à une dizaine de nucléotides en amont du site de démarrage de la transcription. Le promoteur permet de réguler la fixation des facteurs de
transcription, qui à leur tour permettent d’appareiller la machinerie transcriptionnelle localement au niveau du gène à transcrire.
Le promoteur du gène mapt est mal défini et ne contient pas de séquences cis régulatrices eucaryotes classiques, tels que les motifs TATA ou CAAT. Cependant, d’autres motifs de réponse aux facteurs de transcription Sp1 et AP-2 ont été mis en évidence au niveau du promoteur du gène mapt, et seraient impliqués dans son expression spécifique au niveau des neurones (Andreadis, Wagner et al. 1996; Sadot, Heicklen-Klein et al. 1996; Heicklen-Klein and Ginzburg 2000). De plus, le promoteur du gène mapt contient des régions riches en nucléotides cytosine et guanine, appelées îlots CpG. Les îlots CpG subissent des modifications épigénétiques par méthylation, réprimant ainsi l’expression du gène. Une augmentation de la méthylation des îlots CpG du gène mapt a été décrite avec l’âge et serait responsable d’une diminution de l’expression de la protéine tau au cours du vieillissement (Tohgi, Utsugisawa et al. 1999). De plus, les îlots CpG300 et CpG27
situés dans la région du promoteur subissent des variations de méthylation dans différentes régions du cerveau, mais également entre hommes et femmes (Coupland, Mellick et al. 2014; Iwata, Nagata et al. 2014), pouvant ainsi expliquer les différences d’incidence de maladies neurodégénératives entre les genres (Figure 2).
Figure 2. Promoteur de l’expression du gène mapt, îlots CpG et polymorphisme
I. C. Le polymorphisme
Le polymorphisme correspond à des différences dans la séquence en nucléotides d’un gène donné dans une population donnée. En d’autres termes, il définit l’existence de plusieurs allèles pour un gène. Dans la population, il existe deux haplotypes majeurs pour le gène mapt: H1 et H2. L’haplotype H1 est plus représenté dans la population que l’haplotype H2, avec des variations en fonction de la population analysée (Evans, Fung et al. 2004). De plus, le niveau d’expression de la protéine tau peut varier en fonction de l’haplotype, avec une plus forte expression de tau pour les porteurs de l’haplotype H1/H1 comparé aux porteurs aux porteurs des haplotypes H1/H2 et H2/H2 (Valenca, Srivastava et al. 2016), notamment par un changement des niveaux de méthylation de l’ADN (Coupland, Mellick et al. 2014; Li, Chen et al. 2014). Enfin, l’haplotype du gène mapt a été associé à certaines tauopathies, notamment en intervenant au niveau de l’épissage alternatif de plusieurs exons. Nous détaillerons cette partie dans la troisième section de l’introduction.
II. L’EPISSAGE ALTERNATIF
II. A. Généralités sur l’épissage et l’épissage alternatif II. A. 1. Généralités
Après la première étape de transcription du gène en pré-ARNm, celui-ci subit une étape de maturation en ARNm consistant en l’ajout de la coiffe à l’extrémité 5’, la polyadénylation à l’extrémité 3’, l’épissage et l’épissage alternatif pour la plupart des pré-ARNm. L’épissage et l’épissage alternatif sont deux mécanismes cellulaires similaires d’un point de vue moléculaire et ont lieu en même temps au niveau du processus de maturation du pré-ARNm. Cependant, ils ont des rôles distincts.
L’épissage correspond à l’excision des séquences nucléotidiques qui ne codent pas pour la protéine (intron) et à la réunion des séquences nucléotidiques codantes (exon) restantes, dans le but de former un seul brin d’ARN. Cette étape nécessite la reconnaissance des jonctions entre l’intron et l’exon et implique des réactions enzymatiques consistant en une double trans-estérification (Figure 3). L’épissage est sous le contrôle d’un complexe macromoléculaire composé de petits ARN nucléaires et de nombreuses protéines, appelé splicéosome.
Figure 3. Schéma représentant l’épissage d’un pré-ARNm en ARNm
L’épissage est une étape moléculaire post-transcriptionnelle intervenant dans la maturation d’un pré-ARNm en ARNm. Celle-ci fait intervenir un complexe moléculaire appelé splicéosome et consiste en l’excision des introns non codants et à la réunion des exons codants pour ne former qu’un seul brin d’ARNm. Les exons sont représentés par des rectangles roses et les introns par des rectangles violets.
