HAL Id: hal-02123093
https://hal.archives-ouvertes.fr/hal-02123093v4
Submitted on 25 May 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
Sur la taille finale des épidémies dans un environnement
périodique
Nicolas Bacaër
To cite this version:
Nicolas Bacaër. Sur la taille finale des épidémies dans un environnement périodique. Comptes Rendus
Biologies, Elsevier Masson, 2019, 342, pp.119 - 123. �10.1016/j.crvi.2019.07.001�. �hal-02123093v4�
Sur la taille finale des ´epid´emies dans un
environnement p´eriodique
Nicolas Baca¨er
∗Comptes Rendus Biologies 342 (2019) 119-123
doi:10.1016/j.crvi.2019.07.001
Traductions : [ar, de, es, it, ja, nl, pt, ru, zh], [html]
R´esum´e
Il est difficile de pr´edire la taille finale d’une ´epid´emie comme celle de dengue qui s´evit actuellement `a La R´eunion, d’autant qu’il faut tenir compte de la sai-sonnalit´e. Dans cet article, on se propose d’´etudier d’un point de vue th´eorique comment, dans le cadre d’un mod`ele tr`es simple de type S-I-R pour une maladie `a transmission directe, un environnement saisonnier p´eriodique peut modifier la taille finale d’une ´epid´emie. On obtient des r´esultats analytiques en supposant que l’amplitude de la saisonnalit´e est faible.
1
Introduction
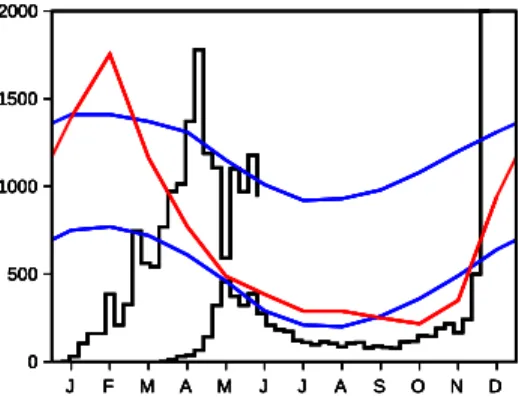
Une ´epid´emie de dengue s´evit actuellement sur l’ˆıle de La R´eunion, dans l’oc´ean Indien. Plus de 5 000 cas ont ´et´e confirm´es biologiquement entre jan-vier et juin 2018, et plus de 16 000 cas ´evocateurs de la dengue ont ´et´e si-gnal´es (figure 1). Fin juin, du fait de l’arriv´ee de l’hiver austral mais aussi de la lutte antivectorielle, l’´epid´emie recule. Cependant il est difficile de pr´evoir si le nombre d’infections diminuera suffisamment pour empˆecher une seconde vague ´epid´emique `a la fin 2018, lorsque les conditions climatiques seront plus favorables aux moustiques vecteurs de la maladie. C’est cette deuxi`eme vague qui en 2006 avait infect´e presque un tiers de la population avec le chikungunya. Il serait tentant de mod´eliser math´ematiquement la propagation de la dengue de mani`ere r´ealiste tout en limitant la complexit´e du mod`ele pour n’avoir que quelques param`etres inconnus, comme on a essay´e de le faire pour le chikun-gunya dans (Baca¨er, 2007). Mais les incertitudes qui p`esent sur ces param`etres et sur leur d´ependance par rapport aux variables climatiques sont si grandes
∗Institut de Recherche pour le D´eveloppement, Les Cordeliers, Paris, France,
J F M A M J J A S O N D 0 500 1000 1500 2000 J F M A M J J A S O N D 0 500 1000 1500 2000 J F M A M J J A S O N D 0 500 1000 1500 2000 J F M A M J J A S O N D 0 500 1000 1500 2000 J F M A M J J A S O N D 0 500 1000 1500 2000
Figure1 – Courbe en noir du haut (´echelle sur l’axe vertical de gauche) : esti-mation du nombre de cas hebdomadaires ´evocateurs de la dengue `a La R´eunion entre janvier et juin 2018, d’apr`es Sant´e publique France (2018). Courbe en noir du bas (mˆeme ´echelle) : cas hebdomadaires de chikungunya en 2005, avant le grand pic de la fin 2005 et du d´ebut de 2006 (Baca¨er, 2007). Donn´ees clima-tiques `a la station de l’a´eroport de La R´eunion, d’apr`es M´et´eo France : en bleu, temp´erature minimale (de 18,0 `a 23,7oC) et temp´erature maximale (de 25,2 `a
30,1oC) ; en rouge, pr´ecipitations mensuelles (de 43 `a 351 mm).
