HAL Id: hal-02958427
https://hal.inrae.fr/hal-02958427
Submitted on 5 Oct 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Quels sont les facteurs de variation de la production
laitière des chèvres Créole allaitantes ? Quels sont les
meilleurs paramètres de croissance des chevreaux
permettant d’estimer indirectement la production
laitière ?
Coralie Machefert
To cite this version:
Coralie Machefert. Quels sont les facteurs de variation de la production laitière des chèvres Créole allaitantes ? Quels sont les meilleurs paramètres de croissance des chevreaux permettant d’estimer indirectement la production laitière ?. Sciences du Vivant [q-bio]. 2020. �hal-02958427�
MASTER Biologie Agrosciences
Parcours Sciences de l’Animal pour l’Élevage de Demain (SAED)
MASTER 1
Année 2019-20
Quels sont les facteurs de variation de la production laitière
des chèvres Créole allaitantes ?
Quels sont les meilleurs paramètres de croissance des chevreaux
permettant d’estimer indirectement la production laitière ?
Coralie MACHEFERT
INRAE Antilles-Guyane - URZ (Unité de Recherches Zootechniques)
Sous la Direction de Madame Nathalie MANDONNET
2
Remerciements
Je souhaite remercier particulièrement ma tutrice de stage Mme Nathalie MANDONNET, directrice de l’Unité de Recherches Zootechniques (URZ) à INRAE Antilles-Guyane, pour son accueil, sa disponibilité, sa gentillesse et son aide à la réalisation de ce mémoire. Aussi, je remercie M. Maurice MAHIEU pour toute l’aide qu’il m’a fournie et les connaissances qu’il a su me transmettre dans la partie statistique de mon mémoire. Je remercie également tous les chercheurs et techniciens de l’URZ pour leur partage et leur gentillesse, ce qui m’a permis d’apprendre une multitude d’informations sur les systèmes d’élevage en zone tropicale.
Je remercie tous les professeurs de l’Agrocampus Ouest de Rennes et les intervenants professionnels qui m’ont permis d’acquérir des connaissances scientifiques et techniques en amont de mon stage.
Enfin, je voudrais remercier ma famille et mes proches pour leur soutien et leurs encouragements.
3
Table des matières
Introduction ... 1
1. L’élevage de chèvres Créole allaitantes en Guadeloupe ... 2
1.1. La conduite des élevages caprins allaitants en zone tropicale ... 2
1.2. Le troupeau expérimental de chèvres Créole à l’URZ d’INRAE Antilles-Guyane ... 3
1.3. Création d’un nouveau modèle biologique : la chèvre Créole mixte ... 4
1.4. Matériels et méthodes ... 4
2. Les résultats ... 7
2.1. Estimation des facteurs de variation environnementaux ... 7
2.2. Choix du meilleur critère de croissance pour l’évaluation indirecte de la production laitière ………..…... 9
2.3. Discussion ... 9
Conclusion ... 10
4
Liste des abréviations :
INRAE : Institut National de la Recherche pour l’agronomique, l’alimentation et l’environnement GMQ : Gain Moyen Quotidien
PAT : Poids Age Type
1
Introduction
Les travaux conduits par l’unité de recherches zootechniques (URZ) d’INRAE Antilles-Guyane visent à l'amélioration des espèces locales (ruminants et porc) en zone tropicale humide. Les problématiques principalement étudiées portent sur la génétique, l’alimentation, la parasitologie et les systèmes d’élevage. De ces travaux, il ressort que la chèvre Créole de Guadeloupe est le fruit de nombreux métissages intervenus lors de la colonisation, de la sélection naturelle à laquelle elle a été soumise en milieu contraignant, et d’une exploitation à des fins multiples (traction, viande, lait, culture hindoue). Ce génotype représente donc un bon modèle d’étude des caractères d’adaptation au milieu car il n’a été sélectionné sur aucun caractère de production.
Pour initier de nouveaux travaux sur le contrôle génétique de la résilience et de l’allocation des nutriments chez des caprins soumis à un stress parasitaire, l’URZ souhaite comparer la souche actuelle de chèvre Créole avec une souche sélectionnée sur sa production laitière. L’objectif de ce mémoire est de rassembler les informations nécessaires à l’analyse de la faisabilité d’une telle sélection dans le troupeau expérimental d’INRAE-PTEA.
L’estimation de la production laitière a été beaucoup étudiée avant les années 2000 en race bovine et ovine (Ricordeau, 1960 ; Poujardieu et al, 1969 ; Le Neindre et Dubroeucq, 1973). Mais peu de travaux se réfèrent à l’étude de la croissance des chèvres à viande en zone tropicale.
Depuis 40 ans, les chevreaux sont pesés de leur naissance au sevrage à 3 mois par INRAE - PTEA. Cette base de données expérimentales permet d’actualiser les connaissances sur les performances zootechniques des chèvres Créole et d’en étudier les principaux facteurs de variation.
La conduite des élevages caprins allaitants en Guadeloupe et du troupeau expérimental caprin d’INRAE sera premièrement présentée. L’évaluation indirecte de la production laitière des chèvres via la croissance des chevreaux sous la mère sera étudiée dans ce mémoire. L’effet de la génétique et des facteurs environnementaux sur la croissance des chevreaux sera estimé. L’objectif final est d’estimer le temps de sélection nécessaire pour que la nouvelle souche diffère significativement de la souche actuelle en matière de production laitière.
2
1.
L’élevage de chèvres Créole allaitantes en Guadeloupe
1.1.
La conduite des élevages caprins allaitants en zone tropicale
La chèvre Créole est la race caprine locale de la Guadeloupe. Ces chèvres sont élevées traditionnellement pour leur viande. Elles sont issues du métissage de chèvres d’Afrique et d’Europe de l’Ouest. L’élevage en milieu tropical est soumis à des contraintes climatiques, alimentaires et pathologiques, auxquelles les chèvres locales Créole s’adaptent facilement contrairement à des races tempérées (Alexandre et Mandonnet, 2005).
Les chèvres Créole sont, pour la majorité, noires mais leur robe peut être de couleur multiple. Cette race caprine est reconnaissable par son poids et sa taille plus faible que les races caprines métropolitaines. Le poids moyen des boucs est de 38 kg et celui des femelles est de 28 kg. La taille moyenne des boucs au garrot et de 61 cm et 51 cm pour les femelles (Chemineau et al, 1984).
Le cheptel caprin en Guadeloupe est représenté par une majorité de chèvres croisées de type viande. Plus de la moitié des troupeaux (62%) associent des chèvres croisées et des chèvres Créole ; les élevages caprins de race Créole ne représentent que 4%. Les élevages caprins allaitants en Guadeloupe sont de petites tailles. La superficie moyenne d’un élevage caprin est de 7 hectares (Gunia et al, 2010). La taille d’un troupeau peut varier de 10 à 300 chèvres avec une
moyenne de 30 chèvres par exploitation (Arquet Remy, communication personnelle). Le cheptel caprin en Guadeloupe présente 20.000 têtes pour 500 exploitations (Agreste, 2007). L’âge d’abattage des cabris est de 12 mois pour un poids d’abattage entre 15 et 20 kg de poids vif (Mahieu Maurice, communication personnelle). La production de viande en petits ruminants s’élève à 210 tonnes équivalent carcasse. L’importation alimentaire est très importante en Guadeloupe, notamment concernant la viande caprine et ovine où environ 1880 tonnes de viande sont importées par an (Agreste, 2007).
Les troupeaux sont conduits toute l’année au pâturage de pangola ou de petit foin avec une complémentation en concentrés en période sèche. Les bâtiments d’élevage sont construits sur caillebotis pour des raisons de disponibilités en paille insuffisantes (Arquet Remy, communication personnelle).
Les chèvres Créole manifestent une activité sexuelle continue tout au long de l’année. Ces caractéristiques leur permettent de suivre facilement un rythme de reproduction intensif (Chemineau
et al, 1991). Elles sont classées parmi les meilleures chèvres en zone tropicale grâce à de bonnes qualités maternelles associées à un taux de fertilité élevé (+ 90%) et une bonne productivité numérique (2,67 chevreaux sevrés par femelle par an) – (Naves et al, 2011). Malgré son petit format, la production de viande par hectare obtenue (755 kilos de carcasse par hectare par an) est supérieure à celle des autres races caprines tropicales.
