REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE
SCIENTIFIQUE UNIVERSITE BATNA 1
INSTITUT DES SCIENCES VETERINAIRES ET DES SCIENCES AGRONOMIQUES
MEMOIRE
Pour l’obtention du diplôme de
MAGISTER
Filière :
Sciences vétérinaires
Option :
Histologie et histopathologie des animaux domestiques
Présenté par :
DOUAKHA Meriem Hanene
THEME
JURY Grade et Université
-Président : AISSI Adel
Prof. Université Batna 1
-Examinateur : BENHAMZA Louiza
Prof. Université Constantine 1
-Examinateur : KHENENOU Tarek
MCA. Université Souk Ahras
-Rapporteur : BENNOUNE Omar
Prof. Université Batna 1
Année universitaire : 2017/2018.
UTILISATION DES TECHNIQUES
HISTOMORPHOMETRIQUES POUR
L’EVALUATION DE LA MICROARCHITECTURE
OSSEUSE CHEZ LE POULET
REMERCIEMENT
Je tiens à remercier en premier lieu mes parents et mes frères
qui m’ont soutenu tout au long de mes études
Mon encadreur et tous mes professeurs qui ont fait de moi la
scientifique que je suis
Sans oublier mes amis et mes collègues qui m’ont encouragé
Et en fin, un remerciement particulier à tous ceux qui œuvrent
pour la noble cause d’offrir le savoir à tous
SOMMAIRE
LISTE DES FIGURES LISTE DES TABLEAUX LISTE DES ANNEXES
LISTE DES ABBREVIATIONS
INTRODUCTION 1
CHAPITRE I : BIBLIOGRAPHIE
1. HISTOLOGIE OSSEUSE : 3
1.1.Particularités du squelette chez la volaille. 3
1.2.Structure de l’os : 3
1.2.1. Os compact et spongieux. 4
1.2.2. Os médullaire. 4
1.2.3. Os primaire et secondaire 5
1.2.4. Système lamellaire. 6
1.2.5. Architecture des os longs. 6
1.3.Constituants de l’os : 6 1.3.1. Matrice. 6 1.3.2. Cellule. 7 1.3.3. Périoste et endoste. 8 1.3.4. Moelle osseuse. 8 2. PHYSIOLOGIE OSSEUSE : 9 2.1.Formation de l’os : 9 2.1.1. Ostéogénèse 9 2.1.2. Croissance. 11 2.2.Remodelage et réparation : 13 2.2.1. Remodelage. 13
2.2.2. Réparation des fractures. 14
2.3.Métabolisme minérale : 14
3. PATHOLOGIES MAJEURES DE L’OS : 15
3.1.Ostéoporose. 15
3.2.Chondrodystrophie. 16
3.3.Dyschondroplasie tibiale 16
3.5.Syndrome de malabsorption 17
4. HISTOTECHNOLOGIE OSSEUSE 17
4.1.Coupes décalcifiées. 17
4.2.Coupes non décalcifiées. 18
4.3.Artéfacts sur coupe décalcifiée. 20
4.4.Coloration spécifiques. 21 4.4.1. Coloration du calcium 21 4.4.2. Coloration de la moelle 21 4.4.3. Marquage au fluorochrome 21 4.4.4. Vascularisation 22 4.5.Analyse histomorphometrique 22 4.6.Microscopie électronique 23 4.7.Autres techniques 24 4.7.1. Minéralisation. 24 4.7.2. Radiographie. 24 4.7.3. Test biomécanique. 25
CHAPITRE II : MATERIEL ET METHODES
1. Matériel animal. 27
2. collecte d’échantillons. 27
3. Préparation des coupes histologiques 27
3.1.Fixation 27
3.2.Décalcification 28
3.3.Mise en block 28
3.4.Coupes et coloration 29
4. Poids sec et minéralisation 31
5. Réalisation des mesures 32
5.1.Mesures métriques et pondérales 32
5.2.Mesures histologiques 32
5.3.Méthodes statistiques 35
CHAPITRE III : RESULTATS ET DISCUSSION
1. Mesures métriques et pondérales. 36
1.1.Poulets de chair 36
1.1.2. Poids net et longueur du fémur 37
1.1.3. Cendres 38
1.2.Poules pondeuses 38
1.2.1. Poids vif des animaux 39
1.2.2. Poids net et longueur du fémur 39
1.2.3. Cendres 40
2. Mesures histologiques sur la diaphyse 40
2.1.Poulets de chair 40 2.1.1. Diamètre 41 2.1.2. Cortex total 41 2.1.3. Périoste 41 2.2.Poules pondeuses 41 2.2.1. Diamètre 42 2.2.2. Cortex total 42 2.2.3. Périoste 43
3. Mesure des volumes de l’os trabéculaire et médullaire 44
3.1.Os trabéculaire 44
3.2.Os médullaire 46
4. Mesures histologiques sur les épiphyses 49
5. Mesures cellulaires 51
5.1.Cellularité 51
5.2.Dénombrement d’ostéoclates. 52
6. Qualité des coupes histologiques 53
6.1.Déplacement et rétrécissement 54 6.2.Effritements 55 6.3.Plis 58 6.4.Os impactés 58 6.5.Détachement du cartilage 59 6.6.Autres 60 6.6.1. Défauts de montage 60
6.6.2. Les tissus étrangers 61
6.6.3. Défauts de coloration 62
CONCLUSION 65 ANNEXES
REFFERENCES RESUME
LISTE DES FIGURES
CHAPITRE I : BIBLIOGRAPHIE
Figure 1 : squelette d’un poulet adulte (McLelland, 1990). 3 Figure 2 : os compact, humérus décalcifié de poulet (Bacha et Bacha, 2000). 4 Figure 3 : os médullaire : tête d’humérus de poulet (Aughey et Frye, 2001). 5 Figure 4 : ostéoclaste multinuclées dans une lacune de Howship sur un os
médullaire.
7
Figure 5 : moelle osseuse du poulet, coloration Giemsa (Bacha et Bacha, 2000). 9 Figure 6 : ossification intra-membraneuse (Gartner et Hiatt, 2002). 10 Figure 7 : Cartilage de conjugaison avec ses 5 zones d’élongation. Humérus
décalcifié chat (Bacha et Bacha, 2000).
12
Figure 8 : ligne cimentaire (Thompson, 2007). 13
Figure 9 : régulation du métabolisme calcique (Larbier et Leclercq, 1992). 15 Figure 10 : Dyschondroplasie tibiale (Guérin et al, 2011). 16 Figure 11 : os non décalcifié obtenu par meulage (X270) (Gartner, 2000). 19 Figure 12 : biopsie d’une crête iliaque non décalcifié au Goldner trichrome (X400)
(An et Gauber, 2003).
21
Figure 13 : Marquage multiple au fluorochrome à 5 jours d’intervalle du tibia d’une ratte de 12 mois sous lumière bleu (450–490 nm) (X400) (Erben, 2003).
22
Figure 14 : microphotographie de la diaphyse fémorale de rat adulte montrant la vascularisation après micro injection d’ancre de chine. Coupe non décalcifié (200µm) d’épaisseur (Villanueva, X10) (Jiang et al, 2003).
22
Figure 15 : mesure directe des paramètres trabéculaire par l’utilisation d’oculaires munis de 2 motifs différents (Carbonane et al, 2005).
23
Figure 16 : MEB d’un os après élimination de la moelle et les cellules, la partie centrale montre une large expansion lisse d’une surface quiescente (X350) (Gruber et Wiggins, 2003).
24
Figure 17 : image radiographique du tibia de poulet de chair (Amorose et al, 2013). 25 Figure 18 : courbe relation contrainte-déformation (Goulard, 2008). 26
CHAPITRE II : MATERIEL ET METHODES
Figure 19 : étapes de préparation des coupes. 30
Figure 20 : étuve. 31
Figure 21 : four à moufle. 32
Figure 22 interface du logiciel ImageJ version 1.48. 33
Figure 23 : les zones de croissance au niveau du cartilage de conjugaison. 34
Figure 24 : schéma suivi pour le comptage cellulaire. 35
CHAPITRE III : RESULTATS ET DISCUSSION Figure 25 : comparaison du cortex au niveau de la diaphyse entre les lots (H&E,
X10).
43
Figure 26 : détachement du périoste, coupe transversale de la diaphyse (H&E, X10). 44 Figure 27 : distribution de l’os médullaire chez les lots de pondeuses (H&E, X4). 47 Figure 28 : os médullaire accolé au cortex chez les pondeuses en fin de ponte (H&E,
X10).
48
Figure 29 : absence du second centre d’ossification, coupe longitudinal de l’épiphyse (H&E, X4).
