Influence of Inorganic Phosphate on Photosynthesis of Wheat Chloroplasts : I. PHOTOSYNTHESIS AND ASSIMILATE EXPORT AT 5 °C AND 25 °C
Texte intégral
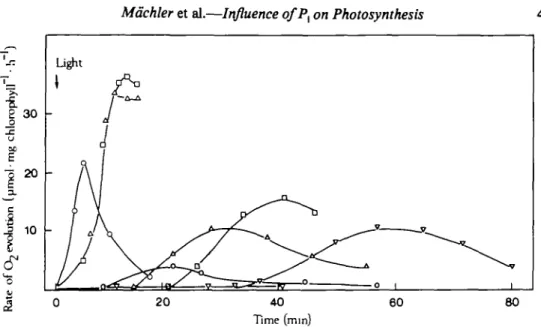
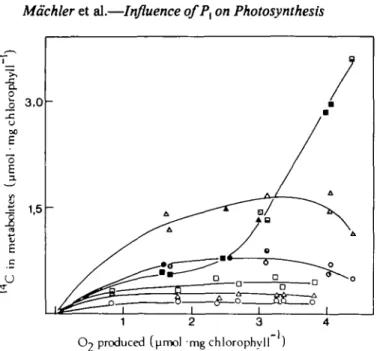
Figure
Documents relatifs
At larger training set sizes, the deep neural network performs much better; kernel ridge regression barely improves as training set size increases, however training wall time
In the present study, we analyzed the transcriptomic effects of in vitro embryo manipulations on the expression profile of placental genes (analyzed at 10.5 days post-fertilization,
Florent Foucaud, Eleonora Guerrini, Matjaz Kovse, Reza Naserasr, Aline Parreau, et al.. Extremal graphs for the identifying code problem.. The problem of finding an identifying code
4.2 Verifying MiniHTTPServer implementation As discussed in Section 2 , FreeSpec introduces two key com- ponents to reason about impure computations: (1) inter- face contracts
It was indicated above that Tumer's model is dynamic, with the ability to point to consequences for society. Thus, although the model does not neatly fit the lI-plus
Lorsque le cation stabilisateur est le tétraméthylammonium Me4W plus petit, les ligands dicarboxylates donnent avec SnPh3Cl des composés d'addition de type (1-4) à structures
One of the interesting questions is, if people’s preference lists are not strictly random, i.e., if we induce a conformity term into the preference structure, i.e., the agents
• We formulate the two-phase objective function under Friedkin-Johnsen model, where a node’s final opinion in the first phase acts as its initial bias for the second phase, and