3
Les problématiques actuellement rencontrées dans les élevages caprins des Antilles sont premièrement un parasitisme important malgré l’utilisation d’anthelminthiques. Également, le vol de cabris provoque de grandes pertes dans les élevages Guadeloupéens.
1.2.
Le troupeau expérimental de chèvres Créole à l’URZ d’INRAE Antilles-Guyane
L’unité de recherches zootechniques (URZ) d’INRAE Antilles-Guyane étudie deux types de conduite d’élevage, le pâturage et la conduite en bâtiment en race locale allaitante caprine, ovine, bovine et porcine. Les expérimentations portent principalement sur l’alimentation et la pathologie parasitaire. Le troupeau de chèvres Créole d’INRAE Antilles-Guyane est présent sur l’unité expérimentale du domaine de Gardel. Sur l’île, l’élevage est situé en zone sèche. Le cheptel est composé de 440 têtes (Arquet Remy, communication personnelle).
Les chèvres sont élevées toute l’année au pâturage, dans un système d’élevage semi-intensif. En plein air, ces animaux sont sujets au parasitisme. Le pâturage tournant permet de limiter la pression parasitaire sur les petits ruminants. Chaque lot d’animaux pâture sur cinq parcelles au total. Le temps de présence des animaux sur une parcelle est d’une semaine. Le fourrage de base est le pangola (Digitaria decumbens). C’est une graminée fourragère largement utilisée aux Antilles. Le chargement des animaux sur les parcelles varie de 50 à 95 mères par hectare, et dépend du lot et du pourcentage de pertes liés au vol (Alexandre et al, 1997). En effet, 30 % de pertes des animaux présents sur la station expérimentale de Gardel sont dues au vol (Arquet Remy, communication personnelle).
Du fait de l’absence de période d’anoestrus saisonnier en chèvres Créole, trois périodes de lutte sont déterminées dans l’année : janvier, mai et octobre. Deux lots de femelles sont mis à la reproduction. Au sein de la station expérimentale de Gardel, les chèvres réalisent trois mise-bas en deux ans. Au pâturage, 10% de mâles sont introduits dans le troupeau de femelles depuis la fin des années 80 (Alexandre et al, 1997). Une durée de 4 mois sépare deux périodes de reproduction. La lutte se réalise donc à la suite du sevrage des chevreaux. Le sevrage se réalise à 3 mois afin de limiter la mortalité en post-sevrage.
En Guadeloupe, trois types de climat diffèrent dans l’année et font varier la production fourragère, le parasitisme et ainsi les performances zootechniques des animaux d’élevage. Les trois périodes de reproduction installées dans la station expérimentale de Gardel s’accordent avec ces trois types de climat. En début d’année, la saison est sèche et en fin d’année la saison est humide. À partir du mois de mai la saison, qualifiée comme intermédiaire, est la plus favorable dans les élevages. Les performances zootechniques telles que la productivité, les poids à âge type et les gains moyens quotidiens sont plus élevées. En saison humide, le parasitisme augmente ce qui conduit à des croissances plus faibles ainsi qu’une hausse de la mortalité (Alexandre et al, 1997).
4
1.3.
Création d’un nouveau modèle biologique : la chèvre Créole mixte
À l’origine, les ateliers caprins en Guadeloupe n’étaient pas spécialisés dans la production de viande. La chèvre Créole était élevée pour sa viande et son lait. Suite à la demande, les éleveurs se sont organisés au sein d’une coopérative en 1978, Cabricoop, afin de spécialiser les élevages dans la production de viande de cabri (Alexandre et al, 1991).
La question de recherche se concentre actuellement sur la conception d’un nouveau modèle biologique : la chèvre Créole mixte. L’étude des facteurs de variation de la production laitière via les performances des chevreaux sera réalisée au cours de ce mémoire. Les objectifs des prochains travaux de recherche seront d’étudier le caractère de résilience de cette race mixte. L’évaluation de la capacité d’adaptation de la race à un nouveau système d’élevage mixte pourra ainsi être étudiée. L’étude portera également sur l’allocation des nutriments chez un ruminant. L’allocation des nutriments concerne l’utilisation des ressources pour satisfaire les besoins de l’animal. Ces deux caractères, résilience et allocation des nutriments, seront étudiés chez la chèvre Créole mixte dans de futurs projets de recherche. Le but final serait d’élever un sous-groupe d’animaux de chèvres Créoles mixte au sein d’INRAE Antilles-Guyane à des fins scientifiques.
1.4.
Matériels et méthodes
Les données utilisées ont été collectées à l’unité expérimentale de Gardel à INRAE Antilles-Guyane. La base de données regroupe plus de 23.000 individus en race caprine Créole.
Mon travail personnel : La première partie de mon travail a été de réaliser plusieurs étapes de correction et de vérification des données enregistrées depuis 40 ans. Puis, j’ai pu sélectionner les principaux facteurs de variation environnementaux et génétiques à placer dans le modèle mixte et d’analyser les données afin d’estimer la production laitière à partir des performances de croissance des chevreaux. La totalité du script réalisé sur le logiciel R est disponible en annexe 1.
Présentation des fichiers : L’ensemble des données sont réparties dans trois fichiers distincts. Le premier présente les mesures effectuées durant la période d’allaitement, telles que les pesées des chevreaux. Le second fichier d’archives généalogiques stocke l’ensemble des animaux qui sont sortis de la station expérimentale, ainsi que des informations portant sur les performances de reproduction des femelles et le numéro des parents. Enfin un dernier fichier répertorie tous les animaux sevrés sur le domaine de Gardel depuis 1980.
5
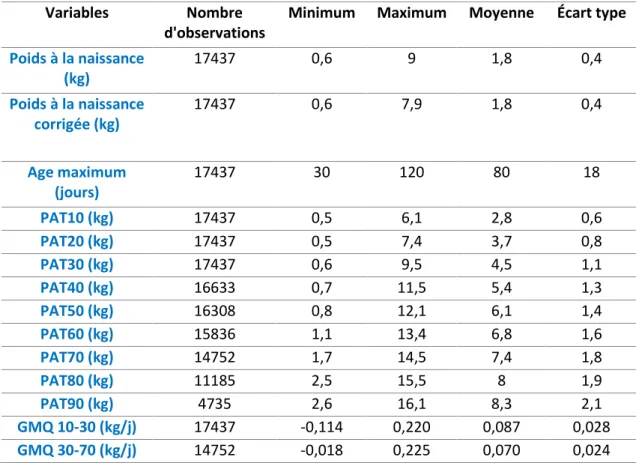
Tableau 1 : Moyennes phénotypiques, écart type, minimum et maximum des paramètres de
croissance des chevreaux Créole d’INRAE Antilles-Guyane de 1980 à 2019
Variables Nombre d'observations
Minimum Maximum Moyenne Écart type
Poids à la naissance (kg) 17437 0,6 9 1,8 0,4 Poids à la naissance corrigée (kg) 17437 0,6 7,9 1,8 0,4 Age maximum (jours) 17437 30 120 80 18 PAT10 (kg) 17437 0,5 6,1 2,8 0,6 PAT20 (kg) 17437 0,5 7,4 3,7 0,8 PAT30 (kg) 17437 0,6 9,5 4,5 1,1 PAT40 (kg) 16633 0,7 11,5 5,4 1,3 PAT50 (kg) 16308 0,8 12,1 6,1 1,4 PAT60 (kg) 15836 1,1 13,4 6,8 1,6 PAT70 (kg) 14752 1,7 14,5 7,4 1,8 PAT80 (kg) 11185 2,5 15,5 8 1,9 PAT90 (kg) 4735 2,6 16,1 8,3 2,1 GMQ 10-30 (kg/j) 17437 -0,114 0,220 0,087 0,028 GMQ 30-70 (kg/j) 14752 -0,018 0,225 0,070 0,024
6
Présentation des logiciels : Le travail d’analyse des données récoltées a été réalisé à partir de quatre logiciels : Libre Office, Logiciel R, SAS et VCE. Le logiciel Libre Office permet de visualiser l’ensemble des variables et covariables importantes à prendre en compte dans le modèle statistique. Le logiciel R permet de regrouper les données des trois fichiers de départ afin de réunir les informations concernant la généalogie, les mises-bas des femelles et les pesées des chevreaux. Plusieurs étapes de vérification des données ont pu être réalisées à partir de ces deux logiciels. Le logiciel SAS permet de préparer un fichier pedigree et un fichier performance dans l’optique de les analyser sur VCE. Le logiciel VCE permet d’estimer le potentiel génétique des individus précisément à partir de la généalogie et des performances des chevreaux.
