Assembly and nuclear export of pre-ribosomal particles in budding yeast
18
0
0
Texte intégral
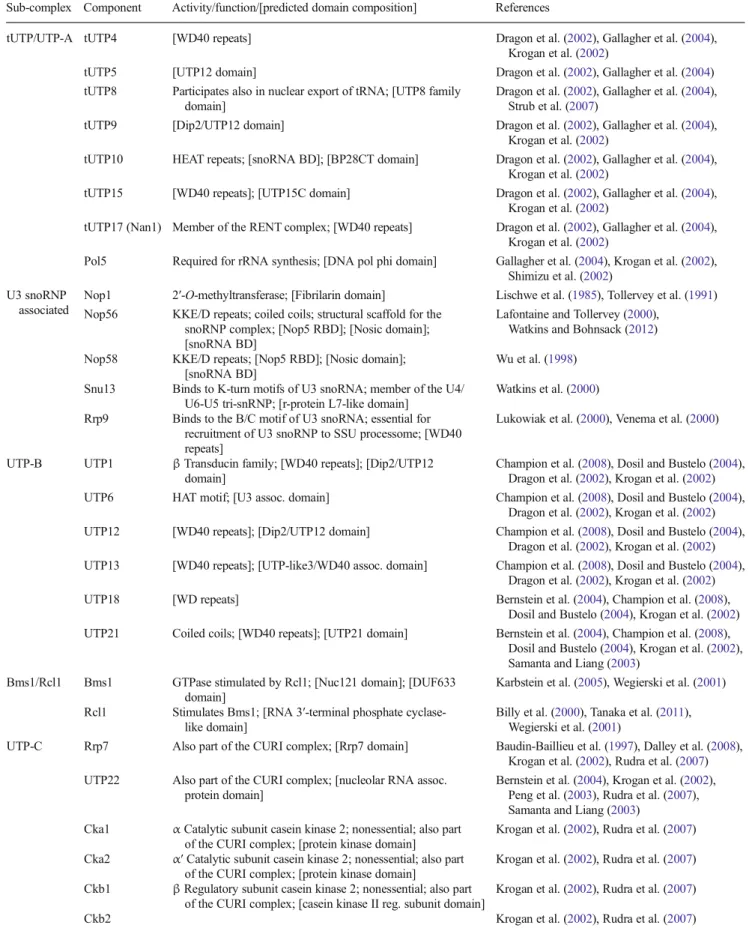
Figure

Documents relatifs