que les r´esultats num´eriques risquent fort d’ˆetre assez douteux (Raoult, 2016). On se contentera ci-dessous d’un mod`ele ´epid´emique extrˆemement simplifi´e `a transmission directe et non vectorielle, qui ne pr´etend donc pas pouvoir ˆetre appliqu´e au cas de la dengue `a La R´eunion. Cela n’aurait pas vraiment de sens de vouloir ajuster ses param`etres aux donn´ees ´epid´emiques. L’objectif est plutˆot d’attirer l’attention sur la question de la taille finale d’une ´epid´emie dans un environnement saisonnier p´eriodique, qui a ´et´e peu ´etudi´ee d’un point de vue th´eorique.
Mˆeme les mod`eles math´ematiques les plus simples qui tiennent compte de la saisonnalit´e pr´esentent de nombreuses difficult´es. On sait depuis longtemps que pour les mod`eles de maladies end´emiques, un coefficient p´eriodique peut engen-drer des oscillations avec une p´eriode diff´erente, voire des oscillations chaotiques (Choisy et Cazelles, 2009). Dans l’´etude (Baca¨er, 2007) inspir´ee de l’´epid´emie de chikungunya `a La R´eunion, on a vu par ailleurs que mˆeme la notion clas-sique de reproductivit´e, not´ee R0 `a la suite de Lotka (1939), dans les mod`eles
p´eriodiques devait ˆetre d´efinie avec quelques pr´ecautions pour que l’in´egalit´e R0 > 1 traduise bien l’instabilit´e de la situation sans ´epid´emie ; la difficult´e
apparaˆıt notamment pour les mod`eles avec au moins deux compartiments d’in-fect´es.
Dans (Baca¨er et Gomes, 2009), on a abord´e le mod`ele S-I-R p´eriodique, simple g´en´eralisation du mod`ele classique de Kermack et McKendrick (Hillion,
1986, p. 75) : dS dt = −a(t) S I/N, dI dt = a(t) S I/N − b I, dR dt = b I, (1) o`u N est la taille de la population, S(t) le nombre de personnes susceptibles d’ˆetre infect´ees, I(t) le nombre de personnes infect´ees, R(t) le nombre de per-sonnes gu´eries, a(t) le taux de contact infectieux, b le taux de gu´erison. Les conditions initiales sont S(t0) = s = N − i avec 0 < i < N , I(t0) = i et
R(t0) = 0. La fonction a(t) est p´eriodique de p´eriode T . Apr`es avoir pos´e
R0= 1 bT Z T 0 a(t) dt,
on a montr´e que la propri´et´e de seuil ´epid´emique se traduisait de la mani`ere suivante : si R0 < 1, alors la taille finale R(∞) de l’´epid´emie tend vers 0 si i
tend vers 0 ; si au contraire R0> 1, alors
R(∞) > N (1 − 1/R0),
quel que soit 0 < i < N . Cette propri´et´e s’´etend d’ailleurs des mod`eles avec une transmission directe comme le syst`eme (1) aux mod`eles avec une transmission vectorielle comme pour la dengue. Dans (Baca¨er et Ait Dads, 2011, figure 1), on a ´etudi´e num´eriquement comment la taille finale R(∞) de l’´epid´emie d´epend de l’instant initial t0 et de l’amplitude du taux de contact infectieux a(t). On
a observ´e que R(∞) pouvait varier du simple au double pour une mˆeme valeur de la reproductivit´e R0.
En combinant m´ethodes num´eriques et m´ethodes analytiques, on ´etudie ici plus attentivement la taille finale de l’´epid´emie dans le cas particulier o`u
a(t) = ¯a(1 + ε φ(t))
avec une fonction φ(t) (disons continue par morceaux) qui est p´eriodique de p´eriode T et de moyenne nulle, et avec un param`etre ε qui est petit de sorte que l’environnement n’est que faiblement saisonnier. Notons Sε(t), Iε(t) et Rε(t) les
solutions correspondantes mais avec toujours la mˆeme condition initiale (N − i, i, 0). On montre dans la section 2 que
Rε(∞) = R0(∞) + N c ε + o(ε)
quand ε → 0, o`u R0(∞) est la taille finale de l’´epid´emie dans un environnement
constant. Le coefficient correcteur c peut ˆetre positif ou n´egatif.