50
Figure 30 : comptage des ostéoclastes au niveau de la diaphyse. 52 Figure 31 : comptage des ostéoclastes au niveau de l'épiphyse proximale. 52 Figure 32 : Comptage des ostéoclastes au niveau de l'épiphyse distale. 53
Figure 33 : les artéfacts les plus rencontrés. 54
Figure 34 : Artéfact de rétraction de la cellule dans sa lacune (H&E, X100). 54 Figure 35 : Artéfact de déplacement et séparation des tissus (H&E, X4). 55 Figure 36 : les types d’artéfacts d’effritements rencontrés (H&E, X4). 57
Figure 37 : artéfact de plis (H&E, X4). 58
Figure 38 : artéfact d’impaction osseuse (H&E, X10). 59 Figure 39 : artéfact de détachement du cartilage articulaire (H&E, X4). 59 Figure 40 : artéfact de montage : bulles d’air emprisonnées dans la résine (H&E,
X10).
60
Figure 41 : artéfact de montage : bulle d’air entre lame et lamelle mais en dehors du tissu (H&E, X4).
61
Figure 42 : tissus étrangers (H&E, X4). 62
Figure 43 : défauts de coloration (H&E, X4). 63
LISTE DES TABLEAUX
CHAPITRE I: BIBLIOGRAPHIE
Tableau 1 : Types d’artéfacts trouvés lors de réalisation de coupes décalcifiées et leurs causes.
20
CAPITRE II : MATERIEL ET METHODES
Tableau 2 : Répartition des animaux en lots. 27
CHAPITRE III : RESULTATS ET DISCUSSION
Tableau 3 : Mesures métriques et pondérales chez les poulets de chair. 35 Tableau 4 : Mesures métriques et pondérales chez les pondeuses. 39 Tableau 5 : Mesures sur le cortex chez les poulets de chair. 40
Tableau 6 : Mesures sur le cortex chez les pondeuses. 42
Tableau 7 : Valeurs des grands et petits diamètres de chaque lot. 42
Tableau 8 : Pourcentage de la surface des trabécules. 44
Tableau 9 : Pourcentage de la surface de l’os médullaire 46 Tableau 10 : Epaisseur cortex et os médullaire accolé au cortex chez les poules en
fin de ponte.
48
Tableau 11 : Longueurs des zones de croissance sur l’épiphyse proximale. 49 Tableau 12 : Longueurs des zones de croissance sur l’épiphyse distale. 49 Tableau 13 : Pourcentage de tissu adipeux au niveau de la moelle. 51
LISTE DES ANNEXES
Annexe 1 : Etapes de la mise en block.
Annexe 2 : Protocole de coloration.
Annexe 3 : Mesures métriques et pondérales.
Annexe 4 : Mesures histomorphométriques effectués sur la diaphyse.
Annexe 5 : Mesures histomorphométriques effectués sur les épiphyses.
LISTE DES ABRREVIATIONS
H&E : hématoxyline éosine
Ca : calcium µCT : micro-computed-tomography 3D: 3 dimensions °C: degré Celsius J : jour h : heur C : poules de 28 jours CC : poules de 56 jours
P : poules en début de ponte
1
INTRODUCTION
L’élevage avicole retient l’attention par son court cycle de production; il est considéré comme un moyen facile pour se procurer des protéines animales accessibles à une large tranche sociale, il faut souligner que l’élevage connaît un grand essor dans le monde y compris l’Algérie.
Il convient de noterque la réduction de l’âge de mise sur le marché, et l’augmentation disproportionnée de la masse musculaire du poulet de chair ont contribué à une forte apparition d’anomalies du squelette (Lilburn, 1994 ; Pitsillides et al., 1999).
Chez la pondeuse, la grande fragilité osseuse est le résultat d’une forte production d’œufs sur des périodes prolongées. La fréquence des fractures et le bien-être des animaux sont devenus alors un problème majeur dans cette industrie (Riczu et al., 2004 ; Pitsillides et al., 1999).
Pour arriver à des résultats de haute production, la bonne gestion de l’élevage constitue un élément essentiel. Cependant, en Algérie, le développement de cette filière est marqué par plusieurs contraintes dont, essentiellement, l’application des bonnes pratiques de l'élevage par les éleveurs. Beaucoup de recherches ont été faites sur les performances zootechniques, mais peu sont celles qui traitent de l’état du squelette dans nos élevages.
La force de résistance de l’os est due à son volume, mais aussi à sa microarchitecture qui est responsable de la variabilité de force trouvée à son niveau (carbonare et al., 2000), ceci montre l’intérêt de l’utilisation de l’analyse histomorphométrique notamment avec les avancées technologiques, les logiciels de traitement d’image offrent désormais des données plus précises, et demandent moins de technicité.
Selon Turner, (1999) la fragilité osseuse chez les pondeuses n’est remarquée que lors du ramassage et le transport vers les abattoirs. L’histomorphométrie est basée sur des mesures quantitatives des structures microscopiques (Egan et al., 2012), elle permet donc de détecter les anomalies cachées.
Il faut signaler cependant que des difficultés sont rencontrées au cours de la préparation histologiques, notamment pour les coupes de routine où la phase de décalcification représente un challenge supplémentaire dans la préservation de la structure tissulaire (Srinivasyaiah et al., 2016). En plus les artéfacts sont souvent mal interprétés,et sont considérés comme lésions
2 Chez les poulets de chair, le fémur représente le maillon faible dans le développement des os longs, ce qui fait de lui le siège de nombreuses anomalies(Lilburn, 1994). Il constitue aussi la principale source de calcium osseux pour la formation des coquilles d’œufs, il devient ensuite fragile et poreux chez les pondeuses (Almeida et Bruno, 2006).
Pour ces raisons, notre choix s’est porté sur le fémur afin d’avoir un aperçu sur l’état du squelette des animaux de nos élevages, ainsi que sur le développement de leurs os et de débusquer un éventuel déséquilibre alimentairesous les conditions d’élevage et de commercialisation usuelles. Par ailleurs, notre étude a porté sur des poulets de chair et des poules pondeuses à différents stades du cycle de productionafin de suivre l’évolutionde la microarchitecture osseuse.
Nous avons pris chez les pondeuses, les animaux en début et fin de ponte ; Chez les poules de chair, ils sont sacrifiés lorsqu’ils ont atteint la mi-temps du cycle de production, et à l’âge de mise sur le marché.
Les coupes réalisées sont celles à la paraffine, colorées à l’hématoxyline éosine (H&E) ; vu la simplicité de la technique et sadisponibilité dans le laboratoire. En plus l’H&E nous offre une large gamme de rouge qui permet d’identifier les différentes structures histologiques, et elle permet aussi de localiser les dépôts de calcium qui prennent une couleur foncée (Lai et Lu, 2012). Les mesures sont réalisées par le logiciel ImageJ.
Les principaux objectifs de cette étude sont les suivants :
- Mesurer les paramètres histomorphométriques de la structure osseuse du fémur.
- Trouver une relation entre les valeurs de ces paramètres, l’état physiologique des animaux et les données anatomiques.
- Identifier les difficultés lors de la réalisation des coupes histologiques à la paraffine décalcifiées et coloré à l’H&E.
- Identifier les difficultés quant à l’utilisation de l’histomorphométrie sur ces coupes avec le logiciel ImageJ.
Chapitre I :
Bibliographie
3
CHAPITRE I : BIBLIOGRAPHIE
1. HISTOLOGIE OSSEUSE :
1.1.Particularités du squelette chez la volaille :
Comparé aux mammifères, le squelette des oiseaux est plus léger avec une plus haute teneur en phosphate de calcium. La plupart des vertèbres du corps sont fusionnés ; Les os des membres sont simplifiés par la fusion ou la suppression de certains os (McLelland, 1990). Le sternum, forme la majorité du plancher thoraco-abdominal, et porte une carène (bréchet) qui sert d’insertion aux muscles du vol (Guérin et al., 2011) [Figure 1].
Figure 1 : squelette d’un poulet adulte (McLelland, 1990).
Une autre particularité aux oiseaux est celle d’avoir des os pneumatisés, ce sont des os pénétrés par des sacs aériens ; Les os concernés sont généralement ceux du crâne, des vertèbres, de la ceinture et des parties proximales des membres (Baumel et Witmer, 1993). La faible densité osseuse chez les oiseaux permet le vol et la flottaison sur l’eau (Sakas, 2002).
1.2.Structure de l’os :
Les os sont classés selon leurs formes anatomiques en os : long, court, plats, irréguliers et sésamoïdes (Gartner et Hiatt, 2006).
Macroscopiquement on distingue deux types de tissu osseux : compact et spongieux, qui ne sont cependant pas séparés par des limites nettes ; La structure microscopique de l’os mature est la même quelque soit le type, et il est appelée tissu osseux lamellaire (Dhem, 2004).