Présentation, transformation et création des variables : Dans ce cas d’étude, les paramètres zootechniques concernant la mère et les pesées des chevreaux sont importants à prendre en compte. La croissance des chevreaux est évaluée par des pesées successives de la naissance jusqu’au sevrage (3 mois) avec un intervalle de 15 jours entre chaque pesée. Ces mesures ont permis de calculer les poids à âge type (PAT de 10 à 90 jours) ainsi que différents intervalles de gains moyens quotidiens (GMQ). La taille de la portée (de la naissance jusqu’au sevrage), et le sexe du chevreau sont également des variables d’intérêt zootechniques. La variabilité paternelle et maternelle est estimée grâce aux données généalogiques. La saison et l’année de la mise à la reproduction sont deux paramètres regroupés dans la variable « Lutte ».
Les dates de naissance et de pesées initialement sous forme de code ont été converties en format date. Les poids de chevreaux enregistrés en hectogrammes ont été recalculés en kilogrammes. Les variables année et saison de lutte ont été extraites de la variable Lutte. La saison de lutte est une variable à trois modalités : saison sèche, intermédiaire et humide. L’âge des animaux à chaque pesée a été premièrement calculé à partir des dates de pesées. Puis l’âge maximum de chaque chevreau a été déterminé afin de pouvoir calculer des poids à âge type uniquement sur les animaux ayant atteint l’âge du PAT calculé (PAT 50 estimé pour les chevreaux ayant un âge supérieur ou égal à 50 jours). Le numéro des animaux, la taille de portée, le sexe, la saison, l’année et le rang de mise-bas ont été transformés en variables qualitatives. Les variables quantitatives sont présentées dans le tableau 1.
Regroupement des variables : Les données des variables « rang de mise-bas » et « taille de portée (en cours d’allaitement) » ont été regroupées en classes selon les résultats de GMQ 10-30 et le nombre d’individus présents dans chaque modalité. Le rang de mise-bas de la chèvre a été scindé en 4 classes : 1 à 2 ; 3 à 7 ; 8 à 10 et supérieur ou égal à 11 mises bas. Les primipares et les chèvres ayant réalisées deux mises bas sont classées ensemble car elles produisent des chevreaux ayant un GMQ10-30 identique (83 g/j). La taille de portée a été dissociée en 3 classes différentes : simple, double, supérieur à 2 chevreaux.
7
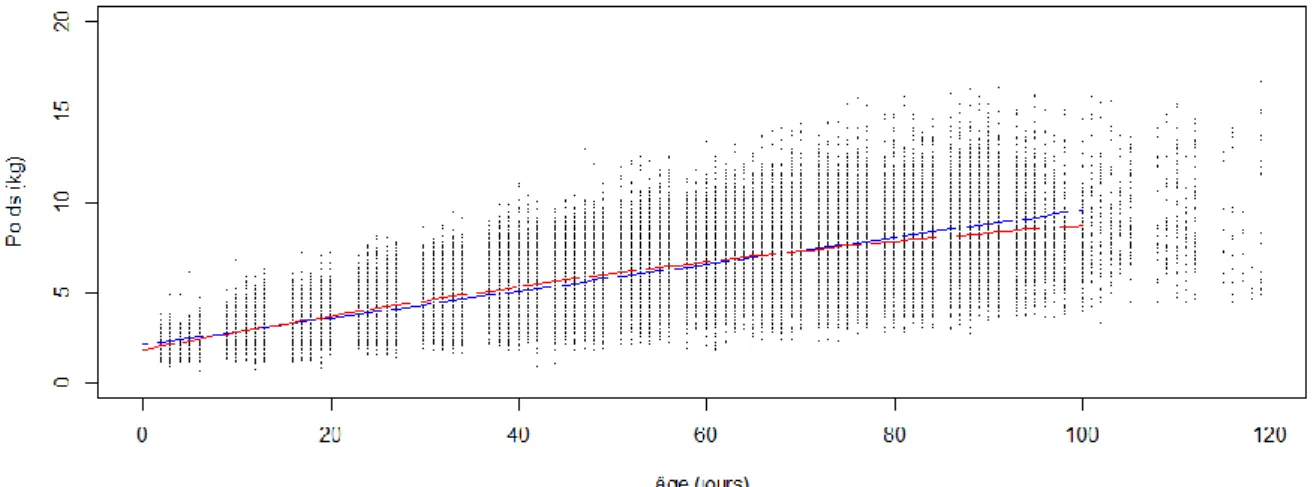
Courbe bleue (modèle linéaire) – courbe rouge (modèle quadratique)
Figure 1 : Évolution du poids des chevreaux de la naissance au sevrage
Figure 2 : Corrélation entre le poids à la naissance mesuré et le poids à la naissance
corrigé par le modèle quadratique
8
Les étapes de correction des données : En amont de l’analyse statistique des variables, plusieurs étapes successives de vérification et de correction des données ont été réalisées afin d’éliminer les valeurs aberrantes. Les erreurs de notation ont été premièrement supprimées ou corrigées. Une sélection des individus a été réalisée selon leur âge et le suivi de pesée jusqu’au sevrage. Les animaux ayant un âge minimum de 30 jours et un âge maximum de 120 jours ont été sauvegardés. Les chevreaux ne possédant pas de poids de naissance, de date de naissance ou de suivi de pesée n’ont pas été gardés dans la base de données. Les chevreaux ont une date et un poids unique pour chaque pesée. Les chevreaux ayant un poids supérieur à 22 kg ont été supprimés.
La taille de portée a été mesurée à la naissance, en cours d’allaitement et au moment du sevrage. La taille de portée en cours d’allaitement représente le nombre de chevreaux allaités par la chèvre depuis plus de 15 jours. Les chevreaux allaités artificiellement ou adoptés ont été éliminés du fichier afin d’estimer au mieux la production laitière de la chèvre. Les chevreaux ayant une taille de portée en cours d’allaitement dépassant 4 chevreaux ont été supprimés.
Une autre vérification des données a été réalisée selon la distribution normale des variables autour de la moyenne de la population. L’intervalle des poids à +/- 2 écart-types a été calculé pour les poids à âge type de 0, 10, 30, 50 et 70 jours afin de repérer les individus ayant des poids estimés incohérents. Après une observation fine des données brutes de ces chevreaux, douze d’entre eux ont été supprimés de la base de données. L’évolution des poids de ces chevreaux était physiologiquement incorrecte ou incohérente avec le reste de la population (exemple 1 : prise de poids de 8 kg en 15 jours ; exemple 2 : perte de poids au bout de 15 jours puis mort de l’animal).
Le modèle statistique : Le modèle mixte est utilisé dans cette étude afin de prendre en compte les mesures de pesée répétées pour chaque animal et l’effet individuel de chaque chevreau
(Henderson, 1985). Le modèle mixte comporte à la fois des effets fixes qui sont les facteurs de variation de la production laitière et des effets aléatoires qui sont le père et la mère du chevreau.
Les poids à âge type et les gains moyens quotidiens ont été calculés selon un modèle linéaire mixte (R² = 0.972) puis un modèle quadratique mixte (R² = 0.989). Les poids à âge type de 10 à 90 jours ont été calculés à partir de trois coefficients du modèle quadratique qui sont : le poids à la naissance corrigé, le gain moyen quotidien et l’âge de l’animal au carré. Les résultats du modèle quadratique ont été sauvegardés pour le reste de l’analyse car les valeurs estimées sont plus proches des valeurs mesurées (Figure 1). La variation des poids des chevreaux en fonction de l’âge
à la pesée n’est pas un effet linéaire. La corrélation entre le poids à la naissance du chevreau mesuré et le poids à la naissance corrigé par le modèle quadratique est forte (R² = 0.84) – (Figure 2). Le poids à la naissance corrigé surestime légèrement le poids à la naissance mesuré de 40 grammes.
La variable réponse du modèle mixte est représentée par les intervalles de gain moyen quotidien 10-30 (modèle 1) et 30-70 (modèle 2). Les effets fixes utilisés dans ces deux modèles mixtes sont :
10
le rang de mise-bas, le sexe des chevreaux, la taille de portée en cours d’allaitement, le lot de lutte (année et saison de lutte) et le poids du chevreau à la naissance mesuré. Deux effets aléatoires ont été incorporé dans les modèles mixtes : l’effet du père (car la conduite d’élevage de Gardel repose sur des familles de père) et l’effet mère intra père.
