Paternal age and reproduction
Texte intégral
Figure
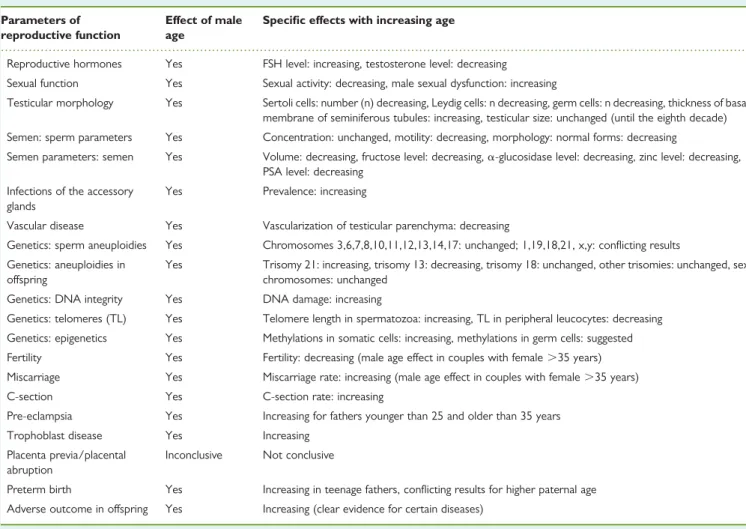
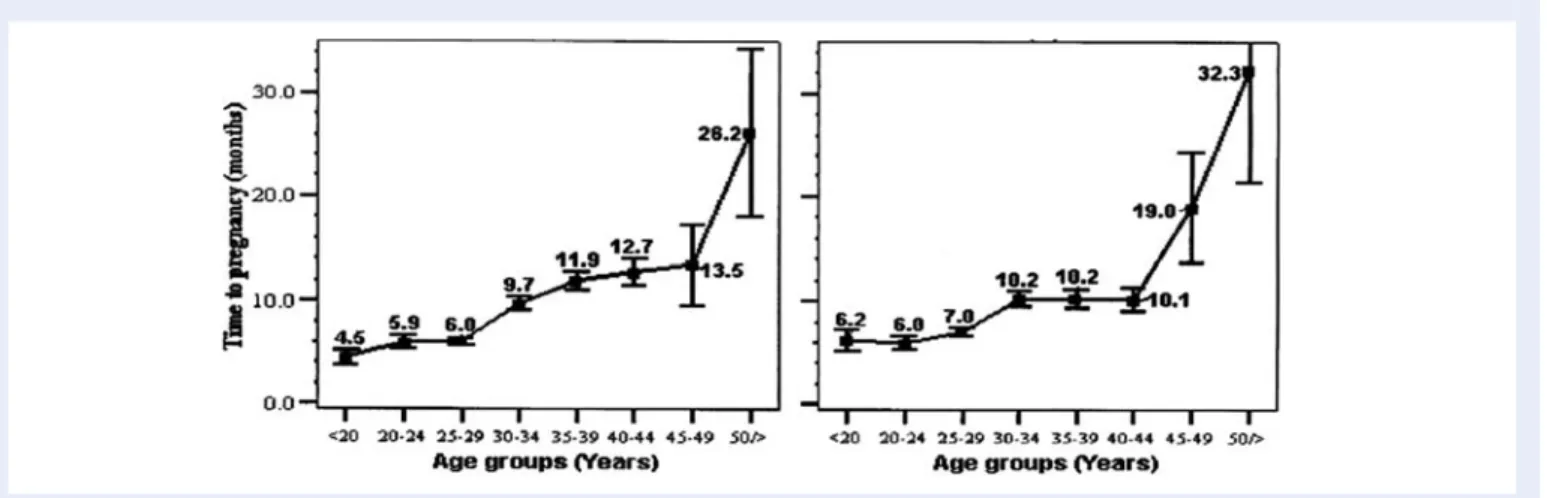
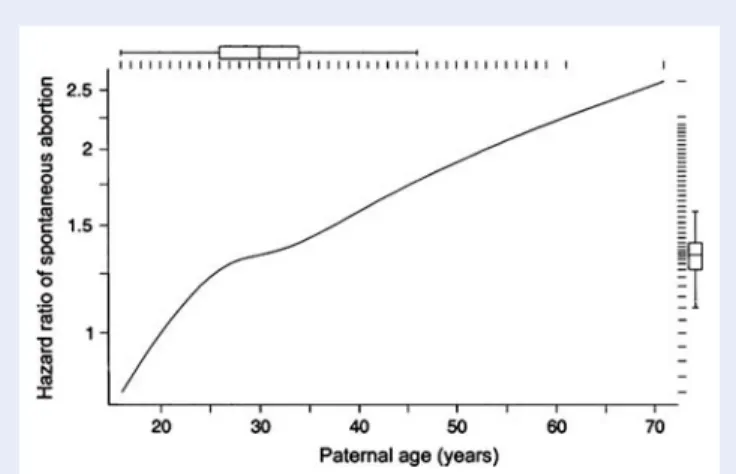
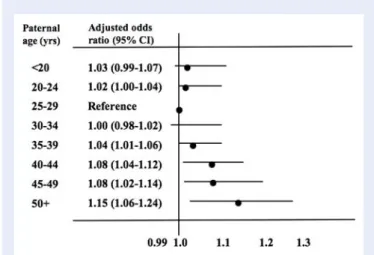
Documents relatifs
In this study, the occurrence and degree of carbon isotope fractionation during microbial dechlorination of PCE to ethene is evaluated in a laboratory experiment (microcosm) and at
As already published before [14], compared to day sleep, night sleep was significantly longer and more efficient at both M0, but especially at M6 at which time there was also
De novo apparently balanced translocations in man are predominantly paternal in origin and associated with a significant increase in paternal age... De novo apparently
We examined the association between perceived lifetime work- place stress and prostate cancer (PCa) risk in a large case–control study.. Methods: Cases were 1,933 men, aged ≤
Mitophagy, a mitochondrial quality control mechanism enabling the degradation of damaged and superfluous mitochondria, prevents such detrimental effects and reinstates
For each nymph and adult data set, hemocyte concentration and total-PO ac- tivity were analyzed using two LMMs, in which either the age of the nymphs (second, third and fourth
De nombreuses études ont confirmé l‟existence d‟une composante génétique à l‟étiopathogénie de la schizophrénie (pour revue, [22]). Cette maladie reste
The aim of the current study was to examine whether paternal age and/or delayed fatherhood (paternity age) predict self-reported positive, negative and/or depressive