4
1.2.1. Os compact et os spongieux :
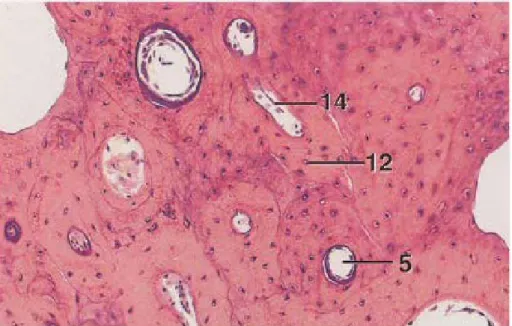
L’os compact apparaît comme une masse solide à l’œil nu, il couvre l’extérieur des os et forme la gaine des os longs ; quant a l’os spongieux (ou trabéculaire) lui est localisé à l’intérieur, il est constitué d’un labyrinthe de spécules osseuses entre lesquelles on trouve des espaces remplis de tissu conjonctif lâche ou de moelle et au moins un vaisseau sanguin. Ces espaces ne sont visibles dans l’os compact qu’a l’aide d’un microscope (Pakurar et Bigbee, 2004) [Figure 2].
Figure 2 : os compact, humérus décalcifié de poulet : 5-canal de Havers 12-ostéocyte 14-canal de Volkmann (Bacha et Bacha, 2000).
1.2.2. Os médullaire :
Présent chez les oiseaux femelles uniquement. Ce sont des spécules qui poussent de la surface de l’endoste à l’intérieur de la cavité médullaire durant la période de reproduction (Baumel et Witmer, 1993). C’est l’os qui subit le plus l’effet de l’œstrogène (Dacke, 1999 ; Dacke et al.,1993). Il sert comme réserve minérale hautement labile pour la formation de la coquille de l’œuf
(Baumel et Witmer, 1993).Il contient moins de collagène que l’os corticale, et il est richement calcifié (Dacke et al., 1993) il possède une structure isotropique : sans orientation de collagène
5
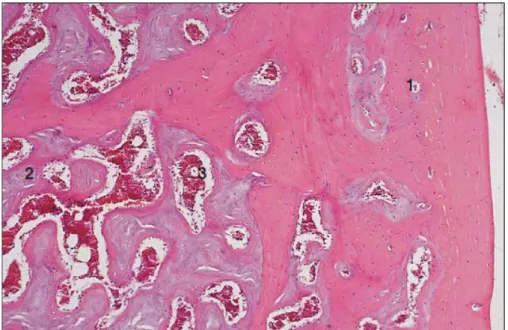
Figure 3 : os médullaire : tête d’humérus de poulet : 1-os cortical éosinophile 2-os médullaire basophile
3- moelle rouge (Aughey et Frye, 2001).
1.2.3. Os primaire et secondaire :
L’os primaire apparait durant le développement fœtal et lors de réparation de fractures, il est remplacé par l’os secondaire (ou lamellaire) sauf à certain endroits : Suture du calvarium, zone d’insertion des tendons et les alvéoles dentaires (Gartner et Hiatt, 2006). Ces deux os peuvent donner et l’os compact et l’os spongieux; aussi l’os mature peut être déposé de novo, sans qu’il y est d’os primaire au préalable (Pakurar et Bigbee, 2004).
Sur des coupes colorées à l’H&E, on constate que la matrice de l’os immature est bleuâtre, par contre, celle de l’os mature est uniformément acidophile (Bacha et Bacha, 2000). Dans le premier cas : les fibres sont déposées aléatoirement, l’os est moins minéralisé, les cellules sont plus nombreuses de forme sphérique et sans arrangement ordonné (Pakurar et Bigbee, 2004).
Dans le second cas : la matrice est déposée en lamelles où les fibres sont arrangées en parallèles, l’os est mieux minéralisé, a moins de cellules, de forme plates et sont situées entre les lamelles (Pakurar et Bigbee, 2004).
Il faut comprendre qu’il n’y a pas de limites nettes entre os mature et immature d’un coté, et entre os spongieux et compact de l’autre, la conversion de l’os spongieux en compact est le résultat de dépôts répétés d’os entre les espaces. Quant à l’os immature c’est la conséquence d’une croissance appositionnelle d’os spongieux, alors que l’os mature est la conséquence d’un remodelage (Cormack, 2001).
6
1.2.4. Système lamellaire :
L’os spongieux possède un arrangement irrégulier de lamelles qui contiennent des lacunes abritant des ostéocytes, ces dernières sont nourris par diffusion à partir de la moelle (Gartner et Hiatt, 2006).
Dans l’os compact, les lamelles peuvent être soit arrangées concentriquement autour d’un canal longitudinal (canal de Havers) pour former un ostéone. Soit elles entourent les gaines osseuses (lamelles circonférentielles), ou bien elles remplissent les espaces (lamelles interstitielles). Ces trois types de lamelles sont souvent le résultat d’un remodelage osseux
(Aughey et Frye, 2001).
Au niveau de l’ostéone le canal de Havers est traversé perpendiculairement par les canaux de Volkmann ; ces systèmes de canaux permettent le passage des vaisseaux sanguins (Aughey et Frye, 2001).
1.2.5. Architecture des os longs :
Selon Pakurar et Bigbee (2004), l’os long est composé de :
- Diaphyse : composé d’os compact, creuse et tapissée par une fine couche d’os spongieux.
- Epiphyses : se sont les éminences aux deux bouts de la diaphyse, composées d’un cortex d’os compact ; l’os spongieux à l’intérieur abrite la moelle rouge.
- Métaphyse : région évasée entre la diaphyse et l’épiphyse.
- Cartilage de conjugaison : type hyalin, sépare l’épiphyse et la métaphyse. A la fin de la croissance, il n’en reste qu’une relique en forme de ligne.
- Cartilage articulaire : type hyalin, couvre les surfaces articulaires, ne possède pas de périchondre.
1.3.Constituants de l’os:
L’os est une forme rigide de tissu conjonctif, formé de cellules dans une matrice intercellulaire calcifiée, couvert d’un tissu spécialisé : périoste et endoste, et contenant une moelle (Aughey et Frye, 2001).
1.3.1. Matrice :
La matrice osseuse est constituée d’une partie organique qui lui offre la résistance à la tension et une partie inorganique qui lui offre la résistance à la compression (Eroschenko, 2008).
7 Avant qu’elle ne soit calcifiée la matrice est appelée ostéoide, la partie minérale constitue 70% du poids net de l’os et est représentée principalement par des cristaux d’hydroxyapathites. L’eau représente seulement 25% de la matrice, et le collagène représente 90% du contenu organique avec principalement du collagène type I et apparait rose dans les coupes colorées à l’H&E (Cormack, 2001).
La substance fondamentale est faite de protéoglycanes spécialement de chondroitine sulfate et kératane sulfate ; et de glycoprotéines tel l’ostéonectine, l’ostéopointine et l’ostéocalcine (protéine liant le calcium) (Dudek, 2000).
1.3.2. Cellules :
• Cellules ostéogénétrices :
Lesostéoblastesforment la matrice osseuse, ils dérivent des cellules mésenchymateuses, sont mononuclées de forme cuboïde à cylindrique et sont localisées sur les surfaces osseuses
(Eurell, 2004).
On trouve aussi sur la surface de l’os adulte, les cellules bordantes, se sont des cellules squameuses dont on pense qu’elles sont des ostéoblastes en repos (Eurell, 2004)
Quand les ostéoblastes sont complètement entourés par la matrice ils deviennent des ostéocytes, ces dernières maintiennent la matrice les entourant et sont capables de redevenir des ostéoblastes en cas de besoin. Ils sont localisés dans des lacunes et plusieurs prolongements irradient de leur corps cellulaires et passent dans des canalicules pour entrer en contact avec les cellules adjacentes (Eurell, 2004).
• Ostéoclastes :
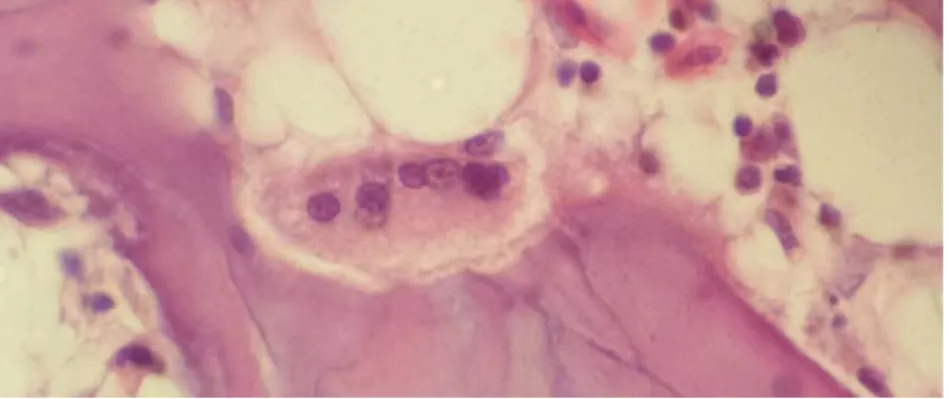
Ce sont des cellules multinuclées qui se trouve dans des lacunes de résorption (de Howship) dans les sites de remodelage (Aughey et Frye, 2001) [Figure 4].