Le modèle mixte va comparer les modalités des diverses variables à un individu de base. Celui-ci sera représenté par la première modalité de chacune des variables. Dans notre cas d’étude, les chevreaux de la base de données seront comparés à un chevreau mâle né d’une portée simple par une primipare ayant été mise à la reproduction en saison sèche durant l’année 2000.
La validation des effets fixes a été réalisée à l’aide de la fonction drop1 sur le logiciel R. Cette fonction a permis de tester l’effet global du modèle comportant tous les effets fixes. Tous les effets fixes, initialement mis dans le modèle mixte, ont un effet significatif sur le GMQ ainsi les cinq variables explicatives sont conservées dans le modèle mixte. L’analyse statistique des données a été réalisée à partir d’une analyse de variance multifactorielle sur le logiciel R.
Calcul des paramètres génétiques : Plusieurs paramètres génétiques ont été calculés à partir du logiciel SAS et VCE. Le modèle utilisé est un modèle animal. Dans l’analyse de variance, on considère les performances de l’individu uniquement et non celles de ses apparentés afin d’estimer précisément la valeur génétique des chevreaux (Bonaiti et al, 1990). Les effets fixes sont identiques à ceux utilisés dans le modèle mixte précédemment. Les effets aléatoires utilisés sont l’effet animal, l’effet mère et l’effet de l’environnement permanent maternel. Les paramètres génétiques d’intérêt calculés sont la variabilité génétique et l’héritabilité additive, maternelle et de l’environnement permanent maternel, ainsi que la covariance et la corrélation génétique animal / mère.
Calcul du progrès génétique : Les résultats concernant le calcul du progrès génétique seront disponibles prochainement.
2. Les résultats
2.1.
Estimation des facteurs de variation environnementaux
Après une correction de tous les facteurs de variation enregistrés (rang de mise-bas, taille de portée, sexe du chevreau, lutte, poids de naissance), le gain moyen quotidien de 10 à 30 jours (GMQ 10-30) d’un chevreau est de 84 (+/- 2.3) g/j et le gain moyen quotidien de 30 à 70 jours (GMQ 30-70) est de 74 (+/- 2.2) g/j.
11
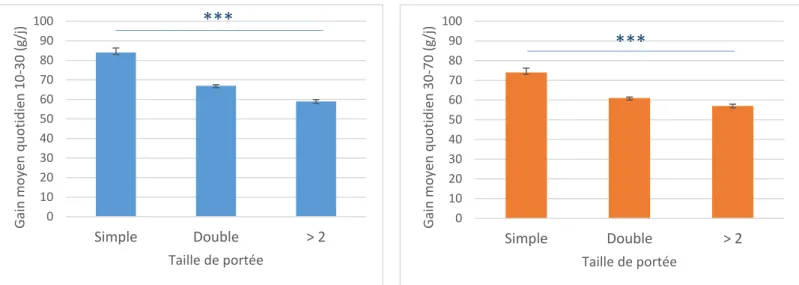
Test ANOVA 3 voies p<0.001 : *** avec la condition taille de portée simple
Figure 3 : Variation du gain de poids des chevreaux selon la taille de portée
Test ANOVA 3 voies p<0.001 : ***
Figure 4 : Variation du gain de poids des chevreaux mâles et femelles
Test ANOVA 3 voies p<0.001 : ***, p<0.01 : ** avec la condition rang de mise-bas 1 à 2
Figure 5 : Variation du gain de poids des chevreaux selon le rang de mise-bas de la
chèvre
0 10 20 30 40 50 60 70 80 90 100 Simple Double > 2 G ain moy en q u o tid ie n 30 -70 (g/ j) Taille de portée***
0 10 20 30 40 50 60 70 80 90 100 Simple Double > 2 G ain moy en q u o tid ie n 10 -30 (g/ j) Taille de portée***
0 10 20 30 40 50 60 70 80 90 100 Mâle Femelle G ain moy en q u o tid ie n 30 -70 (g/ j) 0 10 20 30 40 50 60 70 80 90 100 Mâle Femelle G ain moy en q u o tid ie n 10 -30 (g/ j)***
***
0 20 40 60 80 100 1 à 2 3 à 7 8 à 10 > 10 G ain moy en q u o tid ie n 30 -70 (g/ j) Rang de mise-bas 0 20 40 60 80 100 1 à 2 3 à 7 8 à 10 > 10 G ain moy en q u o tid ie n 10 -30 (g/ j) Rang de mise-bas***
***
***
***
***
**
12
La taille de portée : La taille de portée au cours de l’allaitement a un effet significatif sur les performances de croissance des chevreaux Créole (p<0.001) – (Figure 3). Le gain moyen quotidien des chevreaux diminue lorsque la taille de portée augmente. En considérant tous les effets du modèle, les chevreaux nés double ont un GMQ 10-30 de 67 g/j soit une valeur plus faible de 17 g/j que les chevreaux nés simple. Les chevreaux nés dans une portée de trois chevreaux minimum ont un GMQ 10-30 de 59 g/j (-25 g/j par rapport à un chevreau né simple). Les GMQ 30-70 sont plus faibles pour tous les chevreaux nés d’une portée simple ou multiple. Les différences de GMQ entre le GMQ 10-30 et le GMQ 30-70 vont de 2 à 10 g/j.
La prolificité moyenne des chèvres Créole de Gardel est de 2.25 chevreaux par portée. Plus de 60 % des naissances se sont réalisées avec des portées doubles. Également, plus de 70 % des chevreaux ont été allaités en double et 69 % des chevreaux sevrés ont été élevés en double sous la mère. La taille de portée à la naissance maximale enregistrée est de 7 chevreaux. Les naissances multiples (de 3 à 7 chevreaux par portée) représentent 30 % des naissances totales sur le domaine de Gardel. La productivité numérique moyenne des chèvres Créole de l’exploitation est de 2.79 chevreaux par femelle par an.
Le lot : L’année et la saison de lutte, regroupées dans la variable « Lutte », ont un effet significatif sur la croissance des chevreaux (p<0.001).
La parité : Le rang de mise-bas de la chèvre est un second facteur faisant varier le gain de poids des chevreaux (p<0.001) – (Figure 4). Les chevreaux ayant le gain moyen quotidien le plus élevé en période d’allaitement (GMQ 10-30 : 91g/j, GMQ 30-70 : 78g/j) sont élevés par des chèvres avec un rang de mise-bas compris entre 3 et 7 (p<0.001). En début d’allaitement, les chevreaux nés d’une chèvre ayant un rang de mise-bas compris entre 3 et 10 ont un gain moyen quotidien identique (GMQ 10-30 : 91 g/j). Ces chevreaux ont un gain de poids significativement plus élevé que les chevreaux nés de jeunes chèvres (p<0.001). Les chèvres dites exceptionnelles représentent les femelles ayant réalisées plus de 10 mises bas. Le gain de poids des chevreaux nés de chèvres exceptionnelles ne varie pas significativement de celui des chevreaux nés de jeunes chèvres durant toute la période d’allaitement. Les primipares élèvent des chevreaux ayant une croissance plus lente que les multipares ayant réalisées au minimum 3 lactations. La longévité de la chèvre favorise la croissance des chevreaux.
Le sexe des chevreaux : Parmi les variables étudiées dans le modèle, le sexe des chevreaux est un facteur qui fait varier significativement le poids et le gain de poids des chevreaux (p<0.001) –
(Figure 5). La répartition des chevreaux (où le sexe a été enregistré) est équitable entre les mâles (52%) et les femelles (48%). Les chevreaux mâles ont un GMQ 10-30 supérieur de 5 g/j par rapport aux femelles. De 30 à 70 jours, les mâles ont un GMQ supérieur de 7 g/j par rapport aux chevreaux femelles.
13
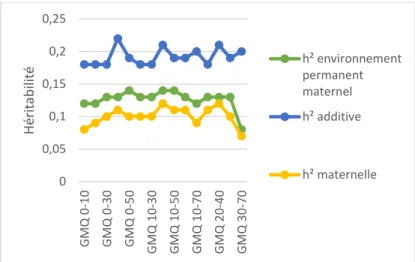
Figure 6 : Evolution des héritabilités selon l'intervalle de gain moyen quotidien
Figure 7 : Evolution des héritabilités selon les poids à âge type
0 0,05 0,1 0,15 0,2 0,25 G MQ 0-10 G MQ 0-30 G MQ 0-50 G M Q 10 -30 G MQ 10-50 G MQ 10-70 G MQ 20-40 G MQ 30-70 H érita b ili té h² environnement permanent maternel h² additive h² maternelle 0 0,05 0,1 0,15 0,2 0,25 PAT 10 PAT 20 PAT 30 PAT 40 PAT 50 H érita b ili té h² environnement permanent maternel h² additive h² maternelle14
2.2.
