HAL Id: hal-02809940
https://hal.inrae.fr/hal-02809940
Submitted on 6 Jun 2020HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Etude des dynamiques de populations de pucerons ailés
en cultures de melon : vers la compréhension et la
gestion des épidémies virales
Laure Douste Bacque
To cite this version:
Laure Douste Bacque. Etude des dynamiques de populations de pucerons ailés en cultures de melon : vers la compréhension et la gestion des épidémies virales. Sciences agricoles. 2013. �hal-02809940�
Etude des dynamiques de populations de pucerons ailés en cultures de melon : vers la compréhension et la gestion des épidémies virales de DOUSTE-BACQUE Laure est mis à disposition selon les termes de la licence Creative Commons Attribution - Pas d’Utilisation Commerciale - Pas de Modification 3.0 non transposé
Ministère de l'Agriculture, de l'Agroalimentaire et de la Forêt
ECOLE NATIONALE SUPERIEURE des SCIENCES AGRONOMIQUES de BORDEAUX AQUITAINE
1, cours du Général de Gaulle – CS 40201 – 33175 GRADIGNAN cedex
M E M O I R E d e f i n d ' é t u d e s p o u r l ' o b t e n t i o n d u t i t r e
d ' I n g é n i e u r d e B o r d e a u x S c i e n c e s A g r o
Etude des dynamiques de populations de pucerons ailés en
cultures de melon : vers la compréhension et la gestion des
épidémies virales
Douste-Bacqué Laure
Option : AGROGER - Agro-écologie et Gestion des ressources
Etude réalisée à : unité de Pathologie Végétale, INRA PACA, Avignon
Ministère de l'Agriculture, de l'Agroalimentaire et de la Forêt
ECOLE NATIONALE SUPERIEURE des SCIENCES AGRONOMIQUES de BORDEAUX AQUITAINE
1, cours du Général de Gaulle – CS 40201 – 33175 GRADIGNAN cedex
M E M O I R E d e f i n d ' étu d es p o u r l ' o b t e n t i o n d u t i t r e
d'In gén ieu r d e B o rdea ux Scie nce s A gro
Etude des dynamiques de populations de pucerons ailés en
cultures de melon : vers la compréhension et la gestion des
épidémies virales
Douste-Bacqué Laure
Maître de stage : Schoeny Alexandra
Option : AGROGER - Agro-écologie et Gestion des ressources
Etude réalisée à : unité de Pathologie Végétale, INRA PACA, Avignon
PREFACE
L’INRA, Institut National de la Recherche Agronomique, est un organisme de recherche scientifique fondé en 1946 dans le contexte de reconstruction nationale de l’après-guerre. Il est placé sous la double tutelle du ministère délégué de l’Enseignement Supérieur et à la Recherche et du ministère de l’Agriculture et de la Pêche, dispose d’un budget annuel de 881.6 millions d’euros et compte 8478 agents (données 2011).
Aujourd'hui, ses recherches concernent trois domaines fortement imbriqués : l'alimentation, l'agriculture et l'environnement avec l'ambition de développer une agriculture à la fois compétitive, respectueuse de l'environnement, des territoires et des ressources naturelles, et mieux adaptée aux besoins nutritionnels de l'homme ainsi qu'aux nouvelles utilisations des produits agricoles.
L'INRA s’impose comme un acteur majeur de la recherche agronomique dans le monde. En nombre de citations, il maintient sa 2e position mondiale dans le domaine de l’agriculture et se place au 4e
rang mondial en sciences des plantes et de l’animal. L’Institut figure en outre parmi les premiers organismes mondiaux en microbiologie, en écologie et en environnement.
Le centre Inra Provence-Alpes-Côte d'Azur est un centre de recherche tourné vers l’agroécologie des systèmes de culture sous serres et en vergers et la modélisation de l’impact régionalisé du changement climatique à l’échelle du paysage. Il rassemble 1000 agents répartis dans 26 unités, localisés sur 10 sites. Les recherches menées à l’unité de Pathologie Végétale, située à Avignon, sont essentiellement focalisées sur les maladies bactériennes, fongiques et virales des fruits et légumes du bassin méditerranéen. L’objectif est le développement de méthodes de lutte efficaces et raisonnées dans un contexte de production agronomique durable et de haute qualité.
REMERCIEMENTS
Je tiens tout d’abord à remercier Cindy Morris, directrice de l’unité de Pathologie Végétale, de m’avoir accueillie comme stagiaire.
Je remercie Alexandra Schoeny, ma maître de stage, chargée de recherches, pour m’avoir fait découvrir l’entomologie, pour ses conseils et le temps qu’elle m’a consacré, notamment lors de la rédaction de ce mémoire, et pour la confiance qu’elle m’a accordé tout au long de mon stage.
Je tiens à remercier également Hervé Lecoq, directeur de recherches, de m’avoir fourni certaines données d’épidémiologie virale et fait partager un peu de son savoir et de son expérience en virologie.
Je remercie Jonathan Gaudin, assistant ingénieur, pour son implication dans ce projet, son aide dans l’identification des pucerons et sa bonne humeur quotidienne. Je n’oublie pas Thomas Costes et Vanina Monteil, stagiaires, pour leur soutien et leur gentillesse.
Enfin, je tiens à remercier l’ensemble du personnel du site Saint Maurice pour leur accueil chaleureux et leur disponibilité pendant la durée de mon stage.