Si φ(t) = cos(ωt), si le nombre de personnes infect´ees au d´epart est petit devant la taille de la population (i ≪ N ), si R0 = ¯a/b > 1 avec R0 proche
de 1, et si i/N ≪ (R0−1)2, alors on d´etermine analytiquement le coefficient
correcteur c en fonction des param`etres N , i, ¯a, b, t0et ω du mod`ele. On trouve
que l’environnement p´eriodique augmente la taille finale de l’´epid´emie (c > 0) si le taux de contact infectieux v´erifie a(t0+ τ ) > ¯a, o`u t0+ τ est le moment o`u
2
Formule exacte pour le coefficient correcteur
Rappelons tout d’abord qu’`a partir du syst`eme diff´erentiel (1), on montre que : Sε(t) + Iε(t) + Rε(t) = N pour tout t ≥ t0; Sε(t) > 0, Iε(t) > 0 et
Rε(t) ≥ 0 pour tout t ≥ t0; Sε(t) est une fonction d´ecroissante qui tend vers
une limite Sε(∞) ; Rε(t) est une fonction croissante qui tend vers une limite
Rε(∞) ; Iε(t) tend vers une limite qui ne peut ˆetre que 0 (Baca¨er et Gomes,
2009). Int´egrons la troisi`eme ´equation diff´erentielle de t0 `a l’infini : cela donne
Rε(∞) = bR ∞
t0 Iε(t) dt. Par ailleurs, la premi`ere ´equation diff´erentielle s’´ecrit
1 Sε
dSε
dt = −a(t)Iε(t)/N . Int´egrons de mˆeme pour obtenir
logSε(∞) N − i = − ¯ a N Z ∞ t0 Iε(t) dt − ¯ a N ε Z ∞ t0 Iε(t) φ(t) dt .
On peut remplacer la premi`ere int´egrale et tenir compte de ce que Sε(∞) =
N − Rε(∞) pour obtenir l’´equation implicite
logN − Rε(∞) N − i + ¯ a b Rε(∞) N + ¯ a N ε Z ∞ t0 Iε(t) φ(t) dt = 0 , (2)
qui est de la forme F (Rε(∞), ε) = 0. Lorsque ε = 0, le dernier terme `a droite
disparaˆıt et on reconnaˆıt l’´equation classique pour la taille finale de l’´epid´emie dans un environnement constant (Hillion, 1986, p. 76). D’apr`es le th´eor`eme des fonctions implicites (Deschamps et Warusfel, 2001, p. 892), la fonction qui `a ε associe Rε(∞) est continˆument d´erivable au voisinage de ε = 0, de sorte que
Rε(∞) = R0(∞) + N c ε + o(ε) pour ǫ → 0, avec
N c = −∂F ∂ε(R0(∞), 0) / ∂F ∂R(R0(∞), 0) . On trouve ainsi c = Na/N¯ N −R0(∞) − ¯ a b Z ∞ t0 I0(t) φ(t) dt . (3)
Rappelons comme dans l’introduction que R0(∞) > N (1−b/¯a). Donc le d´enominateur
de la formule (3) est strictement positif et en particulier non nul. Le coefficient c a le mˆeme signe que l’int´egraleR∞
t0 I0(t) φ(t) dt.
On peut ´evaluer num´eriquement cette int´egrale. On utilise d’abord un logiciel de calcul num´erique (tel que Scilab et sa fonction≪ode≫, voir www.scilab.org/fr)
pour r´esoudre le syst`eme diff´erentiel (1) avec ε = 0. On obtient ainsi (S0(t), I0(t), R0(t))
pour un ensemble discret de valeurs de t, avec un petit pas de temps. On en d´eduit en particulier la valeur R0(∞). Puis on calcule l’int´egrale avec cette mˆeme
discr´etisation du temps. On en d´eduit la valeur du coefficient c.