Choix du meilleur critère de croissance pour l’évaluation indirecte de la
production laitière
Les héritabilités additive et maternelle ont été calculées par le logiciel VCE pour plusieurs intervalles de GMQ (Figure 6) ainsi que pour des poids à âge type allant de 10 à 50 jours (Figure 7). Dans le but d’une évaluation indirecte de la production laitière, l’objectif de ces calculs de paramètres génétiques est de rechercher une variable ayant le plus de variabilité maternelle.
Le GMQ 10-40 est l’intervalle de GMQ présentant le plus de variabilité maternelle. Pour le GMQ 10-40, l’héritabilité additive est de 0.21 (+/-0.018), l’héritabilité maternelle est de 0.12 (+/- 0.016) et l’héritabilité de l’environnement permanent maternel est de 0.14 (+/- 0.010). Pour le poids âge type à 40 jours, les héritabilités sont de 0.22 (+/- 0.018), 0.11 (+/- 0.016) et 0.13 (+/-0.010) respectivement. Ces deux dernières héritabilités sont qualifiées de faible car elles sont inférieures à 0.20. L’héritabilité additive, qui peut être qualifiée de moyenne, est deux fois plus élevée que l’héritabilité maternelle. La corrélation génétique animal / mère est négative et est la plus forte pour ces deux variables.
2.3.
Discussion
Les performances de croissance des chevreaux sont influencées par divers effets individuels et maternels. Les travaux d’INRAE Antilles-Guyane réalisés en 1997 sur la croissance des chevreaux Créole reflètent les mêmes résultats que ceux présentés dans cette étude. Les chevreaux naissant d’une portée multiple ont une croissance plus faible en période d’allaitement (Benyoucef et Ayachi, 1991 ; Alexandre et al, 1997 ; Alexandre et al, 1999). La prolificité moyenne des chèvres Créole de Gardel (2.25 chevreaux / portée) est identique aux résultats trouvés dans l’étude d’Alexandre et al en 1999 et est supérieur à d’autres races caprines allaitantes. La chèvre allaitante de Réunion a une
prolificité de 1.67 chevreaux / portée (Fontaine et Choisis, 2006). Les chèvres ayant réalisées trois mises bas ou plus ont un développement corporel plus important, ce que favorise la prolificité et ainsi la production laitière est plus élevée (Alexandre et al, 1997). Les primipares produisent des chevreaux ayant une croissance plus faible que les multipares et les chèvres exceptionnelles. Conformément aux articles cités précédemment, les chevreaux mâles ont un gain de poids supérieur par rapport aux femelles en période d’allaitement. Enfin la saison de lutte ou la saison de naissance influencera les performances des chevreaux. En saison humide, les poids à âge type ainsi que les GMQ en période d’allaitement sont plus faibles qu’en saison sèche ou en saison intermédiaire du fait de l’augmentation du parasitisme (Alexandre et al, 1997). Toutes ces mêmes variables ont été étudiées sur les paramètres de croissance des chevreaux de Réunion et ont révélé un effet significatif sur leurs GMQ en période d’allaitement (Fontaine et Choisis, 2006).
15
En filière ovine, l’estimation de la valeur laitière est réalisée à partir de la variabilité maternelle calculée par des mesures de performances des agneaux à 30 jours (PAT 30) et sur le gain de poids en début d’allaitement avec le GMQ 10-30 (Idele, 2003). Les héritabilités additive et maternelle calculées pour la chèvre Créole sont plus faibles qu’en ovin allaitant (Menissier et al, 1991). Le sevrage des chevreaux Créole, qui sont des animaux de plus petit format, se réalise plus tardivement que les autres petits ruminants. Les choix réalisés dans cette l’étude sur les critères GMQ 10-40 et PAT40 sont cohérents avec les besoins physiologiques de la race. La période d’estimation de la production laitière serait alors plus étendue en chèvre Créole qu’en ovin allaitant. Le nombre de descendants par père moyen est de 34 chevreaux (min/max : 1 à 150 chevreaux) et le nombre de descendants par mère est de 7 chevreaux (min/max : 5 à 37 chevreaux). Le nombre de descendants par père et par mère est suffisamment élevé pour valider la cohérence des résultats et la faisabilité d’une sélection de la race Créole sur la production laitière.
Conclusion
Les travaux relatifs à l’effet de la production laitière de la chèvre sur la croissance des chevreaux sont peu nombreux pour les races allaitantes et/ou tropicales. Après une analyse des performances des animaux, la chèvre Créole est un animal qui est bien adapté aux systèmes d’élevage semi-intensif, utilisant le pâturage, en Guadeloupe. Elle présente de bonnes performances de reproduction en termes de longévité, de prolificité et de productivité. Cependant, les performances de croissance des chevreaux restent faibles et dépendant de plusieurs facteurs liés à l’environnement.
Malgré une héritabilité faible à moyenne des caractères, les paramètres de croissance choisis dans cette étude, qui sont le GMQ 10-40 et le PAT40, présentent une variabilité génétique exploitable chez les caprins Créole pour permettre une sélection de la race sur des critères laitiers. Le choix de ces paramètres devra également prendre en compte la facilité d’organisation du contrôle de performances en élevage.
L’URZ a su formé une base de données, répertoriant des performances et un pedigree, ce qui permettra de soutenir le développement de la population caprine Créole aux Antilles. Ces données constituent ainsi un référentiel important sur les chèvres allaitantes et la croissance des chevreaux dans ce contexte agricole particulier.
16
Bibliographie
Alexandre G, Aumont G, Fleury J, Mainaud JC, Kandassamy T. Performances zootechniques de la chèvre Créole allaitante de Guadeloupe. Bilan de 20 ans dans un élevage expérimental de l'INRA.
INRA Productions animales, 1997, 10 (1), p.7-20.
Alexandre G, Aumont G, Mainaud JC, FleuryJ, Naves M. Productive performances of Guadeloupean Creole goats during the suckling period. Small Ruminant Research, 1999. 34, p.155-160.
Alexandre G, Borel H, Matheron G, Remy C. Élevages caprins en Guadeloupe. Revue Élev. Méd.
vét. Pays trop. 1991 (n° spécial), p.27-39.
Alexandre G et Mandonnet, N. Goat meat production in harsh environments. Small Ruminant
Research, 2005, 60(1-2), p.53–66.
Bonaiti B, Boichard D, Verrier E, Ducrocq V, Barbat A, Briend M. La méthode française d’évaluation génétique des reproducteurs laitiers. INRA Productions Animales, 1990, 3 (2), p.83-92.
Chemineau P, Cognie Y, Xande A, Peroux F, Alexandre G, Levy F, Shitalou E, Beche JM, Sergent D, Camus E, Barre N, Thimonler J. Le « Cabrit créole » de Guadeloupe et ses caractéristiques zootechniques : monographie. Rev. Elev. Méd. Vét. Pays Trop., 1984, 37 (2), p.225-238.
Chemineau P, Mahieu M, Varo H, Shitalou E, Jego Y, Grude A, Thimonier J. Reproduction des caprins et ovins Créole de Guadeloupe et de Martinique. Revue Elev. Med. Vét. Pays trop. 1991 (numero special), p.45-50.
FassionD. Enquête cheptel caprins 2016 Le cheptel caprin se stabilise. Agreste Guadeloupe n° 8 mars 2017.
Fontaine O, Choisis JP. Performances zootechniques en élevage caprin allaitant - Analyse de la base de données de l’EDE. Cirad, 2006, p.1-13
Gunia M, Mandonnet N, Arquet R, de la Chevrotie C, Naves M, Mahieu M et Alexandre G. Production systems of Creole goat and their implications for a breeding programme. Animal. 2010, 4(12), p. 2099–2105.
Le Neindre P, Dubroeucq H. Observations sur l’estimation de la production laitière des vaches allaitantes par la pesée du veau avant et après la tétée. Annales de zootechnie, INRA/EDP
Sciences, 1973, 22 (4), p.413-422.