Pour finir, je remercie les étudiants de l’option AGROGER avec qui j’ai partagé de nombreux projets, ainsi que notre responsable de formation, Lionel Jordan-Meille, pour avoir contribué à rendre cette dernière année de formation enrichissante dans une ambiance conviviale.
Ce stage a bénéficié du soutien financier du Comité Technique Permanent de la Sélection (projet Parcel-R) et de l’Agence Nationale de la Recherche (projet VirAphid).
SOMMAIRE
INTRODUCTION
... 1
SYNTHESE BIBLIOGRAPHIQUE
... 3
1. Le cas du melon dans le Sud-Est………..3
2. Les maladies virales du melon………. 3
2.1 Les propriétés des virus 3 2.2 Transmission par les pucerons 4 2.3 Les principales maladies virales du melon 4 2.3.1 CMV : Cucumber Mosaic Virus 4
2.3.2 WMV : Watermelon Mosaic Virus 5
2.3.3 ZYMV : Zucchini Yellow Mosaic Virus 6
2.3.4 CABYV: Cucurbit Aphid-Borne Yellows Virus 7
3. Les pucerons, vecteurs de virus………8
3.1 Les pucerons, des Hémiptères 8 3.2 Les pucerons, des ravageurs de cultures 8 3.3 Les pucerons, des vecteurs de phytovirus 9 3.4 Aphis gossypii, Aphis craccivora et Myzus persicae 9 4. Les caractéristiques de la relation puceron-virus………. 11
4.1 la spécificité et l’efficacité de la combinaison virus-vecteur 11 4.2 Epidémiologie et Modélisation 12 5. Les méthodes de lutte contre les virus du melon……….. 13
5.1 Actions sur l’environnement de la culture 13 5.2 Actions sur la culture elle-même 13 6. La lutte génétique : le gène Vat chez le melon………. 14
6.1 Le gène Vat 15 6.2 La durabilité du gène Vat 15 7. L’aménagement de l’environnement parcellaire……….. 16
7.1 Les avantages des associations de cultures 16 7.2 L’aménagement de l’environnement parcellaire contre les pucerons et les virus 17 7.3 Les bandes enherbées et fleuries 18 7.3.1 Critères de choix des espèces végétales utilisées 18
MATERIELS ET METHODES
... 21
1. Dispositif expérimental de l’essai Parcel-R……… 21
1.2 Caractéristiques du dispositif 22 1.3 Composition et gestion des aménagements parcellaires 22 1.4 Données météorologiques 23 2. Suivi des dynamiques de populations de pucerons ailés……….. 24
2.1 Le système de piégeage 24 2.2 Traitement des prélèvements et identification morphologique 24 2.3 Identification par barcode ADN 25 3. Suivi des épidémies virales……….. 26
3.1 Plan de prélèvements 26 3.2 Test DAS-ELISA 26 4. Traitements des données………. 27
4.1 Ajustement des épidémies virales et calcul d’AUDPC 27 4.2 Tests statistiques non paramétriques 29 RESULTATS
... 30
1. Dynamiques de populations de pucerons ailés sur l’essai Parcel-R……… 30
1.1 Traitement des données de l’essai 2013 30 1.1.1 Composition globale des prélèvements 30
1.1.2 Identification des espèces de pucerons piégées 33
1.2 Comparaison interannuelle des piégeages 2011-2013 34 1.2.1 Composition globale des prélèvements 34
1.2.2 Identification des espèces de pucerons piégés 35
2. Etude des épidémies virales sur l’essai Parcel-R………. 37
2.1 Epidémies virales de la saison 2013 37 2.2 Comparaison interannuelle des épidémies virales 2011-2013 38
3. Recherche de liens épidémies virales-pucerons………. 39
3.1 Observation des tendances 40 3.2 Prévision des épidémies à partir du nombre de pucerons capturés 81 DISCUSSION
... 43
1. Effets de l’aménagement parcellaire sur les populations de pucerons ailés………... 43
2. Effets du gène Vat et de l’aménagement parcellaire sur les épidémies virales…………. 44
3. Un protocole opératoire avec ses avantages et ses inconvénients……… 46
4. La relation virus – pucerons……… 47
CONCLUSION ET PERSPECTIVES
... 49
REFERENCES BIBLIOGRAPHIQUES ... 51
TABLES DES FIGURES ET DES TABLEAUX
Table des Figures
Figure 1: Représentation schématique de la transmission d'un virus par un puceron. ... 4
Figure 2: Symptômes du CMV sur feuille de melon: tâches chlorotiques ... 5
Figure 3: Symptômes du WMV sur feuille de melon: mosaïques ... 6
Figure 4: Symptômes du ZYMV sur feuille de melon: boursouflures et cloques vert foncé .... 6
Figure 5: Symptômes du CABYV sur feuille de melon: jaunissement des feuilles âgées ... 7
Figure 6: Pucerons A. Gossypii aptères ... 19
Figure 1: Puceron A. Craccivora ailé ………10
Figure 8: Puceron ailé M. Persicae et larves ... 19
Figure 9: Localisation de la parcelle expérimentale au sein du site de Saint Paul ... 21
Figure 10: Schéma du dispositif expérimental Parcel-R ... 22
Figure 11: Les cinq espèces composant la bande fleurie………...…… ...… 23
Figure 12: Evaluation du pourcentage de couverture du sol le 24 mai 2013, jour de la plantation des melons. ……….….…23
Figure 13 : Suivi de la floraison des différentes espèces composant les bandes fleuries pendant la durée de la saison culturale. ………..…….23
Figure 14: Piège à succion permettant la capture journalière de pucerons. ... 23
Figure 15 : Schéma descriptif des différents organes d’un puceron ailé permettant son identification……… 24