L’épissage alternatif est un mécanisme cellulaire par lequel certains exons ne vont pas être retenus de manière sélective dans l’ARNm. Ainsi, cette étape post-transcriptionnelle permet de générer plusieurs ARNm à partir du même transcrit, chacun ayant une combinaison différente en exons, qui à leur tour seront traduits en protéines différentes d’un point de vue biologie ou encore en isoformes d’une même protéine (Figure 4).
Chez l’humain, environ 95% des gènes sont régulés par épissage alternatif, ce qui permet d’augmenter la diversité biologique des protéines (Pan, Shai et al. 2008; Wang, Sandberg et al. 2008). L’exemple le plus flagrant concerne l’épissage alternatif du pré-ARNm codant pour la protéine Dscam («Down syndrome cell adhesion molecule» en anglais) chez Drosophilia melanogaster puisque ce processus peut produire jusqu’à 38016 ARNm (Schmucker, Clemens et al. 2000). Nous décrirons le mécanisme d’épissage alternatif en détails dans la deuxième section de l’introduction.
Figure 4. Schéma représentant l’épissage alternatif
L’épissage alternatif est une étape moléculaire post-transcriptionnelle intervenant dans la maturation du pré-ARNm en ARNm. Contrairement à l’épissage, l’épissage alternatif permet de former plusieurs ARNm ayant une combinaison en exons différente à partir du même pré-ARNm. Chacun des ARNm code pour une protéine biologiquement différente, avec une structure et une activité distincte. Adaptée du livre « THE CELL, Fourth Edition, Figure 5.5’ ».
II. A. 2. Les différents types d’épissage alternatif
Au niveau d’un pré-ARNm multi-exonique classique, différents types d’épissage alternatif peuvent être observés. Tout d’abord, les exons qui sont systématiquement inclus dans l’ARNm sont appelés constitutifs. Un exon qui est régulé par épissage alternative peut être inclus ou exclu de l’ARNm et donc codera ou non pour une partie de la protéine. Il s’agit donc d’un exon cassette, et constitue la majorité des exons régulés par épissage alternatif.
Cependant, le mécanisme d’épissage alternatif n’est pas aussi simple que d’inclure ou exclure un exon et différents épissages peuvent avoir lieu (Figure 5) :
- les exons mutuellement exclusifs : dans ce modèle deux exons sont en compétition pour être inclus dans l’ARNm. Ainsi, l’inclusion d’un exon entraine de manière systématique l’exclusion de l’autre exon. C’est le cas des exons 2 et 3 du pré-ARNm de l’alpha-tropomyosine (Smith and Nadal-Ginard 1989).
- les sites d’épissage alternatifs : dans ce modèle d’autres sites présents dans l’exon peuvent être utilisés comme site d’épissage au lieu du site traditionnel en 3’ ou 5’, entrainant ainsi la
perte ou non d’une partie de l’exon (Wang, Shi et al. 2006; Bielli, Bordi et al. 2014). Il s’agit le plus souvent d’un site ayant une activité plus forte que le site d’épissage en 3’ ou 5’ traditionnel.
- les promoteurs alternatifs : au lieu d’utiliser l’extrémité 5’ de l’exon comme site d’épissage, ce site servira de promoteur alternatif induisant ainsi l’exclusion de tous les exons en amont du nouveau promoteur alternatif. Un promoteur alternatif a été retrouvé pour le facteur d’échange GDP/GTP kalirin-7 et interviendrait dans son expression spécifique dans plusieurs tissus au cours du développement (Miller, Yan et al. 2016).
- la polyadénylation alternative : dans ce modèle un site d’épissage en 3’ peut être utilisé comme signal de maturation en 3’, donc de polyadénylation. Ainsi, les exons en aval du nouveau site de polyadénylation seront exclus de l’ARNm. Ce modèle d’épissage alternatif permet la synthèse de protéines ayant une région C-terminale de composition différente, mais également de protéines exprimées dans différents tissus (Tian and Manley 2016).
- la rétention d’introns : dans ce modèle une partie ou la totalité de l’intron peut être retenu au niveau de l’ARNm. Les rétentions d’introns peuvent amener à des anomalies le plus souvent via l’apparition d’un codon stop, d’un décalage du cadre de lecture ou de l’expression de protéines non fonctionnelles. Ce défaut d’épissage alternatif a été notamment associé au développement de cancers (Caballero, de Souza et al. 2001).