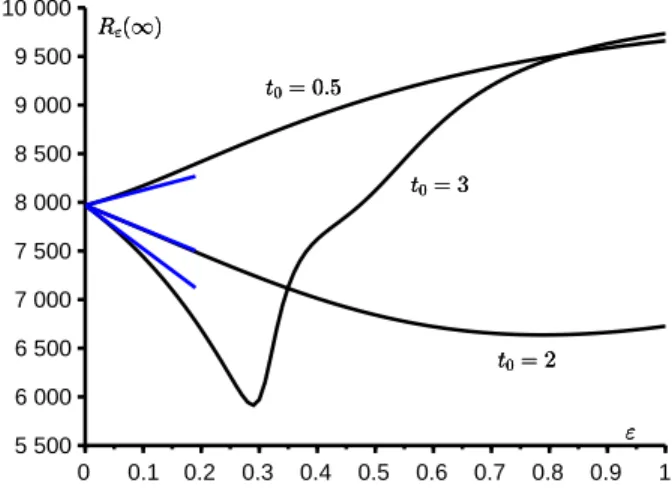
Prenons un exemple. Supposons que φ(t) = cos(ωt) avec ω = 2π/T , N = 10 000, i = 1, T = 12 mois, ¯a = 10/mois et b = 5/mois. Alors la reproductivit´e est R0 = 2 ; la taille finale de l’´epid´emie dans un environnement constant est
R0(∞) ≃ 7 968. La figure 2 montre comment Rε(∞) varie en fonction de ε
moments diff´erents d’introduction du premier cas infect´e dans la population : 0,5 mois, 2 mois ou 3 mois. La figure montre aussi l’approximation R0(∞) + N c ε
pour ε petit, avec le coefficient correcteur c calcul´e selon la formule (3). On observe que le coefficient c peut ˆetre positif ou n´egatif, de sorte que la taille finale de l’´epid´emie peut ˆetre plus grande ou plus petite que dans un environnement constant. On observe aussi que pour t0 = 3 mois, la fonction Rε(∞) varie en
fonction de ε de mani`ere plus compliqu´ee que pour les deux autres valeurs de t0 : en particulier, cette fonction n’est d´ecroissante que tant que ε < 0,3.
0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1 10 000 6 000 8 000 7 000 9 000 5 500 6 500 7 500 8 500 9 500
Figure 2 – La taille finale de l’´epid´emie en fonction de ε pour trois valeurs diff´erentes de t0. En bleu : l’approximation R0(∞) + N c ε.
3
Formules approch´
ees
Kermack et McKendrick ont trouv´e — voir par exemple (Baca¨er, 2012, §2) ou (Gani et Badrikian, 1975, p. 235) — une approximation pour I0(t) `a partir
de l’´equation exacte dR0/dt = b(N − s e−¯aR0/(N b)−R0) v´erifi´ee par R0(t). Un
d´eveloppement limit´e de l’exponentielle e−x= 1 − x + x2/2 + o(x2) tronqu´e `a
l’ordre 2 conduit `a une ´equation de Ricatti r´esoluble pour R0(t) et `a une courbe
en cloche sym´etrique pour I0(t),
I0(t) ≃ N X ch2(Y (t − t0) − Z) pour tout t > t0, o`u X = b 2N W2 2 ¯a2s , Y = b W 2 , th(Z) = ¯ a s b N −1 W ,
et W = ( [a s¯ b N −1] 2+ 2 s i N2 ¯ a2 b2 ) 1/2.
Ici, ch(·) et th(·) d´esignent le cosinus et la tangente hyperboliques. Rappe-lons que s = N − i. L’approximation pour I0(t) est d’autant meilleure que
¯
aR0(∞)/(N b), la valeur maximale du terme dans l’exponentielle, est petit
de-vant 1. Alors c ≃ N¯aX N −R0(∞)− ¯ a b Z ∞ t0 φ(t) ch2(Y (t − t0) − Z) dt . (4)
Supposons plus pr´ecis´ement que R0 = ¯a/b → 1 avec R0 > 1 et que
(i/N )/(R0−1)2→0 ; autrement dit, R0est proche de 1 et i/N ≪ (R0−1)2.
Alors i → 0 et on a bien R0(∞) → 0 d’apr`es l’´equation (2) avec ε = 0. Comme
on a aussi i/N ≪ (R0−1), on trouve que
W ∼ ¯a b −1, X ∼ (1 − b/¯a)2 2 , Y ∼ ¯ a − b 2 et 1 − th(Z) ∼ (b/¯i/Na−1)2. Donc Z → +∞ et 1 − th(Z) ∼ 2 e
−2Z , ce qui donne Z ∼ 1 2log 2(b/¯a − 1)2 i/N .
Posons τ = Z/Y ; c’est en t = t0+ τ que l’approximation de I0(t) atteint son
pic. Notons que Y → 0 et que τ → +∞. Apr`es le changement de variable t = t0+ τ + u, on remarque que la fonction 1/ch2(Y u) est presque nulle en
dehors du voisinage de u = 0, de sorte que Z ∞ −τ φ(t0+ τ + u) ch2(Y u) du ≃ Z ∞ −∞ φ(t0+ τ + u) ch2(Y u) du .
