HAL Id: hal-02663261
https://hal.inrae.fr/hal-02663261
Submitted on 31 May 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
European and East-Asian Wheat Collections Using SSR
Markers
François Balfourier, Lan-Fen Wang, Florence Exbrayat-Vinson, Chenyang
Hao, Yu-She Dong, Hong-Mei Ge, Xue-Yong Zhang
To cite this version:
François Balfourier, Lan-Fen Wang, Florence Exbrayat-Vinson, Chenyang Hao, Yu-She Dong, et al..
Comparison of Genetic Diversity Level Between European and East-Asian Wheat Collections Using
SSR Markers. Scientia Agricultura Sinica, 2007, 40 (12), pp.2667-2678. �hal-02663261�
Scientia Agricultura Sinica
收稿日期:2007-06-27;接受日期:2007-08-20 基金项目:中法小麦合作计划,国家重点基础研究项目(2004CB117202) 作者简介:王兰芬(1968-),女,山东威海人,副研究员,硕士,研究方向为小麦遗传多样性与核心种质。通讯作者张学勇(1962-),男,甘肃临洮 人,研究员,博士,研究方向为小麦遗传多样性、远缘杂交与基因组进化、基因克隆等方面。Tel:010-62135294;E-mail:xueyongz@ public.bta.net.cn欧洲与东亚小麦品种遗传多样性的比较分析
王兰芬
1,BALFOURIER F
2,郝晨阳
1,EXBRAYAT-VINSON F
2,董玉琛
1,盖红梅
1,张学勇
1 (1中国农业科学院作物科学研究所/农业部作物品种资源与生物技术重点实验室/国家农作物基因资源与基因改良重大科学工程,北京 100081,中国; 2INRA, UMR Amélioration et Santé des Plantes, 234 avenue du Brézet, 63100 Clermont-Ferrand, France)摘要:
【目的】在分子水平上回答欧洲和东亚小麦品种的遗传关系和多样性差异;同时对 Genomic-SSR(gSSR)
和 EST-SSR(eSSR)多态性水平进行比较分析。
【方法】利用 38 个 Genomic-SSR 引物对和 44 对 EST-SSR 引物对分
析 371 份欧洲小麦品种和 363 份东亚品种。
【结果】共检测到 865 个等位变异,每个引物对的等位变异数为 1~50,
平均为 10.42;多态性信息含量(
PIC
)为 0~0.91, 平均为 0.53;欧洲和东亚品种分别检测到 730、716 个等位
变异,特有等位变异分别为 150、135,平均遗传丰富度分别为 8.80 和 8.61,遗传多样性指数分别为 0.46 和 0.52。
欧洲和东亚小麦品种在聚类图上明显地划分为两大类群;每个国家或大区聚类结果与其地理分布基本一致,即相
邻国家或地区的品种亲缘关系更近一些。近一半基因座的等位变频率及其分布在欧洲与东亚材料间存在明显差异。
通过标记/性状关联分析,在 4B、5A、6A、7B 染色体上发现 6 个影响穗粒数、千粒重、株高、抽穗期、有效分蘖
等重要农艺性状的基因座,其中个别基因座优势等位变异差异可能与东亚、欧洲品种的分化密切相关。中国 20 世
纪 50~80 年代育成品种在大的聚类上明显靠近欧洲材料、而远离中国 50 年代以前的育成品种和地方品种,这与
中国的育种实际相吻合。基于 Genomic-SSR 和基于 EST-SSR 的聚类图整体趋势是一致的,但由前者估算的遗传距
离远高于后者,因此,一般的 SSR 较功能基因 SSR 更易发生变异,由育种选择所引起的分化更快。
【结论】在中国
今后的小麦育种中,需要通过杂交、回交,对欧洲品种的一些重要基因座等位变异(基因)进行置换,方有可能
实现“洋为中用”的目的。
关键词:小麦;遗传多样性;gSSR;eSSR;地理分化;关联分析;
Comparison of Genetic Diversity Level Between European and
East-Asian Wheat Collections Using SSR Markers
WANG Lan-fen
1, BALFOURIER F
2, HAO Chen-yang
1, EXBRAYAT-VINSON F
2, DONG Yu-shen
1,
GE Hong-mei
1, ZHANG Xue-yong
1(
1Key Laboratory of Crop Germplasm and Biotechnology, Ministry of Agriculture, Institute of Crop Germplasm Resources, Chinese
Academy of Agricultural Sciences, Beijing 100081, China;
2INRA, UMR Amélioration et Santé des Plantes, 234 avenue du Brézet,
63100 Clermont-Ferrand, France)
Abstract
:
【Objective】This research work was performed to reveal the genetic relationship and diversity difference between
European and East-Asian wheat cultivars, and in the meantime to compare the level of polymorphism between Genomic-SSRs and
EST-SSRs. 【Method】 To analyze 371 European and 363 East-Asian wheat collections, 38 Genomic-SSRs and 44 EST-SSRs were
used. 【Result】A total of 865 alleles were detected, with a range from 1 to 50 and a mean of 10.42. Polymorphism information
content (PIC) ranged from 0 to 0.91 with an average of 0.53. Genetic diversity level in East-Asia was higher than that in Europe.
European and East-Asian accessions were divided into two distinct sub-groups by PCO based on 82 SSRs. The position of materials
in the dendrogram was accordant with their geographical location, i.e. the accessions from near geographical location were joining
into neighbors. Allele differentiation between the two continents at some loci such as Xgwm186, Xgwm257, Xgpw7433 and
Xgpw7443 was very significant. This may be corresponding to the natural and breeding selection, which was supported by the
marker/trait association analysis results. Chinese varieties released from 1950 to 1980 were clustered into one big group with
Europeans, and far away from native landraces and varieties released before 1950. This is consistent with the records of wheat
breeding history of China. Diversity revealed by Genomic-SSRs was much higher than that by EST-SSRs. However, the cluster trend
based on the two types of SSR was consistent. Therefore, differentiation of Genomic-SSR was much faster than functional genes
SSR in breeding selection. 【Conclusion】European germplasms can be efficiently used in Chinese wheat breeding program through
backcross breeding method.
