A novel multifunctional factor involved in trans-splicing of chloroplast introns in Chlamydomonas
13
0
0
Texte intégral
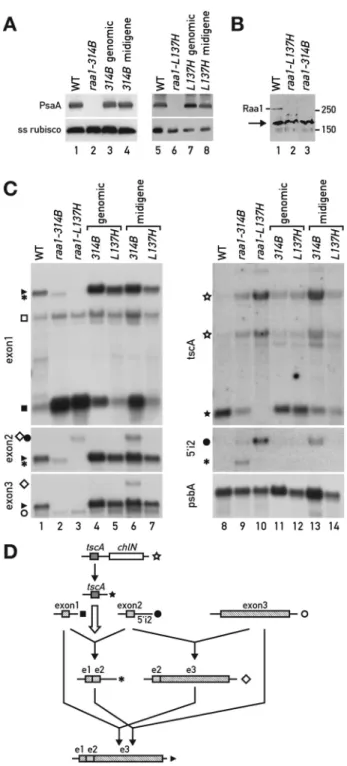
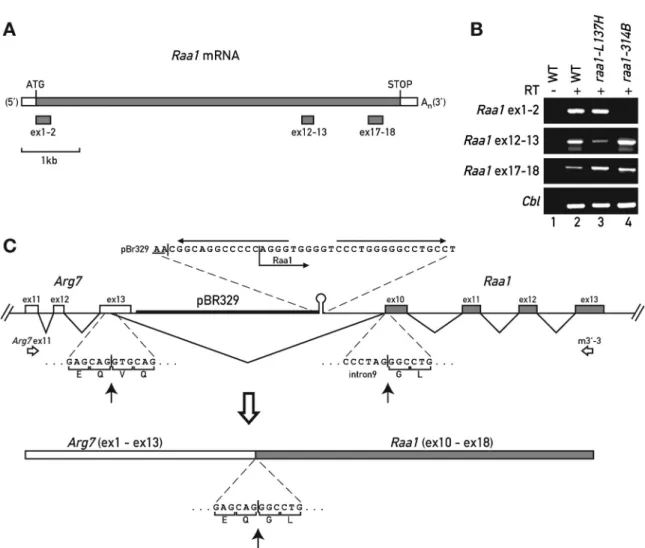
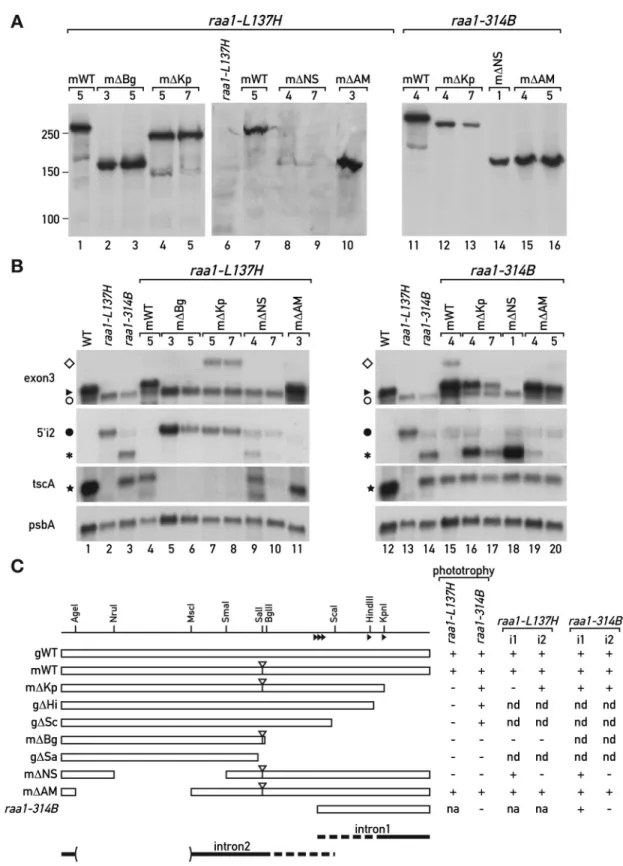
Figure

Documents relatifs