Cette derni`ere int´egrale se calcule explicitement lorsque φ(t) = cos(ωt). En effet, Z ∞ −∞ cos(ω(t0+ τ + u)) ch2(Y u) du = cos(ω(t0+ τ )) Z ∞ −∞ cos(ωu) ch2(Y u)du
puisque l’int´egrale avec la fonction impaire sin(ωu) s’annule. D’apr`es la formule dans l’appendice de (Baca¨er, 2015), on a
Z ∞ −∞ cos(ωu) ch2(Y u)du = πω Y2 sh(πω2Y),
o`u sh(·) d´esigne le sinus hyperbolique. Avec les ´equivalents de X et Y d´ej`a obtenus, on arrive finalement `a
c ≃ cos(ω(tN 0+ τ )) N −R0(∞)− ¯ a b 2πω ¯ a sh(πω ¯ a−b) . (5)
On voit que le signe de c est le mˆeme que celui de cos(ω(t0+ τ )). Donc
l’envi-ronnement p´eriodique augmente la taille finale de l’´epid´emie si a(t0+ τ ) > ¯a, o`u
t0+ τ est le moment o`u l’´epid´emie atteindrait son pic dans un environnement
constant.
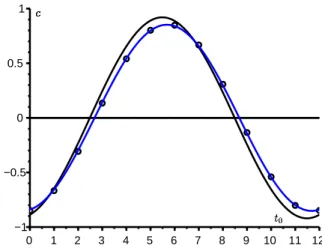
Comme exemple, prenons les mˆemes valeurs des param`etres que dans la figure 2 sauf que ¯a = 6/mois pour avoir R0 = 1,2 plus proche de 1 ; [8,
p. 240] indique d’ailleurs que l’approximation en cloche sym´etrique de Kermack et McKendrick n’est satisfaisante que pour R0< 1,5. La figure 3 compare
l’ex-pression exacte (3) du coefficient correcteur c avec les approximations (4) et (5). Avec nos valeurs num´eriques, ces deux derni`eres approximations sont indiscer-nables. Elles sont d’autant plus proches de la valeur exacte que R0 est proche
de 1. Notons qu’ici i/N = 10−4
≪(R0−1)2= 0,04. La figure s’interpr`ete comme
ceci : si l’´epid´emie d´emarrait `a t0= 0, les valeurs des param`etres conduiraient
`
a un pic de l’´epid´emie environ τ ≃ 6,3 mois plus tard dans un environnement constant ; or l’environnement p´eriodique sera d´efavorable `a ce moment (on sera dans le creux du facteur cos ωt) ; donc la taille finale de l’´epid´emie sera moindre et c < 0. 0 1 2 3 4 5 6 7 8 9 10 11 12 0 −1 1 −0.5 0.5
Figure 3 – Le coefficient correcteur c pour la taille finale de l’´epid´emie en fonction de t0, l’instant du d´ebut de l’´epid´emie. Comparaison de l’expression
exacte (3) [en noir] avec les approximations (4) [points] et (5) [en bleu].
4
Remarque
On peut adapter la formule exacte (3) pour le coefficient correcteur c `a des mod`eles plus complexes. Supposons par exemple que l’´epid´emie se propage entre deux populations (comme des hommes et des vecteurs) suivant le sch´ema S-I-R,
avec N1= S1(t) + I1(t) + R1(t) et N2= S2(t) + I2(t) + R2(t) : dS1 dt = −a(t)S1 I2 N2, dI1 dt = a(t)S1 I2 N2 −b1I1, dR1 dt = b1I1, dS2 dt = −a(t)I1 S2 N2 , dI2 dt = a(t)I1 S2 N2 −b2I2, dR2 dt = b2I2.
Supposons aussi que a(t) = ¯a(1 + ε φ(t)) comme pr´ec´edemment. Notons S1,ε(t),
I1,ε(t), etc., les solutions. Posons i1= I1(t0) et i2= I2(t0). On trouve facilement
l’´equivalent de l’´equation (2), qui est un syst`eme :
logN1−R1,ε(∞) N1−i1 + ¯ a b2 R2,ε(∞) N2 + ¯ a N2ε Z ∞ t0 I2,ε(t) φ(t) dt = 0 logN2−R2,ε(∞) N2−i2 + ¯a b1 R1,ε(∞) N2 + a¯ N2 ε Z ∞ t0 I1,ε(t) φ(t) dt = 0.