II. B. Epissage alternatif de tau
La protéine tau est fortement régulée au niveau de l’épissage alternatif, ce qui conduit à l’expression de plusieurs isoformes. Les isoformes de tau sont régulées de manière tissu-spécifique (Schoenfeld and Obar 1994), mais également au cours du développement et dans le cerveau adulte. Le rôle des différentes isoformes n’est pas encore connu, mais en revanche leur expression spatio-temporelle a été largement étudiée dans le système nerveux.
D’un point de vue anatomique, le système nerveux est divisé en deux parties chez les vertébrés: une partie centrale (SNC) comprenant le cerveau et la moelle épinière et une partie périphérique (SNP) comprenant les ganglions et les nerfs qui partent du cerveau et de la moelle épinière.
Figure 5. Différents types d’épissages alternatifs
L’épissage alternatif est un processus très complexe qui fait intervenir de nombreux mécanismes de régulation. Il existe différents types d’épissage alternatifs qui conduisent à l’inclusion ou à l’exclusion de l’exon. Les carrés gris correspondent aux exons constitutifs et les carrés rose et bleu aux exons alternatifs. Adaptée de « Zahler, A.M. Alternative splicing in C. elegans (September 26, 2005), WormBook, ed. The C. elegans Research Community, WormBook, doi/10.1895/ wormbook.1.31.1».
II. B. 1. Epissage alternatif dans le système nerveux central
Dans le système nerveux central, les exons 2, 3 et 10 sont régulés par épissage alternatif et conduisent à l’expression de six isoformes de la protéine tau (Goedert, Spillantini et al. 1989; Goedert, Spillantini et al. 1989; Andreadis, Brown et al. 1992) (Figure 1).
Les exons 4a et 8 sont systématiquement exclus de l’ARNm dans le SNC. L’exon 2 fonctionne comme un exon casette alors que l’exon 3 n’est pas inclus en absence de l’exon 2. Leur position N-terminal au niveau de la protéine permet d’identifier les isoforms de tau avec la nomenclature 0N (sans les exons 2 et 3 ou 2-3-), 1N (avec seulement l’exon 2 ou 2+3-) et 2N (avec les exons 2 et 3 ou 2+3+). Goedert et collaborateurs ont analysé leur niveau d’expression dans le cerveau humain adulte et ont montré que l’isoforme 1N est majoritaire (54%), suivie de l’isoforme 0N (37%) et de l’isoforme 2N en minorité (environ 9%) (Goedert and Jakes 1990). De plus, une distribution différentielle des isoformes 0N, 1N et 2N de tau a été mise en évidence dans plusieurs structures du cerveau et même d’un point de vue de leur localisation au sein du neurone chez la souris (Liu and Gotz 2013).
L’exon 10 est un exon cassette régulé par épissage alternatif et code pour un domaine de liaison aux microtubules (MTs). Trois autres domaines de liaison aux MTs sont présents dans les exons constitutifs 9, 11 et 12. Ainsi, l’épissage alternatif de l’exon 10 permet de réguler le nombre de domaines de liaison aux MTs et définit deux types d’isoformes. En effet, lorsque l’exon 10 est exclu, les isoformes de tau auront trois domaines de liaisons aux MTs et sont appelées Tau3R (« 3 repeats » en anglais). En revanche, les isoformes de tau possèdent quatre domaines de liaison aux MTs lorsque l’exon 10 est inclus et sont appelées Tau4R (« 4 repeats » en anglais). Les neurones du SNC expriment autant de Tau3R que de Tau4R à l’âge adulte chez l’humain (Goedert and Jakes 1990), mais ce ratio peut changer en fonction de la population de neurones.
En combinant les deux nomenclatures, les abréviations suivantes sont utilisées pour définir les six isoformes de la protéine tau : 0N3R (2-3-10-), 0N4R (2-3-10+), 1N3R (2+3-10-), 1N4R (2+3-10+), 2N3R (2+3+10+) et 2N4R (2+3+10+) (Figure 1).