Ménissier F, Sapa J et Polvey JP. Les qualités maternelles des ruminants a viande : exemple des facilités de vêlage et de l’allaitement. Éléments de génétique quantitative et application aux
populations animales, INRA. 1991, p.1-6
Naves M, Alexandre G, Mahieu M, Gourdine J.L, Mandonnet N. Les races animales locales : bases du développement innovant et durable de l’élevage aux Antilles. Innovations Agronomiques, 2011, 16, p.193-205.
Poujardieu B. Recherche d’une méthode d’estimation de la production laitière des femelles ovines et bovines pendant la phase d’allaitement. Annales de zootechnie, INRA/EDP Sciences, 1969, 18 (3), p.299-315.
Ricordeau G, Boccard R, Denamur R. Mesure de la production laitière des brebis pendant la période d’allaitement. Annales de zootechnie, INRA/EDP Sciences, 1960, 9 (2), p.97-120.
Henderson CR, Best Linear Unbiased Prediction of Nonadditive Genetic Merits in Noninbred Populations, Journal of Animal Science, 1985, 18 (3), p. 111–117.
17
Annexes
Annexe 1 : Script réalisé sur le logiciel R # importation des 3 fichiers
alltot<-read.csv(file.choose(), header=TRUE, sep="", na.strings=".")
inventaire<-read.csv(file.choose(), header=TRUE, sep="", na.strings=".")
genealogie<-read.csv(file.choose(), header=TRUE, sep="", na.strings=".")
summary(alltot)
summary(inventaire)
summary(genealogie)
summary(fusion2)
str(fusion2)
# mettre en caractere qualitatif/quantitatif
alltot$Sexe=factor(alltot$Sexe) alltot$Num=factor(alltot$Num) alltot$TauxAllait=factor(alltot$TauxAllait) genealogie$Num=factor(genealogie$Num) genealogie$RangMB=factor(genealogie$RangMB) genealogie$Portee=factor(genealogie$Portee) fusion2$Sexe=factor(fusion2$Sexe) fusion2$year=factor(fusion2$year) fusion2$PAT90=as.numeric(fusion2$PAT90) fusion2$Saison=factor(fusion2$Saison)
# mettre les poids en kilo
alltot$Poids0<-alltot$Poids0/10
alltot$PoidsX<-alltot$PoidsX/10
inventaire$Poids0<-inventaire$Poids0/10
genealogie$Poids0<-genealogie$Poids0/10
fusion2$Poids0<-fusion2$Poids0/10
# mettre en date (jour/mois/annee)
alltot$Date0<-as.character(alltot$Date0) str(alltot) vecteur1<-vector(mode="numeric",lengt=nrow(alltot)) for(i in 1:nrow(alltot)[1]) { ifelse(nchar(as.character(alltot$Date0[i]))<4,vecteur1[i]<-2000,vecteur1[i]<-as.numeric(substring(alltot$Date0[i],nchar(as.character(alltot$Date0[i]))-4,nchar(as.character(alltot$Date0[i]))-3)))
18 } str(vecteur1) vecteur2<-vector(mode="numeric",lengt=nrow(alltot)) for(i in 1:nrow(alltot)[1]) { ifelse(vecteur1[i]<50,vecteur2[i]<-vecteur1[i]+2000,ifelse(vecteur1[i]<100,vecteur2[i]<-vecteur1[i]+1900,vecteur2[i]<-2000)) } str(vecteur2) d1<-as.numeric(substring(alltot$Date0,nchar(alltot$Date0)-2,nchar(as.character(alltot$Date0)))) d2<-paste(vecteur2,"-01-01",sep="") d3<-as.Date(d2) d4<-julian(d3,origin=as.Date("1900-01-01"))+d1 alltot$D0<-as.Date(d4,origin="1900-01-01") # alltot$DateX<-as.character(alltot$DateX) str(alltot) vecteur3<-vector(mode="numeric",lengt=nrow(alltot)) for(i in 1:nrow(alltot)[1]) { ifelse(nchar(as.character(alltot$DateX[i]))<4,vecteur1[i]<-2000,vecteur3[i]<-as.numeric(substring(alltot$DateX[i],nchar(as.character(alltot$DateX[i]))-4,nchar(as.character(alltot$DateX[i]))-3))) } str(vecteur3) vecteur4<-vector(mode="numeric",lengt=nrow(alltot)) for(i in 1:nrow(alltot)[1]) { ifelse(vecteur3[i]<50,vecteur4[i]<-vecteur3[i]+2000,ifelse(vecteur3[i]<100,vecteur4[i]<-vecteur3[i]+1900,vecteur4[i]<-2000)) } str(vecteur4) d5<-as.numeric(substring(alltot$DateX,nchar(alltot$DateX)-2,nchar(as.character(alltot$DateX)))) d6<-paste(vecteur4,"-01-01",sep="") d7<-as.Date(d6) d8<-julian(d7,origin=as.Date("1900-01-01"))+d5 alltot$DX<-as.Date(d8,origin="1900-01-01") #fonction remove rm(d1,d2,d3,d4,d5,d6,d7,d8,vecteur1,vecteur2,vecteur3,vecteur4,i,year,tt,tt2)
19
# mettre date en age
install.packages(c("lubridate", "magrittr")) library("lubridate") library("magrittr") alltot$age<-time_length(interval(alltot$D0,alltot$DX),"days") str(age) summary(age) alltot$Age<-Age[,-14]
# enlever les valeurs abérrantes
alltot<-alltot[alltot$age>=0,]
alltot<-alltot[alltot$age<120,]
summary(alltot)
range(alltot$age)
# mettre des valeurs uniques
alltot<-unique(alltot)
str(alltot)
# minimal et maximal
range(alltot$age)
# mettre un chiffre par ligne
rownames(alltot)<-1:nrow(alltot)
str(alltot2)
# trier par numero et par age
alltot<-alltot[order(alltot$Num,alltot$age),]
# calculer l'age maximum des chevreaux
alltot$agemax<-0 for(i in levels(alltot$Num)) { alltot[alltot$Num==i,"agemax"]<-max(alltot[alltot$Num==i,"age"]) } summary(alltot) AgeMax<-alltot[alltot$agemax>30,] rownames(AgeMax)<-1:nrow(AgeMax) str(AgeMax) summary(AgeMax)
20 range(alltot$agemax) fusion2$agemax<-fusion2[fusion2$agemax>30,] # data.frame de alltot alltot<-data.frame(alltot$Num,alltot$D0,alltot$Poids0,alltot$TauxNaiss,alltot$TauxAllait,alltot$TauxSevr,alltot$DX,alltot$PoidsX, alltot$age,alltot$agemax) names(alltot)=c("Num","Date0","Poids0","TauxNaiss","TauxAllait","TauxSevr","DateX","PoidsX","age","agemax") alltot2<-unique(alltot[,c(1:6,10)]) # residus d'alltot alltot$resid<-resid(fmi) bbx2<-boxplot(resid(fmi)) alltot2<-alltot[alltot$resid>=bbx2$stats[5],] alltot2<-alltot[alltot$resid>=bbx2$stats[1],]
# calcul du GMQ lineaire naissance - sevrage
ADG<-as.data.frame(coeffmi$Num) names(ADG)<-c("intercept","GMQ") ADG$Num<-row.names(ADG) str(ADG) row.names(ADG)<-seq(1:nrow(ADG)) summary(ADG) names(ADG)=c("intercept","GMQSevr","Num")
# calcul du GMQ quadratique naissance - sevrage
fusion$age2<-fusion$age^2 fmi2<-lmer(PoidsX~age+age2+(age+age2|Num),fusion,REML=F) coeffmi2<-coef(fmi2) ADG2<-as.data.frame(coeffmi2$Num) ADG2$Num<-row.names(ADG2) row.names(ADG2)<-seq(1:nrow(ADG2)) names(ADG2)=c("Num","intercept2","GMQSevr2","age22") str(ADG2) summary(ADG2) # merge fusion<-merge(ADG,alltot,by.x="Num",by.y="Num", all=FALSE,all.x=TRUE,sort=TRUE) fusion<-merge(fusion,genealogie,by.x="Num",by.y="Num",all=FALSE,all.x=TRUE,sort=TRUE)