Figure 16: Schématisation des différentes étapes du test DAS-ELISA ... 25
Figure 17 :Représentation graphique des nombres de pucerons et de non pucerons piégés quotidiennement sur la modalité SN sur l’essai Parcel-R 2013 et volume des précipitations. 27 Figure 18 :Représentation graphique des nombres de pucerons et de non pucerons piégés quotidiennement sur la modalité SN sur l’essai Parcel-R 2013 et vitesse moyenne du vent supérieure à 8km/h. ... 30
Figure 19 :Superposition des courbes d’effectifs journaliers de pucerons sur les trois modalités ... 31
Figure 20 : Superposition des courbes d’effectifs de pucerons journaliers sur la modalité SN pour les trois années ... 32
Figure 21: Evolution du nombre de plantes infectées par le CMV (a), le WMV (b) et le CABYV (c) au cours de la saison culturale 2013 et AUDPC totale (J0 à J56) correspondantes (d, e, f). ………..…………..35
Figure 22: Superposition des courbes d’incidence des différentes épidémies, observées et ajustées par le modèle de Gompertz, pour chaque virus et pendant les trois années sur la modalité SNS ……….………..37 Figure 23 : AUDPC totale (J0 à J56) moyenne des différentes épidémies virales des trois années sur les différentes modalités………..……38 Figure 24 : Représentation graphique des effectifs hebdomadaires de pucerons et des
pourcentages de plantes nouvellement infectées par les différents virus sur l’essai Parcel-R 2013 ... 40 Figure 25 : AUDPC totale des épidémies de CABYV en fonction du nombre d’a. Gossypii de J0 à J14 ... 42
Table des Tableaux
Tableau 1: Effectifs totaux de pucerons et de non pucerons piégés pendant toute la saison culturale, sur les trois modalités et proportions moyennes de pucerons dans les captures ... 61 Tableau 2: Effectifs de pucerons piégés pendant les 21 premiers jours et parts relatives des principales espèces de pucerons pour chaque modalité ... 33 Tableau 3: Effectifs totaux de pucerons et de non pucerons piégés pendant toute la saison culturale, sur les trois modalités pendant les trois années ... 34 Tableau 4 : Effectifs de pucerons piégés pendant les 21 premiers jours et parts relatives des principales espèces de pucerons pour chaque modalité et toutes les années ... 36 Tableau 5 : Etude de l’effet du couplage des deux méthodes de lutte : comparaison des
AUDPC totales moyennes sur les modalités BER et BEF par rapport au témoin SNS ... 38 Tableau 6: Calcul du coefficient de corrélation entre les variables : pourcentage de plantes nouvellement infectées par semaine et le nombre de pucerons piégés les 14 jours précédant les détections de virus sur ces plantes ... 81 Tableau 7: Calcul des coefficients de corrélation entre les variables AUDPC totale (J56) et le cumul de différentes espèces de pucerons de J0 à J14 pour les différents virus
... 42
LISTE DES ABREVIATIONS
ADN: acide désoxyribonucléique
AUDPC: area under the disease progress curve BE: modalité bande enherbée
BER: modalité avec un aménagement bande enherbée et des melons possédant le gène Vat BES: modalité avec un aménagement bande enherbée et des melons ne possédant pas le gène
Vat
BF: modalité bande fleurie
BFR: modalité avec un aménagement bande fleurie et des melons possédant le gène Vat BFS: modalité avec un aménagement bande fleurie et des melons ne possédant pas le gène
Vat
CABYV: Cucurbit Aphid Borne Yellow Virus, virus de la jaunisse des cucurbitacées transmis par puceron
CMV: Concumber Mosaic Virus, virus de la mosaïque du concombre
CVMV : Chilli Veinal Mottle Virus, virus de la marbrure des nervures du piment GAFL: Génétique et Amélioration des Fruits et Légumes, unité INRA
GRAB: Groupe de Recherche en Agriculture Biologique INRA: Institut National de la Recherche Agronomique M10 : essai Melon 2010
P11, P12 et P13 : essais Parcel-R 2011, 2012 et 2013
PLRV: Potato LeafRoll Virus, virus d’enroulement des feuilles de la pomme de terre PVMV: Pepper Veinal Mottle Virus, virus de la marbrure des nervures du poivron PYV: Potato Yellow Virus: virus de la jaunisse de la pomme de terre
SAS: Statistical Analysis System SMV: Soybean Mosaic Virus SN: modalité Sol Nu
SNR : modalité sans aménagement et dont les melons sont pourvus du gène Vat SNS: modalité sans aménagement et dont les melons ne possèdent pas le gène Vat V11, V12 et V13 : essais VirAphid 2011, 2012 et 2013
WMV: Watermelon Mosaic Virus, virus de la mosaïque du concombre
GLOSSAIRE
Acellulaire : qui n'est pas composé de cellules, qui n’est pas vivant.
Anholocyclique : qualifie une espèce d’insecte dont le cycle est incomplet : décrit les espèces de pucerons ayant perdu totalement ou partiellement la possibilité de se reproduire par la voie sexuée et se reproduisant toute l’année par parthénogénèse.
Antibiose : toute relation biologique dans laquelle un être vivant en détruit un autre pour assurer sa propre existence.