II. B. 2. Epissage alternatif dans le système nerveux périphérique
Un transcrit de 9 kb codant pour une protéine tau ayant un poids moléculaire plus élevé d’environ 110 kDa a été retrouvé au niveau du système nerveux périphérique, et notamment dans les ganglions trigéminés et rachidiens (Couchie, Mavilia et al. 1992; Goedert, Spillantini et al. 1992; Nunez and Fischer 1997). Goedert et ses collaborateurs ont réussi à cloner la protéine tau à haut poids moléculaire de 758 acides aminés en 1992 et l’ont baptisée « big tau ». La big tau contient les mêmes exons constitutifs que ceux du système nerveux central, mais possède également les exons alternatifs 2, 3, 4a, 8 et 10 (Figure 6).
Figure 6. Expression de la protéine tau dans le SNP
Représentation schématique du gène mapt, de son ARNm et de l’isoforme de la protéine tau exprimée dans le SNP. Le gène mapt contient 16 exons. Les exons -1 (promoteur), 8 et 14 ne sont pas transcrits et donc ne sont pas présents dans le pré-ARNm (carrés blancs). Dans le SNP, tous les exons alternatifs deviennent constitutifs, conduisant à la synthèse d’une isoforme de tau longue de 758 acides aminés appelée « big tau ». En effet cette isoforme contient les exons 2, 3, 4a (spécifique du SNP) et 6.
La différence majeure entre les isoformes de tau dans le SNC et le SNP réside dans l’inclusion de l’exon 4a, qui code pour 251 acides aminés. Son expression a été retrouvée au niveau des neurones moteurs du ganglion rachidien dès les stades embryonnaires mais également au niveau des neurones de la rétine (Boyne, Tessler et al. 1995). Le rôle de cette protéine n’est pas encore totalement connu, mais elle aurait une implication dans la stabilisation et le diamètre des axones du SNP, qui sont plus longs et plus larges que ceux du SNC (Liu, Wang et al. 2015).
II. B. 3. L’exon 6
L’exon 6 de tau fonctionne comme un exon cassette et peut donc être totalement absent (isoforme 6-) ou présent (isoforme 6+ ou 6c) dans l’ARNm. Cependant, l’exon 6 de tau contient également deux sites alternatifs en 3’ conduisant à l’expression de deux isoformes supplémentaires. On distingue ainsi les isoformes 6p et 6d lorsque les sites alternatifs proximal et distal sont utilisés (Lapointe, Horowitz et al. 2009) (Figure 7).
Figure 7. Expression des isoformes de tau selon l’épissage alternatif de l’exon 6
L’exon 6 de tau est un cas particulier parce qu’il fonctionne comme un exon cassette mais possède également deux sites d’épissage 3’ alternatifs. Ainsi, l’utilisation ou non du site d’épissage conventionnel en 3’ (jonction intron 5/exon 6) permet l’expression d’une protéine tau possédant l’exon 6 (isoforme 6+) ou sans l’exon 6 (isoforme 6-). Cependant, deux autres sites d’épissage 3’ alternatifs peuvent être utilisés et conduisent à l’expression d’une protéine plus courte possédant seulement une partie de l’exon 6 (isoformes 6p et 6d).
Luo et ses collaborateurs ont trouvé que ces variants d’épissage permettent l’expression d’une protéine stable qui est tronquée au niveau de la partie C-terminale (Luo, Leski et al. 2004; Luo, Tse et al. 2004). Dans les tissus du SNC (cerveau adulte, cervelet, hippocampe et moelle épinière), il semble y avoir une plus grande proportion d’isoformes 6+ (2-3-6+ et 2+3-6+) que d’isoformes 6d, 6p et 6- (Wang, Tse et al. 2007). Dans les tissus du SNP (muscle squelettique et cœur), l’isoforme majoritaire est la 2+3-6+ mais d’autres isoformes sont également détectées en plus petite quantité. Une diminution de l’expression des isoformes 6+ et une augmentation des isoformes 6d ont été retrouvées dans les cerveaux de patients atteints de dystrophie myotonique (Leroy, Wang et al. 2006), montrant ainsi que l’épissage alternatif de l’exon 6 est important en condition physiologique.
III. LA PROTEINE TAU
Depuis sa découverte en 1975 par Weingarten comme étant une protéine essentielle à la polymérisation des MTs, la structure, les propriétés et les fonctions de la protéine tau ont été largement étudiées. De nos jours, elle se révèle comme une protéine soluble, stable à différentes températures et notamment à la dénaturation par la chaleur, mais aussi sans conformation
animales telles que le nématode (Goedert 1996; McDermott, Aamodt et al. 1996), les mammifères (Goedert, Spillantini et al. 1989; Riederer and Innocenti 1991; Nelson, Stefansson et al. 1996), les insectes (Cambiazo, Gonzalez et al. 1995) et les amphibiens (Yin, Chou et al. 1995).