Key words: Wheat; Genetic diversity; gSSR; eSSR; Geographic differentiation; Marker/trait association
0 引言
【研究意义】小麦是世界上最重要的粮食作物之
一。欧洲小麦品种在中国育种和生产中曾起过重大作
用,原产于意大利的南大
2419(Mentana)、矮立多
(
Ardito)、中农 28(Villa Glori)、阿夫(Funo)、阿
勃(
Abondanza)、郑引 1 号(St1472/506)及原产于
罗马尼亚的洛夫林
10 和洛夫林 13(Lovrin 10 and 13)
都曾在中国大面积推广;在明确的
16 个骨干亲本中,
就有
4 个来自欧洲
[1]。对中国
1 680 份育成品种(系)
进行遗传多样性分析发现,半个多世纪的育种已导致
小麦品种遗传基础日趋狭窄,大大降低了对逆境和病
虫害流行的抵御和缓冲能力
[2]。因此,进一步引进和
利用国外材料,拓宽品种的遗传基础势在必行。
【前人
研究进展】2004 年,中法两国通过政府间合作,对等
交换了
384 份小麦种质资源,其中绝大部分为育成品
种。董玉琛等
[3]在对这批材料的农艺性状鉴定与评价
后指出:欧洲小麦,不论新品种还是老品种都有很大
的潜在利用价值,特别是在丰产性、抗病性和品质方
面都有可取之处。应加大引种力度,对其进行充分研
究和利用。
【本研究切入点】欧洲,尤其是欧洲西半部
为多云阴湿的海洋性气候,而东亚,特别是中国小麦
主产区以大陆性气候为主,悬殊的气候可能造成品种
间存在比较大的差异,但尚未见两大洲小麦品种的遗
传多样性互相比较分析的报道。【拟解决的关键问题】
对
371 份欧洲小麦品种和 363 份东亚品种共 82 对 SSR
引物的基因型分析,试图在分子水平上回答欧洲和东
亚 小 麦 品 种 的 遗 传 关 系 和 多 样 性 差 异 ; 同 时 对
Genomic-SSR(gSSR)和 EST-SSR(eSSR)多态性水
平进行比较分析。
1 材料与方法
1.1 材料
供试小麦材料
734 份,其中 371 份来自欧洲 18
个国家;363 份来自东亚 7 个国家,中国的材料含盖
了
23 个省份。本研究根据材料的份数和地理位置(即
地理相邻、份数较少的地区或国家间互相合并构成一
个小组)将所有的材料划分为
35 小组(表 1)。
1.2 方法
1.2.1 DNA 提取 每份材料取 1~2 株叶片,用
SIGMA 公司的试剂盒(G2N-350)提取基因组 DNA。
1.2.2
SSR 荧光标记引物的选择及检测 根据引物
的扩增效果及其在染色体上的位置,选取相对均匀分
布于
21 条染色体的 82 对 SSR 引物。其中 38 对为
genomic-SSR,除 cfd17h8 来自法国 INRA-Clermont-
Ferrand
[4]外,该引物对同时在两个基因座扩增,其余
37 对均来自 Röder 等发表的小麦 SSR 连锁图
[5];
44
对
EST-SSR (Xgpw 和 Xcfe)是 INRA-Clermont-
Ferrand 从小麦 EST 中开发的。所用引物及其在欧洲
及东亚的等位变异丰富度和多态性信息含量见表
2。
1.2.3 扩增体系
试验采用两种不同的体系,一种是
直接将荧光染料标记在引物的
5′端,扩增体系参考
Röder 等
[5];另一种是将荧光物质标记
M13 引物,引
物 合 成 时 , 在 前 链
5′ 端 增 加 M13 序 列
(5′-CACGACGTTGTAAAACGAC-3′),扩增体系参
考
Nicot 等的方法
[6]。
1.2.4 扩增产物的检测
扩增产物的电泳采用 DNA
自动分析仪
ABI Prism 3100,数据处理应用 Genescan
(
Version 3.1)和 Genotyper(Version 2.5)等软件。
当某基因座出现杂合时,笔者依据峰的强弱加以取舍。
只有当引物扩增效果很好而扩增两次以上都没检测到
扩增产物的材料,在该基因座赋予“
0”扩增。
1.2.5 遗 传 多 样 性 分 析
采 用 GENETIX
[7]、
ARLEQUIN
[8]、
NTSYS2.10
[9]、
PHYLIP
[10]等软件,对
数据进行统计分析及聚类。统计每对
SSR 引物的等位
变异丰富度(
A
ij)、多态性信息含量,同时计算不同群
体的总等位变异数、平均遗传丰富度(
∑Aij/loci)和
遗传多样性指数(
Ht)。每个基因座多态性信息含量用
PIC(polymorphism information content)表示,PIC
表 1 材料的地理分布、数量、总等位变异、稀有等位变异、平均遗传丰富度及遗传多样性指数
Table 1 Geographic distribution, total alleles, rare alleles, mean genetic richness and
genetic diversity of the 734 collections after
being grouped based on their geographical origin
地理分组 Geographic groups 代号 Code 材料数 No. of accessions 总等位变异数 Total alleles 稀有等位变异 Rare alleles ∑Ai j/loci Ht 欧洲 Europe 芬兰 Finland FIN 10 245 0 2.95 0.42 瑞典 Sweden SWE 22 308 81 3.71 0.42 荷兰 Netherlands NLD 22 278 74 3.35 0.38 英国 England GBR 22 292 75 3.52 0.39 法国 France FRA 22 279 88 3.36 0.39 比利时 Belgium BEL 22 280 73 3.37 0.39 德国 Germany DEU 22 336 106 4.05 0.45 瑞士Switzerland CHE 22 344 94 4.14 0.46 奥地利 Austria AUT 22 307 80 3.7 0.42
捷克斯洛伐克 Czech republic CZE 22 323 87 3.89 0.44