Antixénose : toute réaction biologique dans laquelle un être vivant repousse ou amoindrit les dommages causés par les ravageurs qui l’attaquent.
Aphidien : adjectif qualifiant les pucerons.
Bioagresseur : tout organisme ayant un effet négatif sur le développement des cultures (ravageurs, maladies, mauvaises herbes…).
Callose : polymère de glucose synthétisé par les cellules végétales pour stopper le passage de la sève et d’éléments extérieurs (bactéries, champignons…)
Contournant : individu qui est parvenu à passer au-delà d’un phénomène de résistance. Dioécique : se dit d’une espèce de pucerons dont les générations se développent sur des plantes hôtes différentes au cours de leur cycle biologique.
Fitness : capacité d’un individu à se reproduire.
Holocyclique : qualifie une espèce d’insecte dont le cycle est complet : décrit les espèces de pucerons qui alternent reproduction parthénogénétique et reproduction sexuée.
Hôte primaire : plante sur laquelle un insecte effectue sa reproduction sexuée et dépose son œuf d’hiver.
Hôte alternatif : plante, non propice à la reproduction sexuée, sur laquelle la forme ailé d’un insecte a émigré et où il se reproduit par parthénogénèse exclusivement.
Phytovirus : virus s’attaquant aux organismes végétaux.
Parthénogénèse : reproduction sans fécondation dans une espèce non vertébrée, sexuée. Polyphage : qui mange des substances variées.
1
INTRODUCTION
La plupart des efforts de l’agriculture moderne ont été consacrés à l’augmentation de la quantité et de la qualité de la production agricole. Cependant, très souvent, les maladies des plantes interfèrent avec cet objectif. La France produit chaque année près de 300 000 tonnes de melon, la plaçant au 3e rang des pays producteurs européens. Cette culture est soumise à de nombreux ravageurs dont les principaux sont les pucerons et les virus qu’ils transmettent. Les méthodes de lutte contre ces bioagresseurs ne sont pas totalement efficaces et les producteurs de melons sont à la recherche de moyens de lutte innovants, à la fois économes et faciles à mettre en place.
Les gènes de résistance semblent être une alternative intéressante pour contrôler les ravageurs des cultures tout en réduisant l’utilisation des produits phytosanitaires, répondant ainsi aux exigences réglementaires et aux attentes des agriculteurs et des consommateurs.
La limite majeure de la résistance génétique vient des possibilités d’adaptation des bioagresseurs : les contournements de résistance sont fréquents. Pour faire face à cette difficulté, de nouvelles stratégies permettant de gérer durablement les résistances doivent être envisagées.
C’est dans ce contexte que le Ministère de l'agriculture et de la pêche soutient le projet Parcel-R (2011-2014), qui regroupe des équipes de l'INRA Avignon (Pathologie Végétale et Génétique et Amélioration des Fruits et Légumes), le Groupe de Recherche en Agriculture Biologique (Avignon) et le Centre d'Expérimentation des Fruits et Légumes (Montauban). Ce projet étudie la possibilité d’utiliser l’aménagement de l’environnement parcellaire pour renforcer l'efficacité et la durabilité de la lutte génétique aux pucerons.
Le dispositif expérimental conçu pour ce projet permet de tester l’effet de trois types d’aménagement (sol nu, bandes enherbées et bandes fleuries) sur l’efficacité et la durabilité de la résistance apportée par le gène Vat. L’efficacité est évaluée par l’étude de la dynamique des populations de pucerons visiteurs (ailés) et colonisateurs (aptères), l’étude de l’entomofaune auxiliaire et l’étude des épidémies virales. La durabilité est évaluée par la recherche d’éventuels pucerons contournants et l’analyse de leur fitness.
L’étude des dynamiques de populations de pucerons ailés, arrivant et circulant dans la culture, est indispensable à la compréhension des épidémies virales et à la conception de systèmes de culture permettant de les limiter.
2
Cette étude se concentre sur le suivi des populations de pucerons ailés ainsi que le suivi des épidémies virales, afin d’étudier l’efficacité du couplage « gène de résistance + aménagement parcellaire » et d’initier la recherche des relations liant dynamiques de populations de pucerons ailés et épidémies virales.
3
SYNTHESE BIBLIOGRAPHIQUE
1. Le cas du melon dans le Sud-Est
Le melon est une plante annuelle rampante de la famille des Cucurbitacées, cultivée pour ses fruits. Il est produit en France dans trois principaux bassins : Sud-Est, Sud-Ouest et Poitou-Charentes.
La France est le 3e producteur européen (13%), après l’Espagne (44%) et l’Italie (28%). En 2012, 280 211 tonnes de melons ont été produites en France, dont 43% dans le Sud-Est (Agreste, 2012).
Le melon est cultivé sous serres (5%), sous abris bas (50%) ou en plein air (45%), préférentiellement dans un sol profond à structure aérée. Sa culture s’étend de mi-mars à août selon le mode de production et le climat de la région considérée. Le zéro végétatif du melon se situe à 12°C, sa température de croissance optimale est de l’ordre de 25°C. Son besoin total en eau est de 300 à 400 mm (Guerineau, 1998).
Dans le Sud-Est, cette culture est majoritaire et est soumise à de nombreuses maladies fongiques : l’oïdium, la pourriture grise, la fusariose, le mildiou… Mais les principaux ravageurs du melon sont les pucerons : leurs piqûres et les nombreuses maladies virales qu’ils transmettent sont fortement préjudiciables pour les cultures (Guerineau, 1998).