III. A. Structure
Dans le SNC, l’épissage alternatif des exons 2, 3 et 10 conduisent à la synthèse de six isoformes de taille variable allant de 352 (2-3-10-) à 441 (2+3+10+) acides aminés chez l’humain et de 351 à 440 acide aminés chez la souris. En gel SDS-PAGE, leur poids moléculaire se situe entre 45 et 65 kDa (Buee, Bussiere et al. 2000), dépendamment de leur niveau de modifications post-traductionnelles. D’un point de vue structurel, la protéine tau est divisée en deux grands domaines : le domaine de projection en position N-terminale et le domaine de liaison aux microtubules en position C-terminale (Figure 8). Ce dernier est la partie de la protéine tau qui se fixe sur les dimères de tubuline au niveau des microtubules. Ainsi, l’autre partie de la protéine tau se projette hors des microtubules, d’où le nom de domaine de projection. Les principales fonctions associées à ces deux domaines seront détaillées plus loin dans cette partie de l’introduction.
Figure 8. Représentation schématique structurelle de la protéine tau
D’un point de vue structurel, la protéine tau est divisée en deux grands domaines : le domaine de projection en position N-terminale et le domaine de liaison aux microtubules en position C-terminale. Le domaine de projection comprend une région acidique et une région riche en prolines, il permet l’interaction de la protéine tau à la membrane, de réguler le diamètre des axones et de participer à différentes voies de signalisation. Le domaine de liaison aux MTs assure la fonction principale de tau : se lier aux MTs et réguler leur dynamique. Adaptée de (Buee, Bussiere et al. 2000).
III. A. 1. Le domaine de projection
Le domaine de projection est synthétisé à partir des exons 1 à 9. Sa longueur et sa charge isoélectrique dépendent de l’épissage alternatif des exons 2 et 3. En effet, ces deux exons alternatifs codent chacun pour 29 acides aminés et leur épissage entraine des variations de longueur de la protéine tau allant de 0 à 58 acides aminés. De plus, ces deux exons acidifient la partie N-terminale de la protéine en raison de leur point isoélectrique faible de 3.5 pour l’exon 2 et de 4 pour l’exon 3. Le domaine de projection contient également une région riche en acides aminés proline d’avantage basiques, présentant un point isoélectrique de 11.4 (Kolarova, Garcia-Sierra et al. 2012).
III. A. 2. Le domaine de liaison aux microtubules
Le domaine de liaison aux microtubules est synthétisé à partir des exons 9 à 13 et sa composition chimique est d’avantage basique (Kolarova, Garcia-Sierra et al. 2012). Sa longueur et sa fonction dépendent notamment de l’épissage alternatif de l’exon 10, codant pour 31 acides aminés qui incluent un domaine de liaison aux MTs. La présence ou l’absence de l’exon 10 définit le nombre de domaines de liaisons aux MTs (ou répétition ou R) de la protéine tau. Ainsi, son exclusion induit la synthèse des isoformes Tau3R possédant trois répétitions alors que son inclusion favorise la synthèse des isoformes Tau4R à quatre répétitions. Le domaine de liaison aux MTs d’une longueur variable de 31 à 32 acides aminés est constitué d’une séquence conservée de 18 acides aminés et d’une séquence inter-répétition de 13 à 14 acides aminés (Goedert, Spillantini et al. 1989; Himmler, Drechsel et al. 1989; Kosik, Orecchio et al. 1989; Zhou, Yu et al. 2008). Les quatre répétitions du domaine de liaison aux MTs sont apportées par quatre exons distincts : R1 par l’exon 9, le R2 par l’exon 10, le R3 par les exons 11 et 12 et le R4 par l’exon 12.
III. B. Modifications post-traductionnelles
La protéine tau subit beaucoup de modifications post-traductionnelles, telles que la glycation, la N- et O-glycosylation, la transglutamination, la formation de ponts disulfure et l’ubiquitination (pour revue (Buee, Bussiere et al. 2000)). Cependant, la modification post-traductionnelle majeure de la protéine tau réside dans sa phosphorylation. Dans les prochains paragraphes nous allons décrire brièvement les modifications post-traductionnelles de tau et leur implication fonctionnelle pour la protéine.