波兰 Poland POL 21 291 44 3.51 0.41 保加利亚 Bulgaria BGR 22 329 86 3.96 0.46 罗马尼亚 Romania ROM 22 312 89 3.76 0.39 塞尔维亚 Yugoslavia YUG 22 309 74 3.72 0.43 匈牙利 Hungary HUN 22 324 56 3.9 0.45 意大利 Italy ITA 22 299 75 3.6 0.4 西班牙 Spain ESP 22 378 109 4.55 0.49 葡萄牙Portugal PRT 10 266 0 3.2 0.46 总计 Total 371 730 470 8.8 0.46 东亚East Asia 北京+天津 Beijing+Tianjin BT 32 357 118 4.3 0.45 山西 Shanxi ShanX 18 325 0 3.92 0.46 河北 Hebei Hebei 20 333 0 4.01 0.48 河南 Henan Henan 29 378 110 4.55 0.49 陕西 Shanxi Shxi 20 311 0 3.75 0.45 山东 Shandong ShanD 25 343 97 4.13 0.46 安徽+湖北+浙江 Anhui+Hubei+Zhejiang AHC 20 275 0 3.31 0.4 江苏 Jiangsu Jsu 22 339 96 4.08 0.48 福建+广东 Fujian+Guangdong FG 8 222 0 2.67 0.43 四川+云南+贵州Sichuan+Yunnan+Guizhou SYG 19 275 0 3.31 0.42 黑龙江 Heilongjiang HLJ 34 355 96 4.28 0.45 吉林+辽宁+韩国 Jilin+Liaoning+Korea JLK 16 324 0 3.9 0.49 甘肃+蒙古 Gansu+Mongolia GM 27 361 125 4.35 0.44 青海+西藏Qinghai+Xizang QT 11 277 0 3.34 0.48 新疆Xinjiang SinK 10 272 0 3.28 0.48 日本Japan Japan 24 322 89 3.88 0.47 印度+巴基斯坦+ India+Parkistan+ 缅甸+克什米尔 Myanmar+Kashmir IPB 28 379 103 4.57 0.5 总计 Total 363 716 420 8.61 0.52
表 2 引物所在染色体及其在欧洲及东亚材料中的等位变
异丰富度和多态性信息含量
Table 2 Chromosomal location, total number of alleles and
polymorphic information content (PIC) for 83
microsatellite markers
等位变异丰富度Aij 多态性信息含量PIC 引物 Primers 染色体 Chromosome 欧洲 (Europe) 东亚 (East-Asia) 欧洲 (Europe) 东亚 (East-Asia) Xgpw7072 1AS 7 6 0.70 0.73 Xcfe167 1A 1 1 0.00 0.00 Xcfe189 1A 1 1 0.00 0.00 Xgpw7443 1BL 2 2 0.05 0.48 Xgpw7577 1BL 4 4 0.10 0.34 Xgpw7082 1DS 3 3 0.26 0.10 Xcfe78 1DS 1 1 0.00 0.00 Xgpw7570 2AL 6 3 0.17 0.02 Xcfe175 2A 3 3 0.37 0.48 Xgpw7438 2BS 2 2 0.50 0.50 Xcfe52 2B 8 10 0.43 0.51 Xgpw7325 2DS 1 1 0.00 0.00 Xcfe68 2D 3 3 0.23 0.03 Xgpw7213 3AL 3 3 0.03 0.39 Xgpw7335 3BL 1 1 0.00 0.00 Xgpw7452 3BS 9 5 0.64 0.43 Xgpw7586 3DL 1 1 0.00 0.00 Xgpw7553 3DS 1 1 0.00 0.00 Xcfe172 3D 3 3 0.15 0.13 Xcfe300 4A 11 9 0.70 0.55 Xgpw7241 4BL 2 2 0.01 0.02 Xcfe8 4BS 3 5 0.65 0.66 Xgpw7666 4DL 1 1 0.00 0.00 Xgpw7795 4DS 4 7 0.08 0.06 Xcfe186 5A 3 3 0.58 0.42 Xgpw7218 5AS 4 6 0.54 0.56 Xgwm7574 5BL 1 1 0.00 0.00 Xgpw7425 5BL 4 2 0.11 0.19 Xgpw7107 5DL 1 1 0.00 0.00 Xcfe301 5D 6 5 0.58 0.60 Xgpw7384 6AL 4 4 0.24 0.43 Xgpw7592 6AS 2 2 0.17 0.43 Xcfe273 6AS 2 2 0.18 0.43 Xcfe214 6B 1 1 0.00 0.00 Xgpw7433 6DL 3 4 0.12 0.50 Xcfe95 6D 1 1 0.00 0.00 Xgpw7386 7AL 9 8 0.74 0.79 Xgpw7185 7AS 3 2 0.05 0.07 Xgpw7288 7AS 8 8 0.59 0.76 Xcfe248 7A 1 1 0.00 0.00续表 2
Continue table 2
Xgpw7596 7BL 2 2 0.16 0.40 Xgpw7320 7BS 11 11 0.66 0.77 Xgpw7342 7BS 9 11 0.70 0.77 Xcfe135 7D 2 2 0.36 0.25 Xgwm99 1AL 12 11 0.52 0.72 Xgwm135 1AL 21 13 0.74 0.78 Xgwm11 1BS 16 13 0.80 0.81 Xgwm413 1BS 16 14 0.78 0.86 Xgwm642 1DL 9 5 0.63 0.58 Xgwm337 1DS 19 15 0.82 0.87 Xgwm312 2AL 26 24 0.86 0.87 Xgwm372 2AS 24 28 0.90 0.91 Xgwm257 2BS 8 3 0.56 0.60 Xgwm120 2BL 17 16 0.84 0.78 Xgwm539 2DL 27 31 0.89 0.90 Xgwm261 2DS 13 18 0.71 0.79 Xgwm2 3AS 6 9 0.46 0.71 Xgwm480 3AL 11 11 0.24 0.67 Xgwm285 3BS 37 34 0.75 0.87 Xgwm566 3BS 7 11 0.75 0.81 Xgwm664 3DL 4 5 0.20 0.49 Xgwm341 3DS 19 20 0.87 0.85 Xgwm610 4AS 13 12 0.58 0.74 Xcfd17h8a 4AL 8 8 0.34 0.48 Xgwm251 4BL 15 15 0.83 0.77 Xgwm149 4BL 8 12 0.49 0.62 Xcfd17h8d 4DL 16 17 0.87 0.88 