2. Les maladies virales du melon
2.1 Les propriétés des virus
Les virus sont des particules acellulaires microscopiques infectieuses. Ils sont composés d'une ou plusieurs molécules d'acide nucléique (ADN, ARN, simple ou double brin), entourées d'une coque de protéines appelée capside, et parfois d'une enveloppe. Ces parasites obligatoires utilisent la machinerie cellulaire de leur hôte pour se multiplier. Les virus sont en général pathogènes : la présence d’un phytovirus dans une plante hôte perturbe ses processus physiologiques pouvant entraîner jusqu’à sa mort. Les symptômes se manifestent par des altérations de la couleur, des déformations tissulaires et des nécroses (Astier et al., 2001). En ce qui concerne la production végétale agricole, ces perturbations peuvent conduire à d’importantes pertes de rendement et/ou à une altération de la qualité organoleptique des produits les rendant non-commercialisables.
4 2.2 Transmission par les pucerons
La transmission des virus se fait généralement par l’intermédiaire d’un vecteur, souvent un arthropode qui, en se nourrissant au dépend de la plante, crée une faille physique, permettant l’intrusion de la particule virale dans l’hôte. Les pucerons sont les principaux vecteurs de phytovirus (Astier et al., 2001).
Les virus peuvent être classés en différents groupes en fonction de leur interaction avec le vecteur qui les transporte. Un virus peut être circulant ou non circulant dans le vecteur, selon le temps, le site de rétention et le
cheminement qu’il fait à l’intérieur même du vecteur. Alors que les virus non circulants sont retenus au niveau des pièces buccales, sans pénétrer à l’intérieur du vecteur (Figure 1a), les virus circulants sont transportés dans l’hémolymphe et finissent par rejoindre les glandes salivaires (Figure 1b). Parmi les phytovirus, la plupart sont non-circulants, ils sont dits non-persistants ou semi-persistants en fonction de la durée pendant laquelle le vecteur reste virulent après s’être nourri sur une plante infectée (de quelques secondes à quelques minutes). Lorsque le vecteur reste virulent plus longtemps (de quelques heures à plusieurs jours) les virus sont dits persistants (Nault, 1997 ; Astier, 2001 ; Ng et Perry, 2004).
2.3 Les principales maladies virales du melon 2.3.1 CMV : Cucumber Mosaic Virus
Le virus de la mosaïque du concombre (CMV) a été isolé pour la première fois en 1916 aux USA. Ce Cucumovirus est présent sur tous les continents et sous tous les climats, son incidence peut varier d’une région à l’autre. En France, la fréquence des infections varie beaucoup en fonction de l’espèce, de l’année ou de la région.
Figure 1: représentation schématique de la transmission d'un virus par un puceron. (Astier et al, 2001)
a : acquisition d’un virus non persistant lors de brèves piqûres d’épreuve dans les tissus superficiels. b : acquisition d’un virus persistant lors d’une piqûre d’alimentation dans le phloème et circulation dans l’organisme du vecteur.
5
Les symptômes s’expriment sous forme de quelques taches chlorotiques sur la feuille inoculée, ils se généralisent puis évoluent en mosaïque (figure 2). Les fruits présentent également des mosaïques plus ou moins marquées. En cas d’infection précoce, la production peut être fortement compromise (E-phytia, 2013).
Ce virus est transmis de manière non persistante par plus d’une soixantaine d’espèces de pucerons dont Aphis gossypii, Myzus persicae, Aphis craccivora (Jacquemont, 2012).
Le CMV possède l’une des plus vastes gammes d’hôtes, tant chez les monocotylédones que chez les dicotylédones. Plus de 1200 espèces végétales sont sensibles à ce virus, notamment de nombreuses espèces maraichères (tomate, salade, piment…) et fruitières (cerisier, bananier…) (Lecoq & Desbiez, 2012).
2.3.2 WMV : Watermelon Mosaic Virus
Le virus de la mosaïque de la pastèque (WMV) appartient à la famille des Potyvirus. Il est particulièrement présent dans les zones aux climats tempérés et méditerranéens. Observé pour la première fois en 1954 en Floride, et en 1974 en France, ce virus est aujourd’hui présent dans toutes les zones de production de melon du monde. Il est très fréquent en Europe, et est le virus le plus fréquent chez les cucurbitacées en France (Encyclop’aphid, 2012).
Une grande diversité de symptômes a été observée en fonction de la souche du virus et du cultivar de la plante hôte. Sur le melon, apparaissent sur les feuilles des mosaïques plus ou moins importantes accompagnées d’une coloration vert foncé des nervures et une crispation du feuillage (figure 3). Lorsque les symptômes sont sévères, la taille des feuilles est drastiquement réduite et lesfruits sont décolorés et déformés (Lecoq & Desbiez, 2012).
Figure 2: symptômes du CMV sur feuille de melon: tâches chlorotiques (Source : E-phytia, Blancard, 2013)
6
Au moins 35 espèces de pucerons, dont Aphis gossypii, Myzus persicae et Aphis
craccivora, sont capables de transmettre ce virus non persistant (Lecoq & Desbiez, 2008).
Une grande efficacité de la transmission par ces vecteurs serait à l’origine de la propagation très rapide de la maladie sans qu’une grande quantité de pucerons soit présente.
Le virus compte 170 plantes hôtes dont certaines pérennes peuvent lui servir d’hôtes alternatifs notamment l’hiver.