8 Ils dérivent des monocytes, résorbe la matrice lors du remodelage osseux ou lorsqu’ il y a un besoin en calcium (Bacha et Bacha, 2000).
1.3.3. Périoste et endoste :
Le périoste est un tissu conjonctif particulier doué de potentiel ostéogénique, et possède donc un rôle important en cas de fracture. Il recouvre l’extérieur des os et varie de structure, dans les os en croissance on lui reconnait trois couches : une externe lâche (adventice), riche en vaisseaux qui alimente les canaux de Havers et de Volkmann. Une couche moyenne épaisse, fibro-élastique dont certaines fibres de collagène irradient dans l’os compact pour établir un lien fort (fibres de Sharpey) qui peuvent se calcifier. La troisième couche interne (cambium) ostéoprogénitrices est riche en cellules. Sur l’os formé la couche cambiale s’amincie et le périoste est riche en vaisseaux et nerfs (très sensible) (Dhem, 2004). Le périoste est absent des surfaces articulaires, des os sésamoïdes, et des zones d’insertion des tendons (Gartner et Hiatt, 2006).
L’endoste lui recouvre les cavités internes et les canaux de Havers. C’est une fine couche de cellules ostéoprogénitrices aplaties (Dhem, 2004).
1.3.4. Moelle osseuse :
On trouve la moelle dans le canal médullaire des os longs et dans les petites cavités de l’os spongieux (Mescher, 2013).Là où le tissu hématopoïétique prédomine la moelle active est rouge, là où le tissu adipeux prédomine la moelle inactive est jaune (Aughey et Frey, 2001). Chez le nouveau-né toute la moelle est rouge, chez l’adulte la moelle rouge se trouve au niveau des os plats, les os longs accumulent le gras deviennent jaunes, et perdent leur fonction hématopoïétique (Eroschenko, 2008). Sous certaines conditions comme une sévère hémorragie la moelle jaune se reconvertie en rouge (Mescher, 2013).
La moelle rouge contient un stroma de tissu conjonctif réticulaire, des cordes ou îles de cellules hématopoïétiques et des capillaires sinusoïdes ; le stroma est fait d’un réseau de cellules fibroblastiques et de fibres (Mescher, 2013). Les sinusoïdes offrent une route aux cellules nouvellement formées pour entrer dans la circulation (Cormack, 2001). La progression à partir d’une cellule souche vers une cellule fonctionnelle représente une lignée cellulaire et les formes matures prédominent sur les cellules immatures (Bacha et Bacha, 2000).
Chez le poulet l’érythropoïèse a lieu à l’intérieur des vaisseaux sinusoïdes [Figure 5] (Bacha et Bacha, 2000). Toutes les cellules sanguines gardent leurs noyaux, les cellules hémostatiques
9 sont les thrombocytes, les leucocytes sont : les hétérophiles, éosinophiles, basophiles, lymphocytes et monocytes (Clark et al., 2009). Les mégacaryocytes sont absents et les lymphocytes B se différencient au niveau de la bourse de Fabricius (Eurell, 2004).
Figure 5 : moelle osseuse du poulet, coloration Giemsa (Bacha et Bacha 2000).4-endothélium 5-éosinophile
6-érythrocyte immature 7- érythrocyte mature 8-hétérophile.
2. PHYSIOLOGIE OSSEUSE :
2.1.Formation de l’os :2.1.1. Ostéogénèse :
L’os se développe toujours par un processus de transformation d’un tissu conjonctif préexistant. Il existe deux types de développement osseux qui dépondent d’une différentiation spécifique des cellules dans deux microenvironnements différents, ces deux modes peuvent donner naissance à l’os spongieux et à l’os compact (Eurell et Frappier, 2006).
• Ostéogénèse intra-membraneuse :
Procédé par lequel se forment la pluparts des os plats [Figure 6], il suit les étapes suivantes
(Gartner et Hiatt, 2015):
- Les cellules mésenchymateuses en présence d’une vascularisation se transforment en ostéoblastes et produisent l’ostéoide de façon à donner l’os primaire. C’est le premier centre d’ossification.
- Avec la croissance appositionnelle et la calcification, les ostéoblastes sont emprisonnées dans leur matrice et deviennent des ostéocytes. Les centres de développement sont alors appelés des trabécules.
10 - La fusion des trabécules donne l’os spongieux.
- Les cellules mésenchymateuse non différenciées donnent les cellules hématopoïétiques de la moelle.
- L’endoste et le périoste se développent à partir des couches mésenchymateuse non ossifiées.
- Finalement l’os intra membraneux peut être converti en os lamellaire.
Figure 6 : ossification intra-membraneuse. (Gartner et Hiatt, 2002) Formation de trabécules et début de
formation d’ostéone .Flèche : ostéoblastes. Tête de flèche : ostéocytes
• Ostéogénèse enchondrale :
Les os des extrémités, des vertèbres, du pelvis et de la base du crâne se forment par ossification de modèles initiales de cartilage hyalin (Eurell et Frappier, 2006).
L’ossification enchondrale passe par deux étapes qui sont: Centre primaire d’ossification:
Se développe au centre de la diaphyse de la pièce cartilagineuse et suit les évènements suivants (Gartner et Hiatt, 2015) :
- La vascularisation du périchondre transforme les cellules chondrogéniques en ostéoprogénitrices qui se différencient en ostéoblastes. Cette région du périchondre est alors appelée périoste.
11 - Les ostéoblastes élaborent la matrice sous le périoste par ossification intra membraneuse
et forme le manchon périostique.
- Les chondrocytes au centre s’hypertrophient et dégénèrent, leurs lacunes se confluent et forment éventuellement les espaces médullaires.
- Les ostéoclastes créent des perforations dans le manchon qui permettent la pénétration d’un bourgeon périostique (vaisseaux sanguin, cellules mésenchymateuses et ostéoprogénitrices); Le cartilage des parois alors se calcifie.
- Les nouveaux ostéoblastes élaborent la matrice osseuse sur la surface calcifiée du cartilage, le manchon s’épaissie et s’allonge, puis les ostéoclastes viennent résorber le complexe cartilage-os en agrandissant la cavité médullaire.
- La répétition de ces séquences d’événement fait que la formation de l’os s’étend vers les épiphyses.
Centre secondaire d’ossification:
Les épiphyses cartilagineuses des os longs développent des centres additionnels d’ossification appelés centres secondaires (Eurell et Frappier, 2006). Les séquences d’événement sont similaires à ceux du centre primaire mais sans la formation de manchon périostique(Gartner et Hiatt, 2015). L’ossification ne remplace pas tout le cartilage épiphysaire les vaisseaux ne pénètrent ni le cartilage de conjugaison ni le cartilage articulaire (Eurell et Frappier, 2006).
2.1.2. Croissance :
Le taux de croissance le plus rapide prend lieu durant la période de post-éclosion et le stade juvénile, aussi cette vitesse de croissance diffère entre les deux sexes chez le poulet (Dacke, 1999).
Contrairement au cartilage, l’os croit seulement par apposition. Parce que sa matrice se minéralise rapidement, la croissance interstitielle est impossible, les changements de taille ou de forme se font par ajouts ou élimination de couches (Eurell et Frappier, 2006).
• Croissance en longueur :
Prend lieu au niveau du cartilage de conjugaison qui inclut cinq zones [Figure 7] qui sont comme suit (Dudek, 2000) :
- Zone de réserve : contient des chondrocytes quiescents.
- Zone de prolifération : contient des chondrocytes en mitose et forment des groupes isogènes.
12 - Zone d’hypertrophie : contient des chondrocytes hypertrophiés qui sécrètent la
phosphatase alcaline pour augmenter les niveaux de calcium et de phosphore.
- Zone de calcification : contient des chondrocytes morts et une matrice cartilagineuse calcifier appelée spicules.
- Zone d’ossification : contient des cellules ostéoprogénitrices qui s’amassent sur les spécules et se différencient en ostéoblastes, ces dernières déposent de l’os sur la surface des spécules et forment un spécule mixte.