Xgwm415 5AS 5 4 0.62 0.49 Xgwm186 5AL 15 23 0.83 0.85 Xgwm408 5BL 16 14 0.78 0.87 Xgwm234 5BS 12 10 0.85 0.82 Xgwm272 5DL 8 5 0.65 0.62 Xgwm190 5DS 10 10 0.68 0.69 Xgwm427 6AL 16 16 0.84 0.78 Xgwm219 6BL 20 17 0.86 0.82 Xgwm626 6BS 8 7 0.54 0.50 Xgwm469 6DS 14 14 0.84 0.82 Xgwm325 6DS 8 8 0.73 0.84 Xgwm260 7AS 20 15 0.78 0.86 Xgwm400 7BS 15 13 0.78 0.84 Xgwm46 7BS 19 19 0.85 0.86 Xgwm44 7D 14 17 0.86 0.88 Xgwm437 7DL 20 23 0.81 0.89 黑体表示Genomic-SSRs 引物及其检测数据,非黑体表示 EST-SSRs 引 物及其检测数据Bold represents Genomic-SSRs primer and data tested, the others represents EST-SSRs primer and data tested
部供试材料中的频率。出现频率低于
5%的等位变异
被 界 定 为 稀 有 等 位 变 异 。 多 样 性 指 数 为
H
t=
∑
= k i i/
k
PIC
1,其中
k 表示所检测的基因座总数。
1.2.6
品种间遗传距离的估算 用 NTSYS2.10
[9]软
件中的
DICE 系数表示品种间的遗传相似性,则两个
品种间的遗传距离
D=1-DICE。
1.2.7 组间遗传距离的估算
用 GENETIX
[7]软件中
的
Nei 氏 公 式
[11]计 算 组 间 的 遗 传 距 离 , 即 :
D=D
12(m)-[D
11(m)+ D
22(m)]/2 , 其 中
D
12(m)=1-
m
P
P
m i mi mi∑∑
1 2,
D
11(m)=1-
m
P
m i mi∑∑
2 1,
D
22(m)、D
33(m)…
D
nn(m)计算方法与
D
11(m)算法完全相同,
P
1mi代
表第
1 组第基因座第 i 种等位变异的频率,m 表示所
检测的基因座总数。
1.2.8
标记/性状的关联分析 中国材料于 2001~
2002 年、欧洲材料于 2004~2005 年种植于河南洛阳
市农业科学院,每份材料种植
2 行,行长 2 m,每行
播种
40 粒,在全生育期进行农艺性状的观察和记载。
根据
Flint-Garcia 等
[12,13]和张学勇等
[14]介绍的方法进
行标记
/性状关联分析。由于欧洲、东亚品种明显分成
两大群,因此,关联分析按欧洲、中国两组进行,结
果互相验证。为了排除标记
/性状之间出现伪关联,用
r
2(squared allele-frequency correlations)对连锁不平
衡程度进行检测。具体作法如下,假设考察的两个基
因座
, 其中一个具有等位变异 A 和 a, 另一个具有 B
和
b, 它们在群体中的等位变异频率分别为 π
A、
π
a、
π
B和
π
b,
4 种可能单倍型 AB、Ab、aB 和 ab 的频率分别
为
π
AB、
π
Ab、
π
aB、及
π
ab,那么
b B a Α abπ
π
π
π
D
r
2=
(
)
2,
r
2的
取值范围介于
0~1,其中,D
ab=(π
AB-π
Aπ
B)。通常,
r
2值越大,两基因座间的连锁不平衡性越强
[12]。在实
际操作中常用
TASSEL
[15]软件进行运算。
2
结果与分析
2.1 SSR 基因座的多态性分析
用
82 对 SSR 引物对 734 份材料进行基因型扫描,
共检测到
865 个等位变异。每个引物的等位变异为 1~
50 个,平均 10.42 个;多态性信息含量 0~0.91,平均
0.51。通常基因座的等位变异越多,其多态性信息
含量越高(如
Xgwm539),但也有少数基因座,尽管
等位变异较多,但其多态性信息含量仍然很低(如
Cfe52)。
2.2 不同地理来源和不同时期育成品种的遗传多样
性比较分析
2.2.1 欧洲和东亚材料的遗传多样性分析
欧洲、东
亚小麦材料检测到的总等位变异分别为
730、716;平
均遗传丰富度分别为
8.80 和 8.61;平均遗传多样性指
数为
0.46 和 0.52;特异等位变异分别为 150(20.55%)
和
135(18.85%)。总体上,东亚材料的遗传多样性水
平略高于欧洲。用
NTSYS 2.10 软件对 734 份材料进
行遗传主成分分析(Principal coordinate analysis,简
称
PCO 分析),发现欧洲和东亚材料明显分为两大群
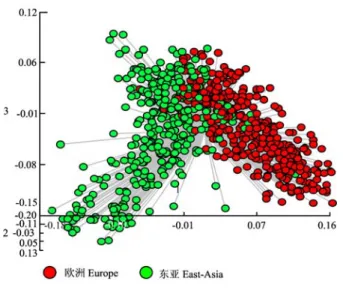
(图
1),表明欧洲和东亚材料在遗传组成上发生了明
显的分化。
图 1 734 份材料的 PCO 聚类图
Fig. 1 PCO analysis of 734 accessions based on 82 SSR loci
2.2.2 不同地理组材料的遗传多样性比较
依据材
料来源将
734 份材料分成 35 组,其中,欧洲材料 18
组,东亚材料
17 组。对 35 组各自总的等位变异、稀
有等位变异、平均遗传丰富度及遗传多样性指数进行
了比较分析(表
1)。结果发现,印度+巴基斯坦+缅
甸+克什米尔组的等位变异数最多(
379),平均遗传
丰富度最高(
4.57),遗传多样性指数最高(0.50);福
建+广州的等位变异数最少(222),平均遗传丰富度
最低(
2.67);甘肃的稀有等位变异数最多(125);荷
兰的遗传多样性指数最低(0.38)。总体上,在欧洲材
料中,东南欧国家(罗马尼亚、保加利亚、塞尔维亚、
匈牙利)、地中海国家(意大利、西班牙)的材料遗传
多样性最高,中欧国家(德国、瑞士、奥地利、捷克
斯洛伐克、波兰)次之,西欧国家(法国、比利时、
英国、荷兰)最低,这与
Roussel 等
[16]的结果一致;
在东亚材料中,印度、巴基斯坦、缅甸、克什米尔组
的遗传多样性最高,中国北方(北京、山西、河北、
河南、陕西、山东)次之,中国南方(安徽、湖北、
浙江、福建、广东、四川、云南、贵州)最低。
采用
GENETIX 软件
[7]的
Nei78 程序
[11]求得
35 组
的遗传距离,通过
PHLIP 软件
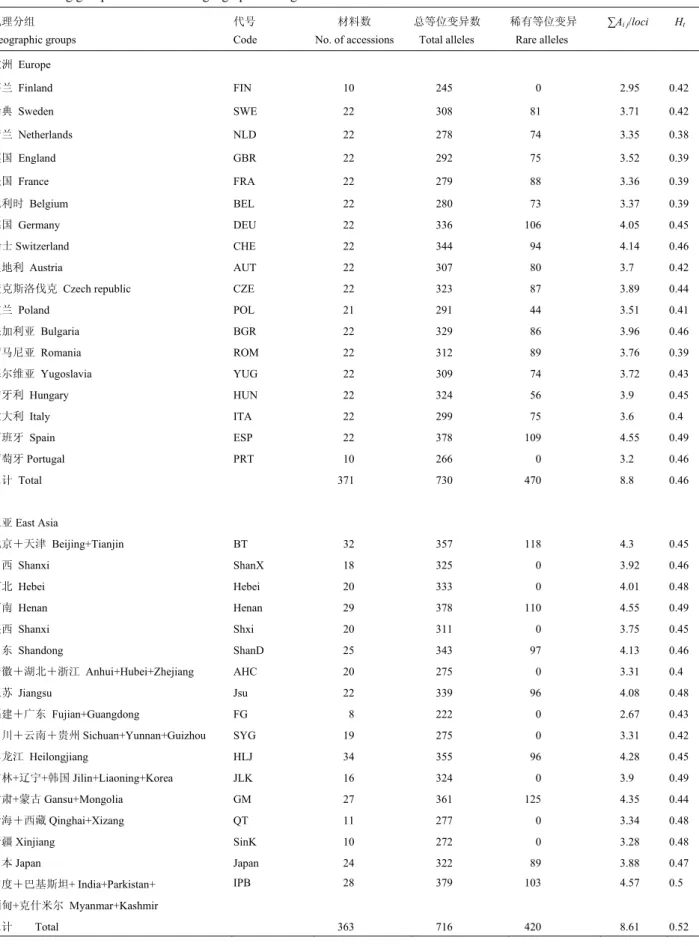
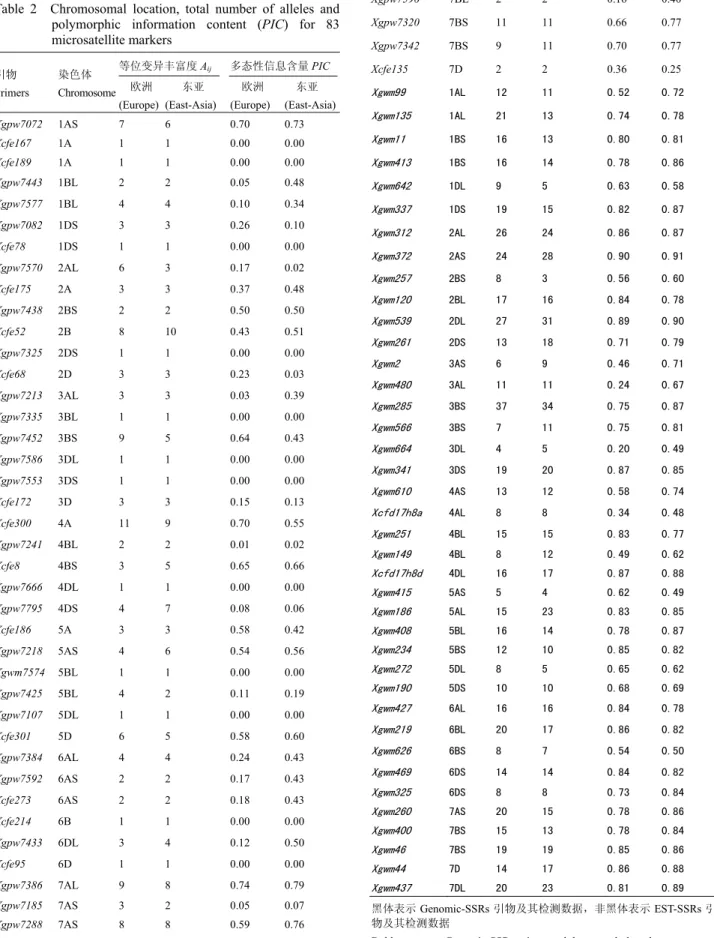
[10]聚类,结果发现整个
聚类图由两部分组成。第一组全由欧洲材料组成,第