2.3.3 ZYMV : Zucchini Yellow Mosaic Virus
Le virus de la mosaïque jaune de la courgette (ZYMV) est un Potyvirus apparu en Italie en 1973 puis en 1979 en France. Il se retrouve aujourd’hui dans pratiquement toutes les zones de production du monde. Il engendre des épidémies irrégulières en France mais sévères. Une attaque précoce peut provoquer une perte totale des récoltes ou de spectaculaires déformations des fruits les rendant non commercialisables.
Les symptômes sur melon sont un éclaircissement des nervures des feuilles, un jaunissement, l’apparition de boursouflures et de cloques vert foncé ainsi qu’un arrêt de croissance (figure 4). Les entre-nœuds restent courts donnant un aspect de buisson au melon. Si le melon possède le gène Fn, le ZYMV entraîne un flétrissement très rapide des plants. Sur les fruits sont observés des mosaïques, des boursouflures, des éclatements et parfois la déformation des graines.
Figure 3: symptômes du WMV sur feuille de melon: mosaïques (Source : E-phytia, Lecoq, 2013)
Figure 4: symptômes du ZYMV sur feuille de melon: boursouflures et cloques vert foncé (Source : E-phytia, Blancard, 2013)
7
Au moins 26 espèces de pucerons sont capables de transmettre ce virus, notamment
Aphis gossypii, Myzus persicae et Aphis craccivora (Katis et al., 2006). De plus, Aphis
gossypii vit plus longtemps et produit plus de descendance, notamment plus d’individus ailés,
sur un plant infecté par le ZYMV que sur un plant non infecté (Blua & Perring, 1992). Ce phénomène expliquerait la rapide dispersion du virus.
Sa gamme d’hôtes est limitée aux cucurbitacées, à quelques plantes d’ornement et mauvaises herbes.
2.3.4 CABYV: Cucurbit Aphid-Borne Yellows Virus
Le virus de la jaunisse des cucurbitacées transmise par les pucerons (CABYV) est un Polerovirus, décrit en 1989 en France, mais sûrement présent auparavant sans avoir été identifié. Ses symptômes peuvent être confondus avec ceux de carences minérales. Il est aujourd’hui présent dans la plupart des pays où l’on cultive des cucurbitacées, excepté dans les Antilles et une partie de l’Amérique du Sud (Lecoq, 1999).
Ce virus n’est pas d’une gravité importante en France, il peut toutefois affaiblir les plantes et entraîner des réductions significatives du nombre de fruits produits, sans altérer leur qualité (Lecoq & Desbiez, 2012).
Les symptômes de ce virus sont le développement de plages internervaires jaunes sur les feuilles âgées, puis le jaunissement s’étend progressivement et affecte plusieurs étages foliaires à la base des plantes (figure 5). Les feuilles âgées s’épaississent et deviennent cassantes (Lecoq et al., 1992).
Les plantes hôtes de ce virus sont les cucurbitacées et de nombreuses plantes adventices. Le CABYV est transmis selon le mode persistant. Il faut pour l’acquérir que le puceron atteigne le phloème, tissus conducteur où circule la sève élaborée dont il se nourrit. Ces piqures d’alimentation sont beaucoup plus longues que les piqûres d’essai.
Figure 5: symptômes du CABYV sur feuille de melon: jaunissement des feuilles âgées (Source : E-phytia, Blancard, 2013)
8
Le virus effectue un cycle dans le corps du puceron et il finira par s’accumuler dans les glandes salivaires de l’insecte le rendant virulent. C’est lors d’une nouvelle piqûre d’alimentation sur une plante saine que le puceron pourra transmettre le virus (Lecoq, 1992). Ce cycle s’effectue sans multiplication virale au sein du vecteur, mais implique des interactions entre le virus et le puceron expliquant la spécificité de vection du CABYV : peu d’espèces de pucerons le transmettent. Aphis gossypii et Myzus persicae en sont capables.
3. Les pucerons, vecteurs de virus
3.1 Les pucerons, des Hémiptères
Les pucerons sont des insectes appartenant à l’ordre des Hémiptères, la plupart sont de la famille des Aphididae. Plus de 4000 espèces de pucerons sont répertoriées dans le monde dont 600 sont présentes en France (Dedryver et al., 2010). Parmi elles une centaine d’espèces ont une importance économique car elles se développent au détriment de plantes cultivées.
Les pucerons sont dotés de pièces buccales de type piqueur-suceur. Elles leur permettent de percer les parois végétales et d’atteindre les vaisseaux du phloème pour y prélever la sève dont ils se nourrissent. Alors qu’en général les pucerons se nourrissent sur une ou quelques plantes hôtes appartenant à la même famille, certaines espèces de pucerons sont polyphages, ils peuvent se nourrir sur une grande variété de plantes hôtes (Ng et Perry, 2004).
Les pucerons ont un fort potentiel de multiplication et de dissémination. Leur multiplication se fait essentiellement par parthénogénèse (reproduction asexuée) avec un temps de génération très court. L’alternance de formes ailées et aptères leur permet de se propager et de changer d’hôte au cours de leur cycle biologique (Astier et al., 2001 ; Simon et al., 2002).
3.2 Les pucerons, des ravageurs de cultures
Les pucerons sont à l’origine de nombreux dégâts sur les cultures (Encyclop’aphid, 2012). Ils peuvent être répartis en deux catégories : les dégâts directs et indirects.