III. B. 1. Phosphorylation
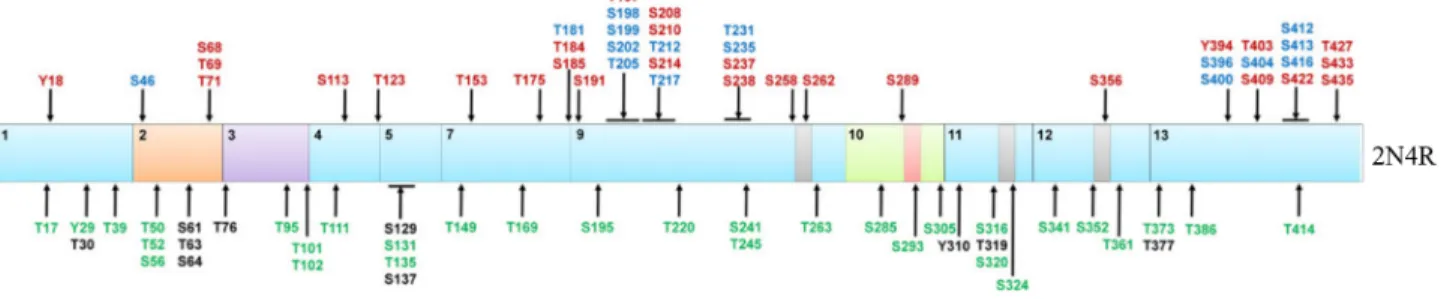
La protéine tau a souvent été décrite comme une protéine hautement phosphorylée ou phospho-protéine (Butler and Shelanski 1986). Au niveau de l’isoforme la plus longue du SNC (2N4R ou 2+3+10+), au moins 85 sites putatifs de phosphorylation ont été décrits et principalement au niveau des acides aminés sérine (45 Ser), thréonine (35 Thr) et tyrosine (5 Tyr) (Buee, Bussiere et al. 2000; Lebouvier, Scales et al. 2009) (Figure 9). Environ 20% des acides aminés de la protéine tau peuvent être phosphorylés, permettant de la désigner comme un « accepteur universel de groupements phosphates » (Stoothoff and Johnson 2005). La majorité des sites de phosphorylation sont retrouvées dans la partie C-terminale de la protéine tau, au niveau de la région riche en prolines et du domaine de liaison aux MTs (Sergeant, Bretteville et al. 2008; Hanger, Anderton et al. 2009). La plupart des sites de phosphorylation analysés sont des Ser/Thr suivies d’une proline, mais ces résidus peuvent également être phosphorylés en absence d’une proline. De nombreuses kinases et phosphatases sont impliquées dans la régulation de l’état de phosphorylation de la protéine tau en jouant sur son équilibre entre phosphorylation et déphosphorylation, respectivement. Les kinases qui ont été décrites pour phosphoryler tau au niveau des sérines et thréonines comptent la GSK-3 (« glycogen synthase kinase 3 » en anglais) (Hanger, Hughes et al. 1992), la famille des MAPK (« mitogen-activated protein kinase » en anglais) (Holzer, Gartner et al. 2001) et la famille des CDK (« cyclin dependent kinase » en anglais) incluant CDK2 et CDK5 (Baumann, Mandelkow et al. 1993; Hamdane, Sambo et al. 2003). Ensuite, des tyrosines kinases telles que les protéines Fyn et Src sont capables de phosphoryler tau au niveau de tyrosines (Lee, Thangavel et al. 2004; Derkinderen, Scales et al. 2005).
Les principales phosphatases exprimées au niveau du système nerveux central sont PP1, PP2A, PP2B et PP5. Toutes ces phosphatases sont capable de déphosphoryler la protéine tau (Tian and Wang 2002), mais PP2A semble avoir une meilleure affinité et être capable de déphosphoryler tous les épitopes de la protéine tau (Goedert, Jakes et al. 1995; Yu, Yoo et al. 2014).