二组除了葡萄牙以外,都来自东亚(图
2)。在欧洲组
中,第一类由东南欧国家材料组成(罗马尼亚、保加
利亚、塞尔维亚、匈牙利)
;第二类分别由西欧国家材
料(包括法国、比利时、英国、荷兰)和中欧国家材
料(德国、瑞士、奥地利、捷克斯洛伐克、波兰)组
成,而芬兰和南欧意大利材料各自成一类。在东亚组
中,中国北方与南方材料基本分成了两类。印度、巴
基斯坦、克什米尔、缅甸的材料与我国东北春麦区的
材料聚在一起;青藏和新疆冬春麦区的材料和北部冬
麦区的材料聚在一起。中国南方类中包括了长江中下
游冬麦区、西南冬麦区和华南冬麦区的材料;但河南、
陕西和甘肃的材料也聚在这类。日本材料基本上自成
一类,遗传上与中国南方品种相近(图
2)。
从聚类图还可以看出,材料间的遗传关系与其原
产地密切相关,来自于相邻地区的材料往往聚在一起,
这与
Huang 等
[17]、
You 等
[18]的结果相吻合。另外,东
亚材料间的遗传差异大于欧洲材料。
图 2 734 份材料分成 35 组后的聚类结果
Fig. 2 Neighbor-Joining tree based on the SSR data after the 734 collections were divided into 35 groups based on their
geographical origins
为了解各组内品种的差异来源,用
ARLEQIN 软
件
[8]对
35 组的分子数据进行方差分析(表 3),发现尽
管欧洲与东亚间、
35 组间的方差成分都达到了极显著
水平,但它们所解释的变异只占总变异的
5.87%、
9.48%,主要变异(84.65%)还是来自组内材料间的
差异。说明尽管地理位置和气候环境对遗传变异有重
要影响,但材料本身的变异、特别是人为的引种、杂
交和育种选择对材料的变异影响更大
[19]。
2.2.3 欧洲和东亚材料间等位变异频率的差异 为
寻找欧洲及东亚材料聚为两类的原因,笔者对
82 对
表 3 欧洲、东亚及 35 组的方差分析(AMOVA)
Table 2 Analysis of molecular variance (AMOVA): effect of geographical group
来源 Source of variation 自由度df 方差成分Variance components 变异比例 Variation accounted for(%) 欧洲、东亚组间Variation between Europe and Asia 1 1.19 Va*** 5.87
35 组间 Variation among the 35 groups 34 1.92 Vb*** 9.48 组内材料间Variation within the groups 699 17.16Vc*** 84.65
总体Total 733 20.28 *** 差异极显著 P<0.001 *** indicates P<0.001
SSR 的等位变异及其在欧洲、东亚及 35 组出现的频率
进行了统计分析,发现
42 对(50.6%)SSR 引物(如
Xgwm135、Xgwm149、Xgpw7795、Xcfe301 等)的等
位变异类型及其频率分布在欧洲和东亚材料间基本一
致(图
3),而 41 对(49.4%)SSR 引物(如 Xgpw7596、
Xgpw7433、Xgpw7443、Xgwm186、Xgwm257 等)的
等位变异组成或其频率在欧洲和东亚材料间表现出较
大差异(图
4)。例如,在 Xgpw7596(EST-SSR)基
因 座 , 检 测 到 两 种 等 位 变 异 即
Xgpw7596-98 和
Xgpw7596-130。91%的欧洲材料携带 Xgpw7596-130
等位变异,而在东亚材料中携带
Xgpw7596-130 的材
料仅占
73%。从图 4 可清楚看出在欧洲各国的材料中,
除了东南欧的保加利亚、罗马尼亚、塞尔维亚及地中
海地区的意大利、葡萄牙、西班牙的少数材料携带
98
bp 的等位变异外,大都只具有 130 bp 这一等位变异;
但中国的
15 个组群都同时具有 98 bp 和 130 bp 两个等
位变异,而且南方的四川、云南及贵州组、福建和广
图 3
欧洲和亚洲材料在
Xgwm135
基因座等位变异组成及其
频率
Fig. 3 Alleles and their frequency at
Xgwm135 in European
and East Asian wheat collections
图 4
Xgpw7596
引物(
7BL)在不同地理组的等位变异频率分布
州组的材料以
98 bp 等位变异为主。正如 Korzun 等
[20]所言,自然和人工选择使材料为适应不同的地理环境,
在某些基因座的等位变异发生了改变。推测正是这类
基因座在不同地区的等位变异组成及其频率的差异,
致使欧洲和东亚材料聚为明显不同的两大类(图
2)。
2.2.4 不同时期育成品种的遗传多样性比较
根据
品种(系)的育成年限,将
338 份欧洲材料及 291 份
中国材料分成
14 组。依据 82 对 SSR 引物的分子数据,
采用
GENETIX 软件
[7]、
NTSYS2.10 软件
[9]对其进行聚
类。所有供试材料(图
5)聚成三大类:欧洲材料聚
为一类,中国
50 年代以后育成的品种聚为一类,中国
的地方品种及上世纪
40 年代以前育成的品种聚为一
类。特别值得一提的是中国
1950 年以后育成的品种与
欧洲材料聚成一大类,而远离中国地方品种和此前育
成的品种,这与中国的育种历史基本相吻合。在中国,
80 年代育成的品种有别于 50~70 年代育成品种,可
能与
70 年代中国育种开始利用从欧洲引进的 1B/1R
材料有关。
欧洲的育成品种与地方品种之间也发生了大的分
化,
1970~1989 年育成的品种又明显不同于此前的品
种,此次品种分化可能与矮秆基因和
1B/1R 材料的广
泛利用密不可分;
1990 年以后育成的 44 个品种又显
著不同于
1836-1989 年间育成的品种(图 5),造成这
次分化的原因有待进一步分析。
(1)~(8)欧洲材料;(9)~(14)中国材料 (1)-(8) European groups; (9)-(14) Chinese groups图 5
中国和欧洲不同时期育成品种的聚类图
Fig. 5 Dendrogram of European and Chinese collections based on
82 SSR after being grouped by their release time
2.3 Genomic-SSR 与 EST-SSR 的聚类结果比较
在检测到的
865 个等位变异中,681 个来自
Genomic-SSR 引物,每对引物的等位变异为 5~50,
平均
17.46;多态性信息含量为 0.37~0.91,平均 0.76;