Les dégâts directs :
Le prélèvement de sève va entraîner un affaiblissement de la plante, voir la mort pour de jeunes plants,
9
La toxicité de la salive du puceron peut causer une grande variété de symptômes sur les feuilles, les fleurs et les fruits : déformation, décoloration…les rendant non commercialisables.
Les dégâts indirects :
Les pucerons libèrent sous forme de déjections une substance riche en sucres, le miellat, qui favorise le développement de champignons saprophytes formant des fumagines grisâtres sur les feuilles et les fruits. Cette formation altère la photosynthèse et la respiration de la plante et rend les fruits non commercialisables.
Des phytovirus sont transmis par les pucerons lorsque ces derniers s’alimentent sur le végétal.
3.3 Les pucerons, des vecteurs de phytovirus
Parmi les 600 pucerons répertoriés en France, 275 sont des vecteurs de virus. La contamination du puceron par une particule virale et sa transmission à une plante ont lieu lors de la phase de nutrition de l’insecte. Son comportement alimentaire très particulier consiste à explorer et échantillonner en faisant des « piqûres d’essai » dans différents tissus végétaux. Ces piqûres très brèves, répétées et superficielles correspondent aux conditions optimales pour l’acquisition ou la transmission des virus non persistants. Si la plante goûtée lui convient, le puceron enfonce complètement ses stylets jusqu’au phloème et se nourrit longuement de la sève (Astier et al., 2001 ; Brault et al., 2010).
Les pucerons possèdent des caractéristiques telles que leur fort potentiel reproductif, leur capacité de dispersion et leur mode d’alimentation tout à fait propices à leur prolifération et qui en font des vecteurs de virus performants (Ng et Perry, 2004 ; Brault et al., 2010). Plus de 300 phytovirus sont transmis par les pucerons.
3.4 Aphis gossypii, Aphis craccivora et Myzus persicae
Ces trois espèces de pucerons sont connues pour leur présence sur culture de melon, mais également en tant que vecteurs performants des principaux virus des Cucurbitacées.
10
Aphis gossypii (figure 6), aussi appelé
puceron du cotonnier et du melon, est le principal ravageur du melon, ses piqûres entraînent une crispation des feuilles et un arrêt de croissance. Ses attaques précoces peuvent entraîner la mort des jeunes plants. Il s’agit d’une espèce anholocyclique qui se multiplie par parthénogénèse essentiellement. Les ailés assurent la dissémination d’une plante hôte à une autre. Il est très polyphage, mais a une
préférence pour les cucurbitacées (melon, concombre, courgette…), les malvacées (cotonnier, hibiscus…) et les rutacées (citrus). Il est capable de transmettre un grand nombre de virus, persistant ou non (Hullé et al., 1999 ; Blackman & Eastop, 2000).
Aphis craccivora (figure 7), aussi appelé
puceron noir de la luzerne, est une espèce de puceron essentiellement anholocyclique : les populations se maintiennent toute l’année de façon parthénogénétique. Cette espèce très polyphage se nourrit préférentiellement sur les pousses en croissance, les inflorescences et les fruits. Il a une large gamme de plantes hôtes, avec une préférence pour les fabacées comme la fève et la luzerne. En
cas de pullulations sur les tissus jeunes, la croissance et la floraison des plantes peuvent être perturbées. Les dégâts de cette espèce sont également dûs aux nombreux virus, souvent non persistants, qu’elle propage (Hullé et al., 1999 ; Blackman & Eastop, 2000).
Myzus persicae (figure 8), aussi appelé
puceron vert du pêcher, peut avoir deux cycles de vie différents. L’espèce est holocyclique dioecique, alternant entre ses hôtes primaires, les espèces du genre Prunus dont le pêcher sur lesquels a lieu la reproduction sexuée, et ses hôtes secondaires où elle se multiplie par parthénogénèse. En l’absence de son hôte primaire ou lorsque le climat est doux en hiver, le puceron du pêcher est anholocyclique et
se multiplie exclusivement par parthénogénèse sur sa très large gamme d’hôtes secondaires.
Figure 6: pucerons A. gossypii aptères (Source : Encyclop’aphid, Chaubet, 2013)
Figure 2: puceron A. craccivora ailé (Source : Encyclop’aphid, Chaubet, 2013)
Figure 3: puceron ailé M. persicae et larves (Source : Encyclop’aphid, Chaubet, 2013)
11
Elle regroupe une cinquantaine de familles botaniques dont les Solanacées, les Brassicacées et les Cucurbitacées. Cette espèce est l’une des plus dangereuses pour un très grand nombre de cultures car elle est capable de transmettre efficacement plus de 100 phytovirus (Hullé et al., 1999 ; Blackman & Eastop, 2000).
4. Les caractéristiques de la relation puceron-virus
L’épidémiologie décrit le mouvement d’une maladie virale au sein d’une population de plantes hôtes saines (Astier et al., 2001). L’analyse des processus épidémiques est importante dans le choix de moyens de lutte contre les virus. Plusieurs facteurs peuvent conditionner la dissémination d’un virus :
les caractéristiques du virus (gamme d’hôtes, abondance des sources…),
les caractéristiques du vecteur (abondance, activité…),
la relation virus-vecteur (spécificité, efficacité de la transmission…),
les conditions environnementales (température, vent...). 4.1 la spécificité et l’efficacité de la combinaison virus-vecteur
Les stratégies de transmission des virus sont variées : elles impliquent pour chaque couple virus-vecteur des interactions moléculaires spécifiques (Astier et al., 2001). Les protéines de capside, partie du virus en contact avec l’extérieur, jouent un rôle majeur dans les phénomènes de reconnaissance. L’acrostyle du puceron, zone située à l’intérieur du stylet, à son extrémité distale, serait un site de rétention des virus non circulants (Uzest et al., 2010). L’interaction virus-puceron est très spécifique concernant les virus circulants puisqu’ils doivent être capables de traverser de nombreuses barrières pour atteindre les glandes salivaires (Hogenhout et al., 2008 ; Fereres & Moreno, 2009).