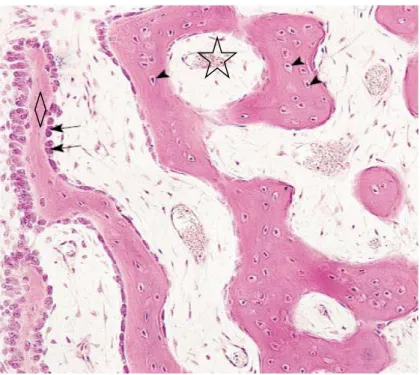
Figure 7 : Cartilage de conjugaison avec les 5 zones d’élongation: 1-moelle, 2-matrice osseuse, 3-cartilage
calcifié 4-ostéoblastes, 5-ostéocytes, 6-os spongieux diaphysaire, 7-os spongieux épiphysaire, 8-zone de calcification, 9-zone d’hypertrophie, 10-zone de multiplication, 11-zone d’ossification, 12-zone de réserve,
Humérus décalcifié de chat (Bacha et Bacha, 2000).
D’après (Eurel et Frappier, 2006) la spongiosa primaire (spécules mixtes) est couverte de nombreux ostéoclastes qui réduisent le nombre de trabécules. Plus loin dans la métaphyse on trouve des spécules sans noyau cartilagineux et sont appelées spongiosa secondaire.
13 • Croissance en diamètre :
L’os trabéculaire formé au niveau de la ceinture diaphysaire est converti en os compact composé d’ostéones primaires. La résorption et la formation simultanées, la première au niveau de l’endoste et la deuxième au niveau du périoste, aboutissent à l’élargissement de la cavité médullaire. Le principe à noter est que la croissance en longueur est le résultat d’une ossification enchondrale, alors que la croissance en diamètre est le résultat d’une ossification intra-membraneuse. Chaque type de croissance répond à différentes règles et peut être affecté séparément sans aucune influence de l’un sur l’autre (Eurell et Frappier, 2006).
2.2.Remodelage et réparation : 2.2.1. Remodelage :
Tout au long de la vie, l’os vieux ou défaillant est constamment renouvelé, ce remodelage prend lieux à un niveau local par coordination entre ostéoblastes et ostéoclastes (unité de remodelage osseux) qui nécessite la médiation d’une interaction complexe entre les hormones systémiques et les cytokines locales. Ceci permet un remplacement des structures internes sans altération de la forme ni de la fonction en générale (Thompson, 2007).
L’ostéone primaire est remplacé par l’ostéone secondaire, plus grand avec un liseré de ligne de cément [Figure 8] (Eurell et Frappier, 2006).
Figure 8: lignes cimentaires, flèches verticales : lignes de quiescence, flèches horizontales : ligne de revirement (Thompson, 2007).
14
2.2.2. Réparation des fractures :
Contrairement aux autres tissus qui guérissent par cicatrisation, l’os est capable de se régénérer et de reprendre son état initial. Le processus suit les étapes suivantes (Thompson, 2007) :
- l’évènement initial est la formation d’un hématome entre les deux extrémités de la fracture, accompagné par une nécrose ischémique due à l’arrêt de l’irrigation et conduisant à une réponse inflammatoire aigue.
- les cellules mésenchymateuses de l’endoste et du périoste prolifèrent et forment un callus de tissu conjonctif lâche puis se différencient en chondrocytes donnant un cartilage hyalin. - entre temps les ostéoclastes éliminent l’os nécrosé ; et sur la surface des extrémités,
commence une néo-ostéogénèse.
- une néo-vascularisation conduit alors à l’ossification enchondrale du pont cartilagineux, puis cet os primaire est remplacé par de l’os lamellaire. Cette dernière phase peut prendre des mois à des années.
- Le processus est plus rapide et plus complet chez les jeunes.
2.3.Métabolisme minéral :
Le calcium est le minéral le plus abondant au sein de l’organisme, très utilisé chez les animaux en production (squelette et œufs) et a un rôle très important dans la régulation de nombreuses fonctions (nerveuses, musculaires…). En faible concentration dans les liquides corporels, sa régulation est d’une grande précision [Figure 9]. Le phosphore lui constitue 16-17% des cendres osseuses, à l’état ionisé il est surtout abondant dans les liquides intracellulaires. Sa régulation se fait à 2 niveaux : os et rein, par la parathormone [Figure 9]. Une carence de phosphore conduit au rachitisme, celle du calcium n’affecte la croissance que chez le très jeune. Aussi d’autres carences en oligoéléments tel le cuivre, le zinc et le manganèse peuvent conduire à des troubles osseux (Larbier et Leclercq, 1992).
La provitamine D3 est secrétée chez le poulet par la glande uropygienne, elle passe ensuite
15
Figure 9: régulation du métabolisme calcique (Larbier et Leclercq, 1992).
3. PATHOLOGIES MAJEURES DE L’OS :
Les affections non infectieuses du squelette des volailles peuvent être distinguées en deux grandes catégories : celle associées à un défaut de minéralisation, entre autre rachitisme et syndrome de la pondeuse ; et celle qui, marquées par des altérations du cartilage de conjugaison, peuvent être considérées comme des chondrodystrophies (D’Autheville, 1979).
3.1.Ostéoporose :
Associée à ce qui est appelée fatigue de la poule en cage, c’est un trouble majeur chez les pondeuses. Elle résulte d’une perte progressive des structures osseuses minéralisées conduisant à une plus grande susceptibilité aux fractures et même à un collapsus de la colonne vertébrale avec paralysie. Elle serait due à une diminution de la formation osseuse avec augmentation de la résorption qui accompagne l’augmentation des œstrogènes à la maturité sexuelle en association avec d’autres facteurs dont la prédisposition génétique, la nutrition et le mode d’élevage. A ne pas confondre avec l’ostéomalacie où la matrice osseuse s’épaissie et se trouve faiblement minéralisée (Whitehead, 2002).
16
3.2.Chondrodystrophie :
La chondrodystrophie est initialement une lésion du cartilage de conjugaison dont l’épaisseur diminue. La croissance en longueur des os de la patte (tibia et métatarse) est réduite mais la minéralisation est normale ; secondairement ces os peuvent être épaissis et cintrés. Il y a enflure importante du talon, rejet latéral du métatarse de façon uni- ou bilatérale et, quelquefois, glissement du tendon hors du condyle articulaire(Sauveur, 1988).
3.3.Dyschondroplasie tibiale :
C’est une lésion du squelette fréquente chez les poulets de chair, identifiée comme une large masse de cartilage avasculaire provenant du cartilage de conjugaison [Figure 10]; elle serai due à une dégradation incomplète des chondrocytes (Cook et al., 1994).
Figure 10 : Dyschondroplasie tibiale (Guérin et al., 2011).
La masse est constituée de chondrocytes transitionnelles caractérisées par de petites cellules ovoïdes et une plus grande quantité de matrice qu’un cartilage hypertrophique normale, reflétant une incapacité des cellules à se différencier proprement. Elle serait associée à la vitesse de croissance vu sa disparition presque complète chez les poulets sous restriction alimentaire (Thorpet al, 1995).
17
3.4.Nécrose de la tête fémorale :
Aussi appelée fracture épiphysaire, cette maladie correspond à la dégénérescence du col et de la tête du fémur (séparation du cartilage articulaire au niveau de la tête épiphysaire du fémur). Elle entraîne des douleurs internes associées à une grande gêne ambulatoire, avec ulcères de la peau sur la zone cutanée correspondante (hanche galeuse). Les lésions ne sont souvent constatées qu’à l’abattoir. Une densité importante des animaux dans le bâtiment, associée à une hygrométrie excessive seraient des causes favorisantes de l’affection (Guérin et al, 2011).
3.5.Syndrome de malabsorption :
Reçu plusieurs appellations (pale bird syndrome aux USA. Brittlebonedisease au Pays Bas. Runtingstuntingdisease UK…). Selon les syndromes observés et le pays où ils ont été décrits. Dans tous les cas, il est lié à un trouble digestif d’origine infectieuse (reproduit par inoculation du contenu digestif) dont les agents semblent être des virus agissant en commun
(Larbier et leclercq, 1992).
Guérin et al.,(2011) y associe un processus toxique (mycotoxines), et le décrit comme un
syndrome nutritionnel, qui semble avoir une transmission horizontale, se manifestant par des diarrhées précoces, persistantes, spumeuses et brunâtres contenant des débris alimentaires intacts, la mortalité reste faible 4% malgré un net retard de croissance.
Deux altérations successives des pattes peuvent apparaître dans les syndromes de malabsorption. La première, visible sur des poulets de deux semaines, présente toutes les caractéristiques du rachitisme avec défaut de croissance globale de l’animal. L’altération suivante est une lésion du fémur visible sur les poulets de 6 semaines et présentant les caractères d’une ostéoporose est la plus souvent décrite. Les syndromes de malabsorption concernent les lipides, les vitamines E et D3, le calcium et le phosphore. Le nettoyage et la désinfection des bâtiments entre deux bandes d’élevage jouent un rôle déterminant dans la prévention de la maladie (Sauveur, 1988).