184 个来自 EST-SSR 引物, 其中 13 对引物在 734 份
材料中未检测到多态性,每对引物的等位变异为
1-14,
平均
4.18;多态性信息含量为 0~0.78,平均 0.28。从
平均遗传丰富度和遗传多样性指数这两项指标来看,
Genomic-SSR 揭示的遗传多样性远高于 EST-SSR(表
4)。
表 4 Genomic-SSR 与 EST-SSR 的遗传多样性比较
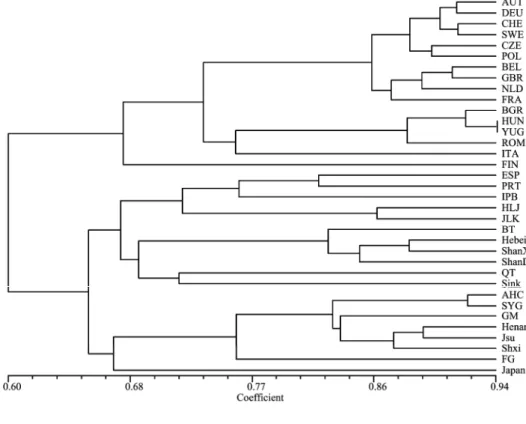
Table 4 Genetic diversity between Genomic SSR and EST-
SSR
EST-SSR Genomic-SSR 总体 Total 检测基因座数 Loci 44 39 83 总的等位变异数 ∑Aij 184 681 865 平均等位变异丰富度 ∑Aij/loci 4.18 17.46 10.4 平均遗传多样性指数 Ht 0.28 0.76 0.51
依据
Genomic-SSR 和 EST-SSR 的分子数据分别
对
35 组进行聚类(图 6,图 7),可以清楚地看出基于
Genomic-SSR 及 EST-SSR 的聚类图除个别地区的位置
发生变化外,整体趋势是一致的。与图
2 相比,图 6
中西班牙与葡萄牙聚在东亚组,意大利聚在了东南欧
组。图
7 中罗马尼亚和甘肃的位置发生了大的改变,
罗马尼亚组从东南欧组中分出而与芬兰聚在一起,甘
肃+蒙古组从东亚组的南方类中分出聚到了北方类。
另外,从聚类图可以看出,图
6 的相似性系数较低,
在
0.60~0.95 之间;而图 7 的较高,在 0.92~1.00 之
间。因此,
Genomic-SSR 较 EST-SSR 更易发生变异,
由育种和自然选择所引起的分化更快(图
6,图 7)。
2.4 分子标记与农艺性状的关联分析
根据
Sourdille 等发表的小麦遗传图谱
[21],求出每
一条染色体不同引物对间的遗传距离,位于不同染色
体两引物间的遗传距离以
250 cM 表示。用 TASSEL2.0
软件
[15]求出
70 个 SSR 引物两两间的 r
2,除
6A 短臂
上的
Xcfe273 与 Xgpw7592 间的 r
2在全体、中国及欧
洲材料中分别为
0.66、0.72 和 0.52 外,其他组合的变
异范围在
0.00002~0.23(图 8,由于 Xgpw7592 在染
色 体 上 的 确 切 位 置 未 知 , 无 法 求 出
Xcfe273 与
Xgpw7592 间的遗传距离,所以其 r
2未在图
8 中显示)。
因为
r
2较小,所以笔者认为在所研究的中国材料、
图 6 基于基因组 SSR 数据 35 组的聚类结果
Fig. 6 UPGMA dendrogram of 35 geographical groups based on 38 Genomic-SSR loci
图 7 基于 EST-SSR 数据 35 组的聚类结果
图 8 70 个 SSR 基因座间的连锁平衡性检测
Fig. 8 The pattern of LD for 70 SSR loci
欧洲材料均不存在突出的群体结构问题。
Xcfe273 和
Xgpw7592 位于同一染色体臂上(6AS),
对农艺性状的影响趋势非常相似,存在明显的单倍型
(
Haplotypes)现象,如在 322 份欧洲材料中,89.75%
的材料携带
Xgpw7592-279 + Xcfe273-336,在东亚材料
中,此单倍型所占的比例为
68.49%,推测这两个
EST-SSR 基因座相距很近。
由于
PCO 分析显示东亚和欧洲材料清楚地聚为
两大类,因此笔者对其分别进行标记
/性状关联分析,
以寻找与两大洲小麦生物学特性分化有关的基因组区
域,这样也就避免了两套材料非同年种植对分析所带
来的困难。对有多态性的
70 对 SSR 引物的等位变异
在欧洲及东亚材料中出现的频率进行分析,发现有
39
基因座(
55.7%)的等位变异出现严重的偏分布现象,
笔者对其中的部分引物与农艺性状进行关联分析,在
4B、5A、6A、7B 染色体上发现 6 个与穗粒数、千粒
重、株高、生育期、有效分蘖等相关联的基因座(表
5)。
其中
7B 染色体上 Xgpw7596 位点 98 bp 和 130 bp 的在
表 5 欧洲、中国小麦品种 6 个 SSR 基因座与农艺性状的关联分析
Table 5 Association analysis of 6 SSR loci with agronomic traits
等位变异(bp)及频率 Major alleles and frequency 穗粒数 Kernels/spike 株高 Plant height (cm) 基因座
Loci
染色体
Chromosome 欧洲 Europe 中国 China 欧洲 Europe 中国 China 欧洲 Europe 中国 China
153 62.53% 153 42.70% - - - - Xgwm149 4BL 其它others 29.11% 其它 others 21.76% - - - - 131 19.95% 131 19.28% 46.03±0.91 - 86.86±1.89 - Xgwm415 5AS 133 50.67% 133 45.18% 43.65±0.54 - 97.21±1.26 - 125 23.99% 121 50.69% - 53.79±1.13 - - Xgwm186 5AL 101 21.02% 其它others 37.47% - 50.99±0.83 - - 336 81.40% 336 42.70% 43.64±0.42 50.58±0.79 102.12±1.40 96.65±1.03
Xcfe273 (EST) 6AS
303 9.43% 303 19.56% 50.27±1.40 53.83±1.28 92.91±1.89 79.26±2.07 279 80.32% 279 42.42% 43.59±0.42 50.67±0.80 96.43±1.01 101.91±1.42 Xgpw7592 (EST) 6AS 288 8.90% 288 19.01% 51±1.45 53.85±1.31 79.06±2.20 92.45±1.94 130 82.75% 130 40.22% 43.94±0.43 49.28±0.72 95.93±1.04 101.57±1.51 Xgpw7596(EST) 7BL 98 7.55% 98 22.59% 49.33±1.58 55.83±1.26 81.36±2.15 93.57±1.69 有效分蘖 Ears/plant 开花期 Flowering time (days) 千粒重 TKW (g)
基因座 Loci 染色体
