Identification of the molecular origins of disease in a cohort of patients with suspected congenital disorders of glycosylation (CDG)
196
0
0
Texte intégral
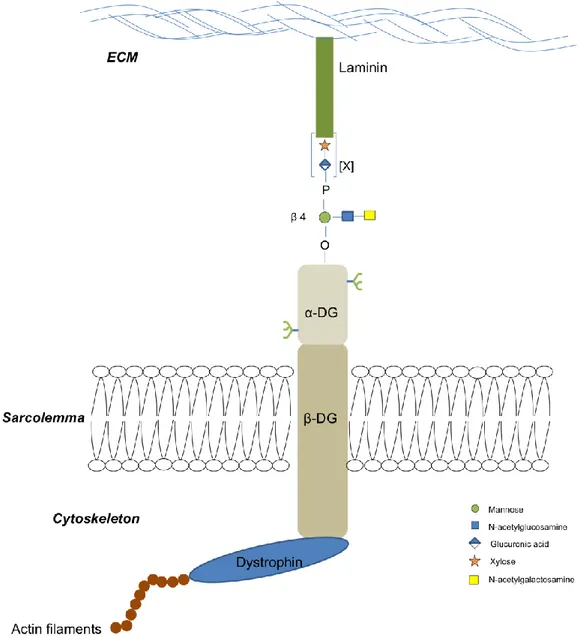
Figure
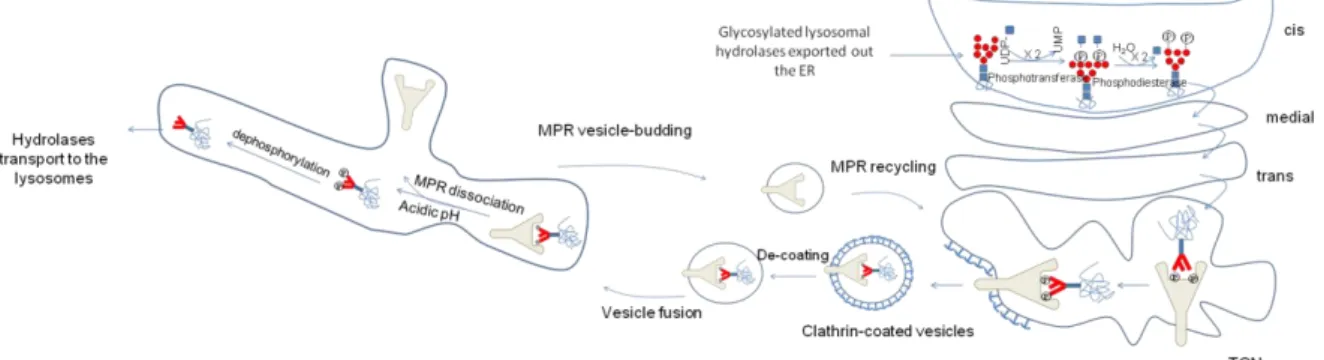
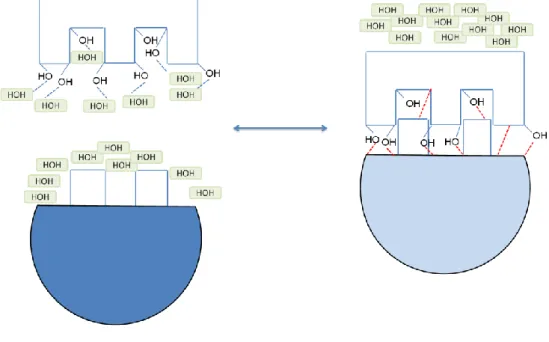
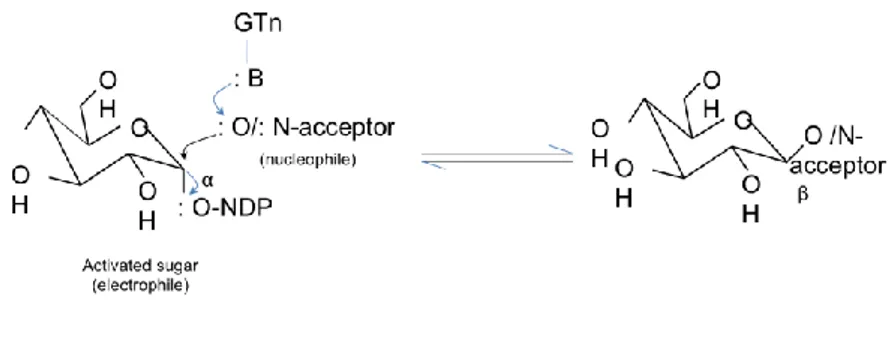
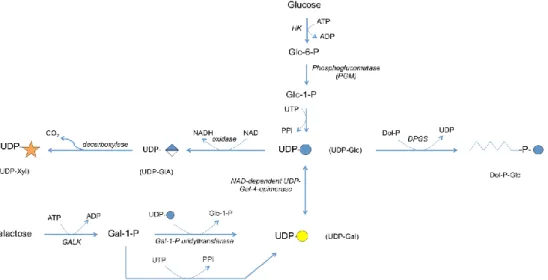
+7

Documents relatifs