Ce syst`eme a des solutions de la forme Rk,ε(∞) = Rk,0(∞) + Nkckε + o(ε) pour
k = 1 ou 2. Avec le th´eor`eme des fonctions implicites, on trouve que N1c1 N2c2 = 1 N1−R1,0(∞) − ¯ a b2N2 −b¯a 1N2 1 N2−R2,0(∞) −1 ¯ a N2 R∞ t0 I2,0(t) φ(t) dt ¯ a N2 R∞ t0 I1,0(t) φ(t) dt .
Il n’est cependant pas possible de poursuivre comme dans la section 3 car on ne dispose pas de formules approch´ees explicites pour I1,0(t) et I2,0(t).
5
Conclusion
On a d´etermin´e de mani`ere analytique comment la taille finale de l’´epid´emie ´etait influenc´ee par un taux de contact infectieux p´eriodique de faible ampli-tude, lorsque la reproductivit´e R0reste proche de 1. Il s’agit d’un petit progr`es
par rapport aux ´etudes pr´ec´edentes qui ´etaient soient qualitatives comme dans (Baca¨er et Gomes, 2009), soit purement num´eriques comme dans (Baca¨er et Ait Dads, 2011, figure 1). Les hypoth`eses sont n´eanmoins assez restrictives. Elles ´evitent en particulier les cas o`u plusieurs pics ´epid´emiques se produisent, puisque l’on reste proche de la situation `a un seul pic des environnements constants. Or ce sont justement ces cas qui risquent de se pr´esenter avec l’´epid´emie de dengue `a La R´eunion.
R´
ef´
erences
[1] N. Baca¨er (2007), Approximation de la reproductivit´e nette R0 pour
les maladies `a vecteurs avec une population p´eriodique de vecteurs, https://hal.archives-ouvertes.fr/hal-01291211.
[2] N. Baca¨er, M.G.M. Gomes (2009), Sur la taille finale des ´epid´emies avec saisonnalit´e, https://hal.archives-ouvertes.fr/hal-01299608. [3] N. Baca¨er, E. H. Ait Dads (2011), Les g´en´ealogies avec
sai-sonnalit´e, la reproductivit´e nette et la pand´emie de grippe, https://hal.archives-ouvertes.fr/hal-01274025.
[4] N. Baca¨er (2012), Le mod`ele de Kermack et McKendrick pour la peste `a Bombay et la reproductivit´e nette d’un type avec saisonnalit´e, https://hal.archives-ouvertes.fr/hal-01340008.
[5] N. Baca¨er (2015), Sur le mod`ele stochastique SIS pour une ´epid´emie dans un environnement p´eriodique, https://hal.archives-ouvertes.fr/hal-01265563.
[6] M. Choisy, B. Cazelles (2009), Cons´equences des dynamiques ´epid´emiques en sant´e publique : rˆole des mod`eles math´ematiques, in : J.-F. Gu´egan, M. Choisy (´ed.), Introduction `a l’´epid´emiologie int´egrative des maladies infectieuses et parasitaires, De Boeck, Bruxelles, p. 3-39.
[7] C. Deschamps, A. Warusfel (2001), Math´ematiques 2eann´ee, Dunod, Paris.
[8] J. Gani, J. Badrikian (1975), Processus stochastiques de population, in : P.-L. Hennequin (´ed.), ´Ecole d’´et´e de probabilit´es de Saint-Flour IV-1974, Springer, Berlin, p. 188-293.
[9] A. Hillion (1986), Les th´eories math´ematiques des populations, Presses Uni-versitaires de France, Paris.
[10] A.J. Lotka (1939), Th´eorie analytique des associations biologiques, 2e
par-tie, Hermann, Paris.
[11] M´et´eo France, Donn´ees climatiques de la station de Gillot-a´eroport, www.meteofrance.re/climat/reunion/gillot-aeroport/97418110/normales. [12] D. Raoult (2016), Les mod`eles pr´edictifs sont des proph´eties modernes, in :
D. Raoult, Arrˆetons d’avoir peur, Michel Lafon, Neuilly-sur-Seine.
[13] Sant´e publique France (2018), ´Epid´emie de dengue `a la R´eunion – Point ´epid´emiologique au 19 juin 2018, www.santepubliquefrance.fr.