Chromosome 欧洲 Europe 中国 China 欧洲 Europe 中国 China 欧洲 Europe 中国 China 9.44±0.21 - - 182.14±0.41 42.20±0.47 Xgwm149 4BL 10.25±0.33 - - 184.08±0.59 39.41±0.73 - - 201.59±0.53 - - 39.292±0.69 Xgwm415 5AS / - 204.05±0.47 - - 42.41±0.40 - - 206.10±0.62 183.91±0.46 34.57±0.54 Xgwm186 5AL - - 200.19±0.54 181.05±0.48 36.42±0.50 9.84±0.19 8.37±0.19 203.92±0.35 184.25±0.42
Xcfe273 (EST) 6AS
8.42±0.50 6.89±0.26 197.71±0.83 179.81±0.43 9.83±0.19 8.32±0.19 203.84±0.36 184.21±0.43 Xgpw7592 (EST) 6AS 8.24±0.51 6.86±0.27 197.85±0.88 179.78±0.43 8.28±0.20 203.82±0.35 184.20±0.45 - 39.80±0.45 Xgpw7596(EST) 7BL 7.03±0.23 196.43±0.66 180.26±0.38 - 43.67±0.74
欧洲和中国的地理分布特点(图
4),明显与对早熟性
的要求不同相关,前者比后者早开花
4~7 d(表 5),
98 bp 主要在一年两熟的国家或地区,小麦成熟早晚对
下季作物的播种和产量影响很大;而
130 bp 则在一年
一季的地方频率很高(图
4)。
3 讨论
3.1 欧洲和中国不同时期育成品种遗传多样性的变化
中国的育成品种聚为两类:
1950 年以前育成品种
和地方品种聚为一类,此后育成的品种(1950~1980s)
聚为另一类,后者与欧洲材料相近。这与中国的小麦
育种历史有关。
1948 年以前育成的品种只有 39 个,
其中,28 个(71.9%)是从地方品种中混选或选育成
的
[1]。它们与地方品种聚为一类是必然的。追踪本研
究所涉及的
1950 年以后中国育成品种的系谱,发现
50%以上的品种含有欧洲品种的血缘(数据未发表)。
南大
2419、阿勃、阿夫、郑引 4 号、洛夫林 10、洛夫
林
13 等是 50~70 年代中国小麦育种的骨干亲本
[1]。
此外,欧洲品种中
Ardito(矮立多)、Villa Glori(中
农
28)、Tevere(吉利)、Guiliari(矮秆红)、Forlani
(弗兰尼)、
Virgolio(维尔)、Bulgarian10(保 10)、
Bulgarian10(保 301)、Jubileina Ⅰ(尤皮 1 号)、Jubileina
Ⅱ(尤皮
2 号)、Heine Hvede(亥恩·亥德)、Etoile de
choisy(舒瓦极星)、San Pastore(阿桑)、Mara(玛拉)、
Neuzucht(牛朱特)等也在中国小麦育种中被成功利
用。由此可见,欧洲品种对中国小麦育成品种的影响
很大,这一时期的品种与欧洲的品种聚为一类不足为奇。
欧洲品种中,地方品种明显地单独聚为一类。在
育成品种中,
1929 年以前育成的品种为一类,20 世纪
30~60 年代的品种聚为一类,70~80 年代的品种聚为
一类,而
90 年代的品种又发生了重要的变化,这与
Roussel 等
[16]的研究结果相吻合。说明从
70 年代开始,
欧洲的育种发生了较大的变化。表型研究说明,70 年
代欧洲小麦的矮化育种取得了突破性进展
[3]。欧洲
70
年代以后育成的品种,特别是
90 年代育成的品种,应
该引起中国育种家的注意。
最令人惊讶的是,一些基因座如
Xgpw7596(7B)、
Xgpw7433(6D)、Xgwm186(5A)等的组成的确在欧
洲、东亚之间存在十分明显的差异(图
4、表 3),表
明欧洲、东亚之间材料的遗传分化带有一定的必然性。
这也就是今天绝大多数从欧洲引进品种不能直接应用
于生产,需要对其进行杂交、回交改造之后,方能应
用于生产的原因所在,其实质就是对一些重要基因座
等位变异(基因)进行置换(图
4)。当然,这也与中
国小麦育种水平及品种综合遗传特性的提高是分不开
的,杂交、回交、选择的过程实际也是取其精华的过
程。
3.2 东亚、欧洲品种之间多基因座比较分析结果对育
种工作的启示
将近一个世纪的小麦育种,对遗传多样性产生了
很强的影响(瓶颈效应),削弱了品种对威胁性病虫害
的抵御或缓冲能力
[2,22,23]。与老品种相比,现代品种在
许多影响重要农艺性状的基因座上拥有更优异的等位
变异
[3,24],但就每一个品种而言,并非所有重要基因
座上所携带的都是最优异的等位变异。如在
6A 染色
体上的
Xcfe273(EST-SSR)引物,303 bp 等位变异无
论对株高、穗粒数、早熟性(抽穗期)的影响都优于
优势等位变异
336 bp,但其比例在欧洲和东亚材料中
都是很低的,分别为
9.43%和 19.56%(表 3),因此,
众多品种还有改进的余地。类似的可供育种家参考的
重要引物还有
7BL 上的 Xgpw7596(EST-SSR),对株
高、穗粒数、早熟性(抽穗期)
、千粒重等性状的改良
和提高都有很好的潜在价值,但其频率并不高,这可
能也是品种的产量水平还在缓慢提高的原因之一。
4 结论
欧洲与东亚小麦品种已明显分化为两大类群。通
过标记/性状关联分析,在
4B、5A、6A、7B 染色体
上发现
6 个影响穗粒数、千粒重、株高、抽穗期、有
效分蘖等重要农艺性状的基因座,其中个别基因座优
势等位变异差异可能与东亚、欧洲品种的分化密切相
关。中国
20 世纪 50~80 年代育成品种在大的聚类上
明显靠近欧洲材料、而远离中国
50 年代以前的育成品
种和地方品种,与中国的育种实际相吻合。在未来的
小麦育种中,需要通过杂交、回交,对欧洲品种的一
些重要基因座等位变异(基因)进行置换,取其精华,
方能实现“洋为中用”的目的。
致谢:谨以此篇献给中国农业科学院 50 华诞,感谢对
中法小麦合作项目给予过支持和帮助的人们。
References
[1] 庄巧生主编. 中国小麦品种改良及系谱分析. 北京: 中国农业出版 社, 2003.Zhuang Q S. Chinese Wheat Improvement and Pedigree Analysis. Beijing: Agricultural Press, 2003. (in Chinese)
[2] 郝晨阳, 王兰芬, 张学勇, 游光霞, 董玉琛, 贾继增, 刘 旭, 尚勋 武, 刘三才, 曹永生. 我国五十年来育成小麦品种的遗传多样性演 变. 中国科学 C 辑, 2005, 35(5): 408-415.