4. HISTOTECHNOLOGIE OSSEUSE ET DETERMINATION DE LA MINERALISATION :
4.1.Coupes décalcifiées :
Les échantillons sont traités avec des agents qui réagissent avec le calcium : on a les acides qui solubilisent les sels de calcium, ils décalcifient rapidement mais détériorent la qualité de coloration après longue incubation. Les chélateurs retirent les ions de calcium, ils ont peu
18 d’effet sur le tissu non osseux mais le temps de décalcification est plus long (Guibas et al., 2014).
• Test de vérification de décalcification :
L’échantillon doit être immédiatement retiré de la solution décalcifiante après décalcification complète, sinon les qualités tinctoriales seront réduites, et pour vérifier la décalcification complète de l’échantillon trois méthodes peuvent être utilisées (Luna, 1968) :
- Radiographie : représente la meilleure méthode.
- Tests chimiques : 5ml de la solution décalcifiante est testé avec un mélange de 5ml d’hydroxyde d’ammonium et 5ml d’oxalates d’ammonium, si après 15-30 min la solution est trouble ; la décalcification est incomplète et la solution décalcifiante doit être changée et le test refait ultérieurement.
- Tests physiques : nécessite de l’expérience, se fait par fléchissement ou par appui et on note la résistance.
4.2.Coupes non décalcifiées :
Procédure spéciale pour l’évaluation du tissu osseux, dentaire et spécialement les spécimens contenant des implants métalliques. Les échantillons sont enrobés dans des plastiques ou résines tel : le glycol méthacrylate, méthyle méthacrylate ou résine de Spurr ; le but est d’avoir un médium de la même dureté que l’os, on peut alors obtenir des coupes fines car le milieu offre un meilleur support aux composants cellulaires [Figure 11]. Trois méthodes de coupe peuvent être utilisées : direct par utilisation d’un microtome robuste, sciage, et sciage-meulage (An et Gruber, 2003).
19
Figure 11 : os non décalcifié obtenu par meulage (X270), on observe le canal de Havers (C), les lacunes et les canalicules (flèche) (Gartner, 2000).
20
4.3.Artéfacts sur coupes décalcifiées :
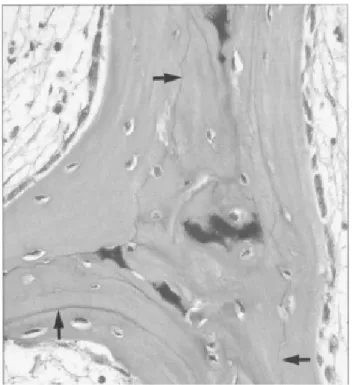
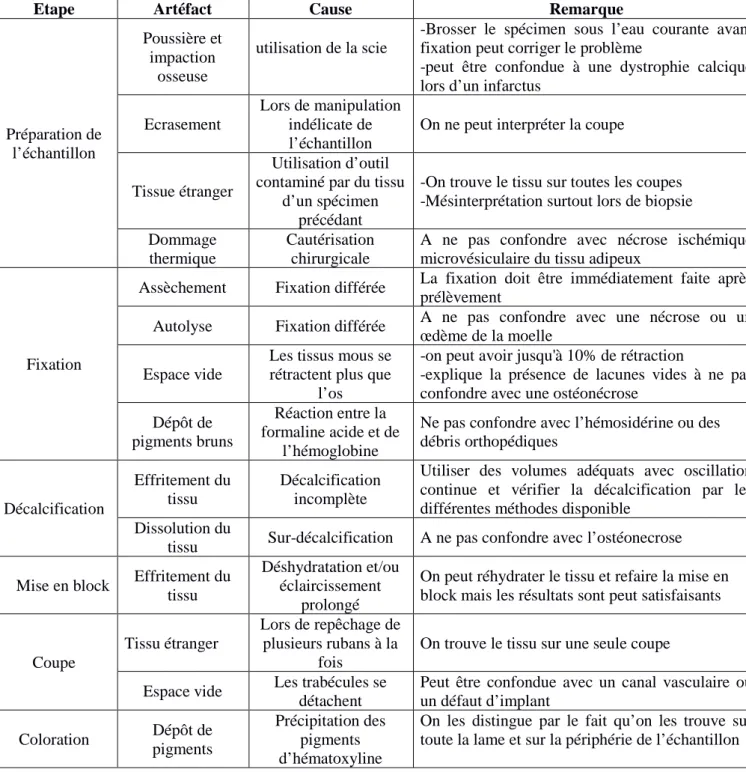
Les artéfacts peuvent être classés en majeurs et mineurs selon que le tissu d’origine est suffisamment présent pour établir un diagnostic correct ou non (Igho, 2012). Le tableau1 résume les artéfacts qu’on peut rencontrer à chaque étape selon Bauer, (2003):
Tableau 1 : types d’artéfacts trouvés lors de réalisation de coupes décalcifiées et leurs causes.
Etape Artéfact Cause Remarque
Préparation de l’échantillon Poussière et impaction osseuse utilisation de la scie
-Brosser le spécimen sous l’eau courante avant fixation peut corriger le problème
-peut être confondue à une dystrophie calcique lors d’un infarctus
Ecrasement
Lors de manipulation indélicate de l’échantillon
On ne peut interpréter la coupe
Tissue étranger
Utilisation d’outil contaminé par du tissu
d’un spécimen précédant
-On trouve le tissu sur toutes les coupes -Mésinterprétation surtout lors de biopsie Dommage
thermique
Cautérisation chirurgicale
A ne pas confondre avec nécrose ischémique microvésiculaire du tissu adipeux
Fixation
Assèchement Fixation différée La fixation doit être immédiatement faite après prélèvement
Autolyse Fixation différée A ne pas confondre avec une nécrose ou un œdème de la moelle
Espace vide
Les tissus mous se rétractent plus que
l’os
-on peut avoir jusqu'à 10% de rétraction
-explique la présence de lacunes vides à ne pas confondre avec une ostéonécrose
Dépôt de pigments bruns
Réaction entre la formaline acide et de
l’hémoglobine
Ne pas confondre avec l’hémosidérine ou des débris orthopédiques Décalcification Effritement du tissu Décalcification incomplète
Utiliser des volumes adéquats avec oscillation continue et vérifier la décalcification par les différentes méthodes disponible
Dissolution du
tissu Sur-décalcification A ne pas confondre avec l’ostéonecrose Mise en block Effritement du
tissu
Déshydratation et/ou éclaircissement
prolongé
On peut réhydrater le tissu et refaire la mise en block mais les résultats sont peut satisfaisants
Coupe
Tissu étranger
Lors de repêchage de plusieurs rubans à la
fois
On trouve le tissu sur une seule coupe Espace vide Les trabécules se
détachent
Peut être confondue avec un canal vasculaire ou un défaut d’implant Coloration Dépôt de pigments Précipitation des pigments d’hématoxyline
On les distingue par le fait qu’on les trouve sur toute la lame et sur la périphérie de l’échantillon
21
4.4.Coloration spécifiques :
4.4.1. Coloration du calcium :
La coloration Goldner trichrome et Von Kossa permettent de différencier l’ostéoide de la matrice minéralisée. Se font sur coupes non décalcifiés(Gruber et Ingram, 2003) [Figure 12]. Une autre coloration bien spécifique au calcium est la rouge alizarine S (Wulff, 2004).
Figure 12 : biopsie d’une crête iliaque non décalcifié au Goldner trichrome (X400). Os cortical (vert) ostéoide
(magenta) (An et Gruber, 2003).
4.4.2. Coloration de la moelle :
Les colorations de Romanovski (polychromatique de Giemsa) offrent un large arrangement de couleur qui permettent de différencier les précurseurs des cellules hématopoïétiques et de détecter ainsi les troubles sanguins et les tumeurs (Wulff, 2004).
4.4.3. Marquage au fluorochrome :
Utilisé pour examiner la croissance et le remodelage. Il nécessite des coupes non décalcifiés vu qu’il se base sur la formation de liaison du fluorochrome au calcium, spécialement au front de minéralisation à cause de la petite taille des cristaux d’apatites en ces lieux, ils sont alors observés après excitation au microscope à ultra-violet ou à la lumière bleu. L’administration du label ou fluorochrome se fait sur l’animal vivant, après leur incorporation, le label n’est relâché qu’après résorption de l’os, aussi plusieurs combinaison de labels peuvent être utilisées pour un aperçu chronologique du cycle osseux [Figure 13]. Pour les produits utilisés on peut citer la tétracycline, calceine, alizarine (Erben, 2003).
22
Figure 13 : marquage multiple au fluorochrome à 5 jours d’intervalle du tibia d’une ratte de 12 mois sous
lumière bleu (450–490 nm) (X400) (Erben, 2003).
4.4.4. vascularisation :
Des injections intraveineuses d’encre de chine avant fixation peuvent être utilisées pour visualiser la vascularisation [Figure 14](AN et Gruber, 2003).