III. B. 2. Autres modifications post-traductionnelles
La O-glycosylation est un processus post-traductionnel dynamique qui se caractérise par l’ajout d’un glucide (N-acetylglucosamine ou O-GlcNAc) au niveau d’un oxygène de la chaîne latérale d’une sérine ou d’une thréonine à proximité d’une proline (Haltiwanger, Kelly et al. 1992). La O-glycosylation affecte notamment les protéines associées aux MTs telles que MAP2 (« microtubule-associated protein 2 » en anglais) (Dong, Xu et al. 1993) et tau (Ding and Vandre 1996), mais en
plus faible proportion que la phosphorylation (Arnold, Johnson et al. 1996). Ces deux modifications ont un effet opposé sur la fonction de la protéine tau et la O-glycosylation intervient également dans la localisation cellulaire et la dégradation de la protéine tau (Arnold, Johnson et al. 1996). De plus, la O-glycosylation de tau est augmentée dans la maladie d’Alzheimer et interviendrait dans la formation et la stabilisation des agrégats de tau (Griffith and Schmitz 1995).
Figure 9. Sites putatifs de phosphorylation de la protéine tau
La plus longue isoforme de la protéine tau (2N4R) contient 85 sites putatifs de phosphorylation au niveau de sérines (45), de thréonines (35) et de tyrosines (5). Tous ces sites ne sont pas phosphorylés en même temps et en condition physiologique. Les sites de phosphorylation retrouvés dans un cerveau normal sont indiqués en vert, ceux retrouvés dans un cerveau normal et de patients atteints de la maladie d’Alzheimer sont indiqués en rouge et enfin ceux retrouvés seulement dans les cerveaux de patients atteints de la maladie d’Alzheimer sont indiqués en bleu. Certains sites n’ont jamais été décrits (en noir). Adaptée de (Martin, Latypova et al. 2011).
L’ubiquitination est une marque de stress impliquée dans la dégradation des protéines normales, anormales ou endommagées via le protéasome (Hershko and Ciechanover 1998). Plusieurs résidus au niveau du domaine de liaison aux MTs sont capables de recevoir une ubiquitination, telles que les lysines 254, 257, 311 et 317. Dans la maladie d’Alzheimer, l’ubiquitination de la protéine tau a été fortement détectée et notamment au sein des agrégats de tau (Mori, Kondo et al. 1987; Morishima-Kawashima, Hasegawa et al. 1993).
La glycation est une réaction entre la partie amine de la chaine latérale d’un acide aminé et la partie carboxyle d’un sucre réducteur, tel que le glucose. Treize résidus lysine ont été identifiés sur la plus longue isoforme de la protéine tau du SNC (2N4R) comme sites de glycation (Ledesma, Bonay et
anticorps reconnaissant un peptide glyqué et comprend une région de la protéine impliquée dans la liaison de tau aux MTs. Ainsi, une glycation de ses résidus pourrait diminuer la liaison de tau aux MTs (Ledesma, Bonay et al. 1995). Enfin, la glycation de tau induit un stress oxydatif, participant à la neurodégénérescence (Yan, Yan et al. 1995).
Une des premières étapes essentielles à la formation des agrégats de tau serait la formation de dimères reliés par ponts disulfures (Wille, Drewes et al. 1992). Les cystéines 291 et 322 des répétitions du domaine de liaison aux MTs R2 et R3 peuvent former des ponts disulfures de manière intramoléculaire (conformation en mâchoire), inhibant ainsi la formation de dimères entre les isoformes Tau4R, seules porteuses du R2 (Schweers, Mandelkow et al. 1995). Cependant, d’autres ponts disulfures intermoléculaires peuvent se former via la cystéine 322 et donc interveniennent dans l’agrégation de la protéine tau, mettant en relief l’importance du potentiel d’oxydo-réduction au sein des neurones.
La transglutaminase est une enzyme catalysant la formation de ponts entre des résidus glutamine. De la tau recombinante traitée avec de la transglutaminase est capable de former des structures filamenteuses in vitro (Appelt and Balin 1997), suggérant ainsi l’implication de cette enzyme dans la formation des agrégats de tau (Miller and Johnson 1995; Norlund, Lee et al. 1999).
III. C. Fonctions de la protéine tau
La protéine tau est une protéine principalement axonale, et sa fonction majeure est de participer à la polymérisation et la stabilité des microtubules dans l’axone (Cleveland, Hwo et al. 1977; Cleveland, Hwo et al. 1977; Avila, Lim et al. 2002). Cependant, la protéine tau a également été retrouvée au niveau de la membrane plasmique, des dendrites (synapse) et du noyau suggérant d’autres rôles au sein du neurone. De plus, une distribution différentielle des isoformes de tau dans plusieurs compartiments neuronaux a été rapportée, suggérant également un rôle propre de chaque isoforme et donc de l’épissage alternatif (Liu and Gotz 2013).