21 fusion<-data.frame(fusion$Num,fusion$Mere,fusion$Pere, fusion$Date0,fusion$Poids0,fusion$TauxNaiss,fusion$TauxAllait,fusion$TauxSevr,fusion$Sexe.y,fusion$DateX,fusion$P oidsX,fusion$age,fusion$agemax,fusion$intercept,fusion$GMQSevr,fusion$Lutte,fusion$RangMB,fusion$Portee) names(fusion)=c("Num","Mere","Pere","Date0","Poids0","TauxNaiss","TauxAllait","TauxSevr","Sexe","DateX","PoidsX","a ge","agemax","intercept","GMQSevr","Lutte","RangMB","Portee") merge(fusion,ADG2,by.x="Num",by.y="Num", all=FALSE,all.x=TRUE,sort=TRUE)
fusion2<-merge(ADG2,alltot2, by="Num", all=FALSE,all.x=TRUE,sort=TRUE)
fusion2<-merge(fusion2,genealogie,by="Num",all=FALSE,all.x=TRUE,sort=TRUE) fusion2<-data.frame(fusion2$Num,fusion2$Mere,fusion2$Pere,fusion2$Date0,fusion2$Poids0,fusion2$TauxNaiss,fusion2$TauxAll ait,fusion2$TauxSevr,fusion2$Sexe,fusion2$Lutte,fusion2$RangMB,fusion2$agemax,fusion2$intercept2,fusion2$GMQ2, fusion2$age22) names(fusion2)<-c("Num","Mere","Pere","Date0","Poids0","TauxNaiss","TauxAllait","TauxSevr","Sexe","Lutte","RangMB","agemax","interc ept2","GMQ2","age22") summary(fusion2)
# regression avec des donnees repetees mixte + calcul du R²
require(lme4) fmi<-lmer(PoidsX~age+(age|Num),alltot,REML=F) summary(lm(fitted(fmi)~alltot$PoidsX)) # R² = 0.9724 summary(fmi) str(fmi) coeffmi<-coef(fmi) str(coeffmi)
# fitted avec le fichier 1 (fusion)
fusion$age2<-fusion$age^2 summary(lm(fitted(fmi)~fusion$PoidsX)) fmi2<-lmer(PoidsX~age+age2+(age+age2|Num),fusion,REML=F) summary(lm(fitted(fmi2)~fusion$PoidsX)) # R²=0.989 summary(fmi2) alltot$age2<-alltot$age^2 fmi2<-lmer(PoidsX~age+age2+(age+age2|Num),alltot,REML=F) summary(lm(fitted(fmi2)~alltot$PoidsX)) # R²=0.989 summary(fmi2) tt<-0:100 tt2<-tt^2
22 lines(tt,fmi@beta[1]+(tt*fmi@beta[2]),col="blue") # lineaire # extraire l'annee install.packages(c("lubridate", "magrittr")) library("lubridate") library("magrittr") fusion2$year<-year(fusion2$Date0)
# extraire le mois (effet saison)
fusion2$month<-month(fusion2$Date0)
fusion2$Saison<-ifelse(fusion2$month<5,"Dry",ifelse(fusion2$month<9,"Interm","Rain"))
# calcul du PAT linéaire
fusion$PAT10<-(fusion$GMQSevr*10)+fusion$intercept fusion$PAT20<-(fusion$GMQSevr*20)+fusion$intercept fusion$PAT30<-(fusion$GMQSevr*30)+fusion$intercept PAT40<-data.frame(fusion$Num,fusion$intercept,fusion$GMQSevr,fusion$agemax) names(PAT40)=c("Num","intercept","GMQSevr","agemax") PAT40<-PAT40[PAT40$agemax>=40,] range(PAT40$agemax) PAT40$PAT40<-(PAT40$GMQSevr)*40+PAT40$intercept PAT40$intercept<-NULL PAT40$GMQSevr<-NULL PAT40$agemax<-NULL fusion<-merge(fusion,PAT40,by.x="Num",by.y="Num", all=FALSE,all.x=TRUE,sort=TRUE) names(fusion)=c("Num","Mere","Pere","Date0","Poids0","TauxNaiss","TauxAllait","TauxSevr","Sexe","DateX","PoidsX","a ge","agemax","intercept","GMQSevr","Lutte","RangMB","Portee","year","SaisonSeche","SaisonHumide","PAT10","PAT20 ","PAT30","PAT40")
# calcul du PAT quadratique
fusion2$PAT10<-round((fusion2$GMQ2*10)+(fusion2$age22*10*10)+fusion2$intercept2,2) fusion2$PAT20<-round((fusion2$GMQ2*20)+(fusion2$age22*20*20)+fusion2$intercept2,2) fusion2$PAT30<-round((fusion2$GMQ2*30)+(fusion2$age22*30*30)+fusion2$intercept2,2) fusion2$PAT40<-ifelse(fusion2$agemax>=40,round((fusion2$GMQ2*40)+(fusion2$age22*40*40)+fusion2$intercept2,2),".") fusion2$PAT50<-ifelse(fusion2$agemax>=50,round((fusion2$GMQ2*50)+(fusion2$age22*50*50)+fusion2$intercept2,2),".") fusion2$PAT60<-ifelse(fusion2$agemax>=60,round((fusion2$GMQ2*60)+(fusion2$age22*60*60)+fusion2$intercept2,2),".") fusion2$PAT70<-ifelse(fusion2$agemax>=70,round((fusion2$GMQ2*70)+(fusion2$age22*70*70)+fusion2$intercept2,2),".") fusion2$PAT80<-ifelse(fusion2$agemax>=80,round((fusion2$GMQ2*80)+(fusion2$age22*80*80)+fusion2$intercept2,2),".")
23
fusion2$PAT90<-ifelse(fusion2$agemax>=90,round((fusion2$GMQ2*90)+(fusion2$age22*90*90)+fusion2$intercept2,2),".")
# correction des PAT negatif
fusion2<-fusion2[fusion2$PAT30>=0.5,]
range(fusion2$PAT30)
# calcul du PAT selon le controle de performance
fusion$PAT30bis<-alltotPAT30$Poids2+(((alltotPAT30$Poids3-alltotPAT30$Poids2)/15)*(30-15))
# calcul des intervalles de GMQ (kg/j)
fusion2$GMQ0.10<-round(((fusion2$PAT10-fusion2$intercept2)/10),3)*1000 fusion2$GMQ0.20<-round(((fusion2$PAT20-fusion2$intercept2)/20),3)*1000 fusion2$GMQ0.30<-round(((fusion2$PAT30-fusion2$intercept2)/30),3)*1000 fusion2$GMQ0.40<-round(((fusion2$PAT40-fusion2$intercept2)/40),3)*1000 fusion2$GMQ0.50<-round(((fusion2$PAT50-fusion2$intercept2)/50),3)*1000 fusion2$GMQ10.20<-round(((fusion2$PAT20-fusion2$PAT10)/10),3)*1000 fusion2$GMQ10.30<-round(((fusion2$PAT30-fusion2$PAT10)/20),3)*1000 fusion2$GMQ10.40<-round(((fusion2$PAT40-fusion2$PAT10)/30),3)*1000 fusion2$GMQ10.50<-round(((fusion2$PAT50-fusion2$PAT10)/40),3)*1000 fusion2$GMQ10.60<-round(((fusion2$PAT60-fusion2$PAT10)/50),3)*1000 fusion2$GMQ10.70<-round(((fusion2$PAT70-fusion2$PAT10)/60),3)*1000 fusion2$GMQ20.30<-round(((fusion2$PAT30-fusion2$PAT20)/10),3)*1000 fusion2$GMQ20.40<-round(((fusion2$PAT40-fusion2$PAT20)/20),3)*1000 fusion2$GMQ20.50<-round(((fusion2$PAT50-fusion2$PAT20)/30),3)*1000 fusion2$GMQ30.70<-round(((fusion2$PAT70-fusion2$PAT30)/40),3)*1000
# correlation entre poids de naissance et poids de naissance corrigé
plot(intercept2~Poids0,fusion2,xlim=c(0,5),ylim=c(0,5),ylab="Poids à la naissance corrigé (kg)",xlab="Poids à la naissance (kg)", main="Corrélation entre le poids à la naissance mesurée et le poids à la naissance corrigé par le modèle quadratique")
summary(lm(intercept2~Poids0,fusion2)) # r²=0.84
abline(lm(intercept2~Poids0,fusion2),col="blue")
abline(h=0)
# plot
plot(alltot$age,alltot$PoidsX,ylim=c(0,20),pch=".",xlab="âge (jours)",ylab="Poids (kg)",main="Evolution du poids des chevreaux de la naissance au sevrage")
plot(fusion2$PAT30~fusion2$RangMB,type="c=l",pch=".",xlab="Rang de mise-bas",ylab="PAT30",main="Poids des chevreaux selon la parité de la chèvre")
24
# supprimer une colonne
fusion2$month<-NULL
# moyenne, ecartype, variance
mean(fusion2$PAT40)
sd(fusion2$GMQ2)
var(fusion2$PAT30)
skimr::skim(fusion2)
# distribution des variables a +/- 2 ecart-types
fusion2$Poids0Sdd<-fusion2$Poids0<=1.30