Figure 14 : microphotographie de la diaphyse fémorale de rat adulte montrant la vascularisation après micro
injection d’ancre de chine. Coupe non décalcifier (200µm) d’épaisseur (Villanueva, X10) (Jiang et al., 2003).
4.5.Analyses histomorphométriques :
L’examen histomorphométrique des tissus est basé sur des mesures quantitatives des structures microscopiques. Il est utilisé pour fournir des informations sur les réponses cellulaires, les pathologies tissulaires et métaboliques. L’analyse des paramètres osseux requière trois types de mesures : surfaces, périmètre, et distances entre deux segments
23 d’intérêt. Ces trois mesures basiques peuvent être traduites en indices primaires dont : le volume tissulaire, le volume osseux, surface de minéralisation…etc. (Egan et al., 2012).
Pendant plusieurs années les mesures ont été réalisées à l’aide d’oculaires menu de motifs en
points ou en quadrillage [Figure 15] (Aaron et shore, 2003).
Figure 15: mesure directe des paramètres trabéculaire par l’utilisation d’oculaires muni de deux motifs différents (Carbonane et al., 2005).
Désormais l’utilisation et l’avancée des logiciels de traitement d’images ont permis plus d’automatisation (Egan et al., 2012) [Figure 22].
Aussi un besoin pour une standardisation de la terminologie s’est ressentie pour faciliter la communication entres les chercheurs. L’ASBMR (Américan society of bone and mineralresearch) a proposé un système qui a été adopté par le « Journal of bone and mineralresearch » comme part de ses instructions aux auteurs (Parfitt et al., 1987).
4.6.Microscopie électronique :
La microscopie électronique à balayage (MEB) montre la topographie osseuse et permet la visualisation de l’activité métabolique sur de larges surfaces [Figure 16], la surface osseuse est alors plate en zones de quiescence, avec des prolongations noueuses en zones de minéralisation et des dépressions festonnées en zones de résorption (Gruber et Wiggins, 2003). La microscopie électronique à transmission permet une caractérisation histologique, cristallographique et chimique utilisée pour l’évaluation du processus biologique et biophysique qui prend lieu à l’interface os-implant dans l’étude du comportement des implants à long terme (Hemmerlé, 2003).
24
Figure 16: MEB d’un os après élimination de la moelle et les cellules, la partie centrale montre une large
expansion lisse d’une surface quiescente (X350) (Gruber et Wiggins, 2003).
4.7.Autres techniques :
4.7.1. Minéralisation :
La mesure de la quantité de cendre dans le tibia est considérée depuis longtemps comme un bon représentant de la concentration de calcium et de phosphore, elle est recommandée par l’AOAC (Association officielle des chimistes analytiques). Vu sa croissance rapide le tibia est très sensible aux carences, la procédure appliquée fait appel à l’extraction des graisses avant incinération mais l’utilisation par l’auteur du tibia sans extraction, donne des résultats similaires (Yan et al., 2005).
4.7.2. Radiographie : • Densitomètrie optique :
Une technique pour déterminer le degré de minéralisation de l’os. La quantité de calcium et phosphore accumulée est comparée à celle de l’aluminium trouvé dans une échelle prédéterminée vu que la densité de l’aluminium et celle de l’hydroxyapatite sont similaires
25
Figure 17 : image radiographique du tibia de poulet de chair (Amorose et al., 2013).
• Microtomographie au rayon X (µCT) :
Technique d’imagerie 3D à haute résolution. Mesure directement la microarchitecture osseuse sans se baser sur la stériologie contrairement à l’histologie, en plus on peut analyser un volume plus large, elle est plus rapide, et non destructive ce qui permet de faire d’autres tests sur l’échantillon tels que les tests biomécaniques (Bouxsein et al., 2010). Cette technique utilise des rayons X atténués, les données prises à plusieurs angles servent à faire une reconstitution en 3D. Les données qu’elle génère sont en corrélation avec d’autres techniques plus invasives tel la minéralisation (Korver et al., 2004).
La résonance magnétique a une résolution équivalente à la µCT (jusqu’à 10µ pour des spécimens de moins de 10mm diamètre ex vivo) mais ne différencie pas l’ostéoide de la phase minéralisée. Le synchrotron qui utilise une source de rayon monochromatique permet une meilleure distinction entre l’ostéoide et la phase minéralisée mais peut générer jusqu'à 100GB de données (Parkinson et Fazzalari, 2011).
4.7.3. Test biomécanique :
Le squelette est sujet à différents types de forces (compression, tension, torsion…), une blessure peut être produite par application de l’une de ces forces à forte-magnitude, ou par la répétition de la charge à faible magnitude pendant une longue durée. Cette dernière est appelée fracture de stress ou de fatigue (Bankoff, 2012).
26 L’os possède une réponse aux forces mécaniques qui suit la loi de Wolff : l’os est déposé là où il est nécessaire (lors d’application de stress) et est résorbé là où il n’est pas nécessaire (lors d’immobilisation) (Thompson, 2007).
Les tests biomécaniques évaluent les propriétés structurales de l’os, les paramètres peuvent être déterminés sur os entier, un spécimen standard ou même sur des régions microscopiques du cortex et des trabécules par le biais de la nanoindentation(Ritchie et al., 2008).
Les propriétés mécaniques structurales sont décrites par des paramètres déterminés en examinant la relation entre la charge appliquée et la quantité de déformation qui prend place, représentée par la courbe charge-déformation [Figure 18] (Bankoff, 2012).
On reconnait à l’os deux caractéristiques : il est anisotropique (sa réaction change selon la direction de la charge appliquée), et visico-élastique (répond différemment selon la vitesse à laquelle la charge est appliquée et sa durée) (Bankoff, 2012).
Chapitre II :
27
CHAPITRE II : MATERIEL ET METHODES
1. Matériel animal :
L’étude s’est faite sur 20 individus femelles, réparties en quatre lots selon l’âge et le type de production comme suit :
Tableau 2: répartition des animaux en lots.
N° lot Abréviation Caractéristiques Nombre d’individus
1 C Poules type chair âgées de 28 jours 5
2 CC Poules type chair âgées de 56 jours 5
3 P Poules pondeuse en début de ponte de 5
mois 5
4 PP Poules pondeuses réformées d’1 an et 7
mois 5
L’étude a été réalisée dans la région de Guelma, durant la période d’Avril à Mai. Les poules type chair sont issues d’un même élevage. Tandis que les pondeuses proviennent de deux élevages différents.
2. Collecte d’échantillon :
Les animaux ont été pesés avant d’être sacrifiés par saignement et les valeurs du poids vif sont enregistrées. Les fémurs droit et gauche de chaque individu ont été prélevés. Le fémur droit est destiné aux coupes histologiques, alors que le gauche est congelé et gardé pour les mesures métriques et pondérales.
3. Préparation des coupes histologiques :
Afin de pouvoir faire des mesures histomorphométriques, nous avons réalisés des coupes histologiques. La méthode suivie est celle utilisée en routine c'est-à-dire : échantillon décalcifiées avec inclusion à la paraffine et colorée à l’hématoxyline éosine (H&E).
3.1.Fixation :
La fixation c’est faite immédiatement après la collecte des échantillons, le fémur droit est découpé en une diaphyse et deux épiphyses [figure 19-1]. Les différentes parties sont alors identifiées et immédiatement fixées dans une solution de formol tamponné à 10%.
28 La solution de formol est préparée selon la formule suivante adaptée de Luna, (1968) :
- 100 ml de formaline 37-40%. - 900 ml eau distillée.
- 4 g sodium phosphate monobasique.
- 6,5 g sodium phosphate dibasique (anhydre).
La durée de fixation varie suivant le lot avec la variation de la taille du spécimen. Les durées précises pour chaque lot sont détaillées dans l’ (annexe 1).
3.2.Décalcification :
Après un lavage à l’eau courante, la décalcification a été réalisée selon la technique décrite par An et Gruber (2003). Ainsi, une solution d’acide formique à 5% a été utilisée, sauf pour le premier lot où une solution à 5‰ a été utilisée vu la petite taille du spécimen et sa faible calcification. Les durées précises sont détaillées dans l’ (annexe 1). La décalcification adéquate a été vérifiée par test de fléchissement. [Figure 19-3].
Après décalcification, les os sont mis dans des casettes [figure 19-2] ; Pour cela ils sont coupés pour avoir une épaisseur de 4mm, la diaphyse est coupée transversalement, alors que les épiphyses sont coupées longitudinalement afin de mettre en évidence le cartilage de conjugaison. Les lots de poulets de chair de 56 jours et de poules pondeuses en fin de ponte ont été coupés et mis en casette avant décalcification pour accélérer le processus.