Hao C Y, Wang L F, Zhang X Y , You G X, Dong Y S, Jia J Z, Liu X, Shang X W, Liu S C & Cao Y S. Genetic diversity in Chinese modern wheat varieties revealed by microsatellite markers. Science in China:
Series C Life Sciences, 2006, 49(3): 218-226.
[3] 董玉琛, 郝晨阳, 王兰芬, 张学勇, 高海涛, 张灿军. 358 个欧洲小 麦品种的农艺性状鉴定与评价. 植物遗传资源学报, 2006, 7(2): 129-135.
Dong Y C, Hao C Y, Wang L F, Zhang X Y, Gao H T, Zhang C J. Evaluation of agronomic traits of 358 wheat varieties introduced from Europe. Journal of Plant Genetic Resources, 2006, 7(2): 129-135. (in Chinese)
[4] Guyomarc'h H, Sourdille P, Charmet G, Edwards K J, Bernard M. Characterization of polymorphic microsatellite markers from Aegilops
tauschii and transferability to the D-genome of bread wheat. Theoretical and Applied Genetics, 2002, 104: 1164-1172.
[5] Röder M S, Korzun V, Wendehake K, Plaschke J, Tixier M H, Leroy P, Ganal M W. A microsatellite map of wheat. Genetics, 1998, 149: 2007-2023.
[6] Nicot N, Chiquet V, Gandon B, Amilhat L, Legeai F, Leroy P, Bernard M, Sourdille P. Study of simple sequence repeat (SSR) markers from wheat expressed sequence tags (ESTs). Theoretical and Applied
Genetics, 2004, 109: 800-805.
[7] Belkhir K, Borsa P, Chikhi L, Raufaste N, Bonhomme F. GENETIX
4.05, logiciel sous Windows TM pour la génétique des populations.
Laboratoire Génome, Populations, Interactions, CNRS UMR 5171, Université de Montpellier II, Montpellier (in France), 2004.
[8] Schneider S, Roessle D, Excoffier L. Arlequin (2.000). Genetics and Biometry Laboratory, Department of Anthropology and ecology, University of Geneva, Switzerland, 2000.
[9] Rohlf F J. NTSYS-pc: Numerical Taxonomy and Multivariate Analysis
System, Version 2.1. Exeter Software, Setauket, N.Y, 2000.
[10] Felsenstein J. PHYLIP: Phylogeny Inference Package, Version 3.65 (Department of Genome Sciences and Department of Biology, University of Washington), 2004.
[11] Nei M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics, 1978, 89: 583-590. [12] Flint-Garcia S,Jeffry M, Thornsberry E S, Buckler I V . Structure of
linkage disequilibrium in plants. Annual Review of Plant Biology, 2003, 54: 357-374.
[13] Flint-Garcia S, Thuillet A, Yu J M, Pressoir G, Romero S et al. Maize association population: a high-resolution platform for quantitative trait locus dissection. Plant Journal, 2005, 44: 1054-1064.
[14] 张学勇, 童依平, 游光霞, 郝晨阳, 盖红梅, 王兰芬, 李 滨, 董玉 琛, 李振声. 选择牵连效应分析: 发掘重要基因的新思路. 中国农
业科学, 2006, 39(8): 1526-1535.
Zhang X Y, Tong Y P, You G X, Hao C Y, Ge H M, Wang L F, LI B, Dong Y C, Li Z S. Hitchhiking effect mapping: a new approach for discovering agronomic important genes. Agricultural Sciences in
China, 2007, 6(3): 255-264.
[15] Edward Buckler Lab: Maize Diversity Research (http://www. maizegenetics.net/bioinformatics/).
[16] Roussel V, F. Exbrayat, Balfourier F. SSR allelic diversity changes in 480 European bread wheat varieties released from 1840 to 2000,
Theoretical and Applied Genetics, 2005, 111: 162-170.
[17] Huang X Q, Borner A, Röder M S, Ganal M W. Assessing genetic diversity of wheat (Triticum aestivum L.) germplasm using microsatellite markers. Theoretical and Applied Genetics, 2002, 105: 699-707.
[18] You G X, Zhang X Y, Wang L F. An estimation of the minimum number of SSR loci needed to reveal genetic relationships in wheat varieties. Information from 96 random accessions with maximized genetic diversity. Molecular Breeding, 2004, 14: 397-406.
[19] 张学勇, 庞斌双, 游光霞, 王兰芬, 贾继增, 董玉琛. 中国小麦品 种资源 Glu-1 位点组成概况及遗传多样性分析. 中国农业科学, 2002, 35(11): 1302-1310.
Zhang X Y, Pang B S, You G X, Wang L F, Jia J Z, Dong Y S. Allelic variation and genetic diversity at Glu-1 loci in Chinese wheat (Triticum aestivum L.) germplasm. Agricultural Sciences in China, 2002, 1(10): 1074-1082.
[20] Korzun V, Röder M S, Ganal M W, Worland A J, Law C N. Genetic analysis of the dwarfing gene (Rht8) in wheat. Part I. Molecular mapping of Rht8 on the short arm of chromosome 2D of bread wheat (Triticum aestivum L.). Theoretical and Applied Genetics, 1998, 96: 1104-1109.
[21] Sourdille P, Singh S, Cadalen T, Brown-Guedira G, Gay G, Qi L L, Gill B S, Dufour P, Murigneux A, Bernard M. Microsatellite-based deletion bin system for the establishment of genetic-physical map relationships in wheat ( Triticum aestivum L.). Functional &
Integrative Genomics, 2004, 4: 12-25.
[22] Bertin P, Gregoire D, Massart S, Froidmont D de. Genetic diversity among European cultivated spelt wheats revealed by microsatellites.
Theoretical and Applied Genetics, 2001, 102: 148-156.
[23] Röder M S, Wendehake K, Korzun V, Bredemeijer G, Laborie D, Bertrand L, Isaac P, Rendell S, Jackson J, Cooke R J, Vosman B, Ganal M W. Construction and analysis of a microsatellite-based database of European wheat varieties. Theoretical and Applied
Genetics, 2002, 106: 67-73.
[24] Tanksley S D, McCouch S R. Seed bank and molecular maps: Unlocking genetic potential from the wild. Science, 1997, 277: 1063-1066.
![Fig. 2 Neighbor-Joining tree based on the SSR data after the 734 collections were divided into 35 groups based on their geographical origins 为了解各组内品种的差异来源,用 ARLEQIN 软 件 [8] 对 35 组的分子数据进行方差分析(表 3) ,发现尽 管欧洲与东亚间、 35 组间的方差成分都达到了极显著 水平,但它们所解释的变异只占总变异的 5.87%、](https://thumb-eu.123doks.com/thumbv2/123doknet/14563636.538710/7.892.158.726.445.874/FigNeighborJoining为了解各组内品种差异来源软件组的分子数据进行方差分析.webp)