III. C. 1. Liaison de tau aux microtubules
Les microtubules sont des structures dynamiques majoritairement présentes au niveau des axones du neurone, et sont plus faiblement représentées dans les dendrites. Les MTs sont des composants du cytosquelette, et interviennent notamment dans la forme cellulaire. Au sein des axones des neurones, les microtubules forment un réseau du corps cellulaire vers la synapse qui est essentiel au transport axonal des neurotransmetteurs, des enzymes, des constituants de la synapse et d’autres
protéines. D’un point de vue moléculaire, les microtubules sont constitués de 13 protofilaments associés entre eux en forme de cylindre d’un diamètre de 24 nm (Figure 10) (Burns and Surridge 1994).
Figure 10. Schéma représentant la formation des microtubules
Les microtubules sont des composants du cytosquelette et sont présents dans les axones des neurones, où ils participent notamment au transport synaptique. Les MTs sont des structures cylindriques creuses de 24 nm constituées de treize protofilaments, eux même constitués d’hétérodimères de tubuline α et β. Les microtubules sont orientés et définissent une extrémité (+) permettant la polymérisation des MTs (élongation) et une extrémité (-) servant de site de dépolymérisation. Adaptée de (Jordan and Wilson 2004).
Les protofilaments des MTs sont eux même constitués d’hétérodimères de tubuline α chargée négativement et de tubuline β chargée positivement s’associant les uns les autres par des liaisons non covalentes en longs filaments. Les dimères de tubuline sont polarisés au niveau du protofilament formant ainsi une extrémité positive (+) servant de site de polymérisation et une extrémité négative (-) servant de site de dépolymérisation. Dans un état d’équilibre, le taux de polymérisation est égal au taux de dépolymérisation, on parle alors de tapis roulant. Dépendamment des facteurs présents et des conditions cellulaires, l’instabilité dynamique des MTs peut pencher vers un effondrement du réseau de MTs ou vers son expansion.
capable de se fixer au niveau des dimères de tubuline et stabilise les extrémités des MTs. La liaison de tau aux MTs se fait grâce à la présence de trois ou quatre répétitions (R1, R2, R3 et R4) du domaine de liaison aux MTs, codés par les exon 9 à 12 (Lee, Neve et al. 1989). De part cette liaison, la protéine tau est capable d’induire la polymérisation des MTs (Weingarten, Lockwood et al. 1975; Cleveland, Hwo et al. 1977; Cleveland, Hwo et al. 1977; Brandt and Lee 1993; Brandt and Lee 1993). De plus, la protéine tau est capable d’augmenter la vitesse de polymérisation à l’extrémité (+) et d’inhiber la dépolymérisation à l’extrémité (-) (Drechsel, Hyman et al. 1992). Au sein de chaque répétition, une séquence conservée de 18 acides aminés se lie aux MTs et présente une certaine flexibilité permettant la liaison des trois ou quatres répétitions sur plusieurs dimères de tubuline (Butner and Kirschner 1991) (Figure 11).
Figure 11. Fixation de la protéine tau au niveau des microtubules
La protéine tau est capable de se lier aux hétérodimères de tubuline grâce à la présence de répétitions du domaine de liaison aux MTs. Dépendamment de l’épissage alternatif de l’exon 10, la protéine tau possède 3 répétitions (R1, R3 et R4) ou 4 répétitions (R1, R2, R3 et R4). Ainsi, les différentes répétitions du domaine de liaison aux MTs sont capables de se lier à plusieurs hétérodimères de tubuline et de polymériser les MTs. Adaptée de (Goode, Denis et al. 1997).
III. C. 2. Polarité cellulaire
La protéine tau intervient dans de multiples processus cellulaires de part sa composition et sa localisation cellulaire, mais surtout grâce à sa liaison aux MTs. Ainsi, en régulant la dynamique des MTs, la protéine tau peut agir au niveau de la polarité cellulaire et du transport axonal. En effet, la protéine tau stabilise les extrémités (+) et (-) des MTs, ce qui est essentiel au maintient de la polarité neuronale (Caceres and Kosik 1990). De plus, tau est d’avantage exprimée dans le compartiment