3.3.Mise en bloc :
La confection d’un bloc à la paraffine suit les étapes suivantes adaptées de An et Gruber, (2003), dont les durées précises sont dans l’ (annexe 1) :
• Déshydratation : faite par immersion dans des bains successifs d’alcool à concentration croissante, l’alcool utilisé est l’éthanol. Le but est de remplacer l’eau du tissu non miscible avec la paraffine.
• Eclaircissement : fait par immersion dans des bains de xylène, qui est miscible avec la paraffine et l’alcool, mais non miscible avec l’eau, d’où la nécessité de l’étape précédente. • Infiltration : faite par immersion dans de la paraffine mise à fusion à 59°C. cette étape
offre au tissu une résistance à la pression de coupe.
• La mise en bloc proprement dite : offre un meilleur soutien au spécimen, et une plus large surface de coupe [figure 19-5].
L’immersion dans les différents bains s’est faite grâce à un automate d’histologie BAVIMED [Figure 19-4]
29
3.4.Coupe et coloration :
Cette étape permet d’avoir des coupes fines qui montrent les détails tissulaires sous le microscope, la coloration permet plus de contraste entre les différentes structures :
• Les coupes sont réalisées à l’aide d’un microtome Leica Histocut 820 [figure 19-6]. Les rubans obtenues sont déplissés dans un bain-marie et étalées sur une lame identifiée et enduite d’une solution collante de Mayer préparée selon Luna, (1968) : 50 ml Blanc d’œuf + 50 ml glycérine, mixer et filtrer. La solution doit être conservée au réfrigérateur, puis diluer avant utilisation dans de l’eau distillée à raison de 1ml de la solution pour 20ml d’eau distillée.
• Après séchage à l’air, les lames sont colorées à l’hématoxyline-éosine (H&E) [figure 19-7] selon la procédure du labo adapté de Prophet et al., (1992) (annexe 2) .
• Enfin les lames sont nettoyées avant de procéder au montage, qui est la dernière étape et où les lames sont recouvertes d’une lamelle collé par une résine. La résine utilisée ici est le Baume De Canada.
30
Figure 19 : étapes de préparation des coupes.
1 2
3 4
5
31
4. Poids sec et minéralisation :
Selon la méthode citée par Hall et al., (2003), Les os sont nettoyés de tout tissu résiduel accolé et même de leur cartilage articulaire, pour cela les os sont mis dans un autoclave à 6,82 Kg de pression pendant 8-12 minutes, ils sont laissés à sécher puis sont pesés et les valeurs sont enregistrées comme poids net.
La deuxième étape consiste à les enfourner à 105°C durant 24h afin d’extraire l’eau, une étuve de marque ESCO Isotherm est utilisée [figure 20]. Les os sont alors pesés et les valeurs sont enregistrées comme poids sec.
Figure 20 : étuve.
La dernière étape consiste à les incinérés à 600°C durant 24h, elle c’est faite dans un four à moufle de marque Nabertherm [figure 21]; les valeurs pondérales sont enregistrées comme cendre osseuse.
32
Figure 21 : four à moufle.
L’utilisation des méthodes alternatives, sans extraction des lipides, comme recommandée par l’association officielle des chimistes analytique, offre des résultats comparables à la méthode recommandée (Yan et al., 2005).
5. Réalisation des mesures :
5.1.Mesures métriques et pondérales :
Les mesures sont réalisées sur le fémur gauche. Selon Park et al, (2003), la réfrigération et congélation n’ont aucun effet sur la cendre osseuse.
Les mesures métriques ont été réalisées à l’aide d’un pied-à-coulisse, nous avons mesuré la grande longueur et les deux diamètres (grand-petit) qui sont mesurés au centre de la diaphyse. Les mesures pondérales réalisées sont :
• Poids vif de l’animal. • Poids net du fémur. • Poids sec du fémur.
• Poids de la cendre osseuse du fémur.
5.2.Mesures histologiques :
Les coupes ont été observées à l’aide d’un microscope Zeiss Axioscope 20, les photos prisent par une caméra DOM300, et l’image est visionnée par un logiciel Oasis (Oasis Scientific Inc).
33 Les mesures sont réalisées à l’aide du logiciel open source ImageJ version 1.48 [figure 22], un logiciel de traitement d’image crée par le National Institute of Health (NIH) (Reinking, 2007), qui permet selon (Egan et al., 2012) d’obtenir des mesures précises comparables à d’autres logiciels plus couteux.
Figure 22 : interface du logiciel ImageJ version 1.48.
Les mesures prises sont : • Mesures linéaires :
Épaisseur du cortex, et du périoste. Mesurées au niveau de la diaphyse sur deux champs microscopiques avec l’objectif (X10).
Longueur des zones de croissance, mesurées au niveau des deux épiphyses avec l’objectif (X4) [Figure 23] et qui consistent en :
- Zone de réserve (R) : ici le cartilage articulaire.
- Zone proliférative (P) : les chondrocytes sont organisés en colonne.
- Zone hypertrophique (H) : chondrocytes hypertrophiées sans matrices calcifiée.
34
Figure 23 : les zones de croissance au niveau du cartilage de conjugaison.
• Mesures de surface exprimées en pourcentage sur deux champs microscopiques avec l’objectif (X4) :
-Surface des trabécules/ surface de l’os. -Surface de l’os médullaire/ surface de l’os -Surface du tissu adipeux/ surface de l’os.
-Surface du tissu osseux/ surface du cortex = Densité osseuse du cortex (%) : au niveau de la diaphyse avec l’objectif (X10)
• Mesures cellulaire :
Le comptage cellulaire lui s’est fait manuellement. Les cellules dénombrées sont les ostéoclastes, facile à reconnaitre et indicateur de l’activité métabolique de l’os. Le comptage s’est fait pour la diaphyse au niveau de l’endoste [figure 24-1] et pour les épiphyses au niveau de la spongiosa primaire [figure 24-2]. Les cellules sont dénombrées sur dix champs macroscopiques avec un objectif (X40).
Dans les trois parties de l’os [diaphyse/ épiphyses].
R
P
H
35
Figure 24 : schéma suivi pour le comptage cellulaire : 1-au niveau de la diaphyse, 2-au niveau des épiphyses.
Toutes les mesures en détail sont mises en annexe :
- Annexe 3 : pour les mesures métrique et pondérale.
- Annexe 4 : pour les mesures histologiques effectuées au niveau de la diaphyse. - Annexe 5 : pour les mesures histologiques effectuées au niveau des épiphyses.
5.3.Méthodes statistiques :
Les statistiques réalisées sont la moyenne, les écarts types en plus du coefficient de corrélation. Le test student fut utilisé pour déterminer si les mesures étaient statistiquement différentes, la différence était considérée significative à P<0,05. Toutes les statistiques étaient générées en utilisant le logiciel Microsoft Excel 2007.
Chapitre III :
36
CHAPITREII : RESULTATS ET DISCUSSION
1. Mesures métriques et pondérales : 1.1.Poulets de chair :
Les résultats des mesures métriques et pondérales sont représentés dans le tableau ci-après (moyenne ± écart type) :
• Poids relatif = poids fémur X100/ poids vif.
• Pourcentage cendre = poids cendre X100/poids sec.
Tableau 3 : Mesures métriques et pondérales chez le poulet de chair.
Paramètre /lot C CC
Poids vif des animaux (g) 395± 69,37* 2320± 458,05*
Poids net fémur (g) 0,96±0,33* 7,71±2,47*
Poids relatif fémur(%) 0,24±0,05* 0,33±0,06*
Longueur fémur (mm) 37,4± 2,33* 72± 5,96*
Poids net fémur/ longueur fémur (g/mm) 0,02± 0,01* 0,1± 0,02*
Poids sec fémur (g) 0,53±0,14* 4,24±1,09*
Poids des cendres de fémur (g) 0,25±0,06* 1,97±0,44* Pourcentage cendre (%) 47,04±4,34 46,69±1,87
C : animaux de 28 jours ; CC : animaux de 56 jours.
*: les valeurs dans une seule ligne sont significativement différentes (P< 0,05).
1.1.1. Poids vif des animaux:
L’âge des animaux lors de la mise sur le marché est assez tardif en Algérie, ce que note aussi Mouhous et al., (2012) et Meziane et al., (2012) dont les deux études ont été effectuées ici en Algérie.
Chez nos animaux le poids vif à 56 jours est six fois celui trouvé à 28 jours avec une variabilité entre individus assez élevée démontrée par les valeurs élevées de l’écart type ; mais la moyenne en fin de cycle reste légèrement faible par rapport aux deux auteurs précédents qui ont trouvé un poids vif de 2,76 Kg et 2,5 Kg respectivement.
Ces mêmes auteurs jugent les performances enregistrées comme étant loin derrière celles des pays européen et américain, et cela serait dû à la mauvaise maitrise dans notre pays de