fusion2$PAT10Sdd<-fusion2$PAT10<=1.6
fusion2$PAT30Sdd<-fusion2$PAT30<=2.3
fusion2$PAT50Sdd<-fusion2$PAT50<=3.3
fusion2$PAT70Sdd<-fusion2$PAT70<=3.8
# identification des chevreaux avec un GMQ 10-30 negatif
fusion2$GMQnegatif<-fusion2$GMQ10.30<0
nrow(fusion2[fusion2$GMQ10.30<0,])
row.names(fusion2[fusion2$GMQ10.30<0,])
# supprimer une ligne par le numero du chevreau
fusion3=fusion2 fusion2<-fusion2[fusion2[,1]!="2017405",] # boxplot boxplot(fusion2$GMQ10.30~fusion2$Sexe,data=fusion2) boxplot(fusion2$GMQ10.30~fusion2$RangMB,data=fusion2) boxplot(fusion2$GMQ10.30~fusion2$TauxNaiss,data=fusion2)
# nombre de descendants moyens par pere et par mere
# 14053/416 = 34 chevreaux par pere
# 14053/2118 = 7 chevreaux par mere
# nombre de descendants total par pere et par mere
summary(fusion2$Pere,maxsum=422) # de 1 à 150 chevreaux / pere
summary(fusion2$Mere,maxsum=2200) # de 5 à 37 chevreaux / mere
# mettre une mere et un pere aleatoire
25
fusion2$Perebis<-sample(fusion2$Pere,nrow(fusion2),replace=TRUE)
fusion2$Merebis["NA"]<-sample(fusion2$Mere,nrow(fusion2),replace=TRUE)
# regrouper les donnees des variables TauxAllait et RangMB
fusion2$TauxAllait<-ifelse(fusion2$TauxAllait==1,1,ifelse(fusion2$TauxAllait==2,2,"3et+")) fusion2$TauxAllait<-factor(fusion2$TauxAllait) fusion2$RangMB1[fusion2$RangMB==1]<-"1-2" fusion2$RangMB1[fusion2$RangMB==2]<-"1-2" fusion2$RangMB1[fusion2$RangMB==3]<-"3-7" fusion2$RangMB1[fusion2$RangMB==4]<-"3-7" fusion2$RangMB1[fusion2$RangMB==5]<-"3-7" fusion2$RangMB1[fusion2$RangMB==6]<-"3-7" fusion2$RangMB1[fusion2$RangMB==7]<-"3-7" fusion2$RangMB1[fusion2$RangMB==8]<-"8-10" fusion2$RangMB1[fusion2$RangMB==9]<-"8-10" fusion2$RangMB1[fusion2$RangMB==10]<-"8-10" fusion2$RangMB1[fusion2$RangMB==11]<-"11et+" fusion2$RangMB1[fusion2$RangMB==12]<-"11et+" fusion2$RangMB1[fusion2$RangMB==13]<-"11et+" fusion2$RangMB1[fusion2$RangMB==14]<-"11et+" fusion2$RangMB1[fusion2$RangMB==15]<-"11et+" fusion2$RangMB1[fusion2$RangMB==16]<-"11et+" fusion2$RangMB1[fusion2$RangMB==17]<-"11et+" fusion2$RangMB1<-factor(fusion2$RangMB1) # exporter write.table(fusion2,file="ChevreCreole_Coralie.csv",quote=FALSE,sep=";",na=".",row.names=FALSE,col.names=TRUE) ################## ANALYSE DE VARIANCE ############################### # analyse de variance require(lme4) require(lmerTest) mod0 = lmer(GMQ10.30~RangMB+Lutte+Sexe+Poids0+TauxAllait+(1|Mere:Pere)+(1|Pere),fusion2,REML=F) anova(mod0) summary(mod0) coef(mod0) str(coef(mod0)) summary(coef(mod0)) vcov(mod0)
26 mod0=lm(GMQ10.30~RangMB+year+Saison+Sexe+Poids0+TauxAllait,fusion2) drop1(mod0,test='F') plot(mod0) tab1<-as.data.frame(fitted(mod0)) plot(fitted(mod0)~fusion2$GMQ10.30) mod1 = lmer(GMQ30.70~RangMB+Lutte+Sexe+Poids0+TauxAllait+(1|Mere:Pere)+(1|Pere),fusion2,REML=FALSE) anova(mod1) summary(mod1) mod2 = lmer(GMQ10.40~RangMB+Lutte+Sexe+Poids0+TauxAllait+(1|Mere:Pere)+(1|Pere),fusion2,REML=FALSE) anova(mod2) summary(mod2)
# analyse des effets individuels
mod2=lm(GMQ10.30~RangMB,data=fusion2)
anova(mod2)
summary(mod2) # ***
# analyse de variance avec interaction
library(FactoMineR) AovSum(PAT30~GMQ2+Sexe+GMQ2:Sexe,data=fusion2) # ** AovSum(PAT30~GMQ2+Portee+GMQ2:Portee,data=fusion2) # *** AovSum(PAT30~GMQ2+RangMB+GMQ2:RangMB,data=fusion2) AovSum(PAT30~GMQ2+Sexe+Poids0+Portee+TauxAllait,data=fusion2) # *** AovSum(PAT30~GMQ2+Poids0+GMQ2:Poids0,data=fusion2) # *** AovSum(PAT30~Pere+Poids0+Pere:Poids0,data=fusion2)
# analyse de variance avec la fonction Anova 2 facteurs
require(car) mood<-lm(PAT30~GMQSevr*Sexe,data=fusion) Anova(mood) # *** summary(mood) # test de Student t.test(PAT30~Sexe, var.equal=TRUE,data=fusion2)
# test d'un effet lineaire
fit=fitted(fusion2)
plot(fusion2$PoidsX~fusion2$age,pch=".",xlab="Age (jours)",ylab="Poids (kg)", main="Poids du chevreau de la naissance au sevrage")
mod=lm(PoidsX~age,data=fusion2)
abline(mod,col="blue")
boxplot(fusion2$PoidsX~fusion2$age,pch=".",xlab="Age (jours)",ylab="Poids (kg)", main="Poids du chevreau de la naissance au sevrage")
27
Résumé
Les chèvres Créole de Guadeloupe sont élevées pour la viande en système allaitant. L’étude porte sur l’estimation indirecte de la production laitière des chèvres via les performances de croissance des chevreaux. Les mesures de performances sont réalisées depuis 1980 au sein de la station expérimentale de Gardel d’INRAE Antilles-Guyane. Le modèle mixte utilisé comprend des effets fixes (rang de mise-bas, taille de portée, sexe des chevreaux, lutte, poids à la naissance) et des effets aléatoires (mère, père). Toutes les variables étudiées ont un effet significatif sur le GMQ 10-30 et 30-70 des chevreaux. Après une mesure de la variabilité additive et maternelle de plusieurs paramètres de croissance, le meilleur critère de croissance permettant l’estimation indirecte de la production laitière en chèvre Créole est le GMQ 10-40 et le PAT40. La sélection d’une souche de chèvre Créole sélectionnée sur sa production laitière est faisable dans le troupeau expérimental d’INRAE-PTEA.
MOTS CLEFS : chèvre allaitante, production laitière, croissance
Abstract
The Creole goats of Guadeloupe are raised for meat in a suckling system. The study focuses on the indirect estimation of the milk production of the goats with the growth performance of the kids. Performance measurements have been carried out since 1980 at the Gardel experimental station of INRAE Antilles-Guyane. The mixed model used includes fixed effects (farrowing rank, litter size, sex of the kids, group, birth weight) and random effects (mother, father). All the variables studied had a significant effect on the DWG 10-30 and 30-70 of the kids. After measuring the additive and maternal variability of several growth parameters, the best growth criterion allowing the indirect estimation of milk production in Creole goats is the DWG 10-40 and the LW40. The selection of a strain of Creole goat selected on its milk production is feasible in the INRAE-PTEA experimental herd.