La dissémination d’un virus est entièrement dépendante du comportement de l’insecte vecteur et de sa capacité à se propager de plantes en plantes. Son comportement, dans la recherche d’une plante, après qu’il se soit posé dessus ou lors des ponctions de sève, va fortement jouer sur sa capacité et son efficacité à transmettre des virus (Fereres & Moreno, 2009).
Les pucerons ont une relation spécifique avec toutes les plantes : les composés volatils qu’elles émettent vont l’attirer ou le repousser.
Les pucerons non colonisateurs de la plante sont généralement les principaux vecteurs des virus transmis au cours des piqûres d’essais (virus non persistants et semi-persistants
12
retenus sur le stylet) alors que les pucerons colonisateurs sont les principaux vecteurs des virus qui requièrent de plus longues périodes de nutrition et qui ont souvent un contact direct avec les tissus conducteurs de sève (virus persistant et quelques semi-persistants retenus dans l’intestin) (Fereres & Moreno, 2009).
Certains pucerons vont être préférentiellement attirés par des plantes infectées par un virus, favorisant sa prolifération. Certains, lorsqu’ils se nourrissent sur une plante infectée par un virus, voient augmenter leur potentiel biologique, par un accroissement naturel supérieur, et leur potentiel de migration, par la production importante d’individus ailés (Blua & Perring, 1992).
4.2 Epidémiologie et Modélisation
Une épidémie peut être divisée en deux étapes :
les contaminations primaires : elles initient l’épidémie, la source du virus peut être intrinsèque à la culture (graine contaminée) ou extérieure (adventice ou autre culture à proximité, source du virus),
les contaminations secondaires : les plantes infectées par le virus deviennent des sources de virus pour les plantes voisines.
L’analyse des courbes épidémiques permet de caractériser leur initiation (précoce/tardive) et leur vitesse de développement (rapide/lente). De plus, les épidémies virales ont souvent des allures de sigmoïdes qui peuvent être ajustées par différents modèles (logistique, Gompertz, Weibull, etc…). Les paramètres caractérisant les courbes épidémiques brutes ou lissées peuvent, dans certains cas, être expliqués par des variables liées aux vecteurs et/ou au climat. Lorsque ces relations sont établies, elles peuvent permettre la conception de modèles de simulation voire de prévision des risques épidémiques.
Ainsi pour le SMV (Soybean Mosaic Virus, non persistant) dont le principal vecteur est le puceron Aphis glycines, l’augmentation du nombre de plants de soja infectés coïncide avec l’augmentation de l’effectif de A. glycines arrivant sur la culture (Burrow et al., 2005). De même, le nombre de plants de pomme de terre infectés par le PYV (Potato Virus Y, non persistant) et le PLRV (Potato Leafroll Virus, persistant) est corrélé à l’effectif de Myzus
persicae (Ioannou, 1989).
Connaissant ces relations, le suivi des populations aphidiennes permet de prévoir les épidémies virales et de les anticiper. Par exemple, la destruction des fanes de pomme de terre
13
avant qu’un seuil, prenant en compte le nombre et l’efficacité des principaux pucerons vecteurs, ne soit atteint permettrait de limiter la prolifération du PVYN (Van Harten, 1983).
5. Les méthodes de lutte contre les virus du melon
Les plantes ne disposent pas d’un système immunitaire, une fois infectée par un virus, elles resteront malades toute leur vie. Les moyens de lutte contre les virus sont généralement préventifs. De plus, la nuisibilité d’une épidémie virale est d’autant plus importante que son initiation est précoce. Donc toute pratique permettant de retarder l’épidémie pourra avoir un effet positif sur le rendement.
5.1 Actions sur l’environnement de la culture
Les virus CWV, WMV et CABYV ne se transmettent pas par la graine. Le ZYMV peut se transmettre par la graine, des précautions doivent donc être prises lors de la sélection des semences. Les jeunes plants sains doivent le rester le plus longtemps possible. En jouant sur l’environnement de la culture, par un désherbage soigneux des abords des parcelles, on élimine les adventices servant de réservoirs à virus ou d’hôtes aux pucerons. Il est également préférable de ne pas installer côte à côte des cultures d’âges différents pour éviter que la plus âgée joue le rôle de source de virus ou de vecteurs pour la jeune plantation. Le choix de la date de plantation, en dehors des périodes de principaux vols de pucerons, est également une méthode de lutte.
Il a également été mis en évidence que les paillages naturels ou synthétiques, en plus de maintenir la température et l’humidité du sol élevées, ont un effet répulsif sur les pucerons ne leur permettant pas de distinguer la culture. Cette méthode de répulsion semble efficace jusqu’à ce que le feuillage recouvre le paillage. Grâce à cette technique, les épidémies virales (CMV, ZYMV et WMV) peuvent être retardées jusqu’à 3 à 6 semaines et les rendements sont améliorés (Stapleton & Summers, 2002). Néanmoins, elle est compliquée à mettre en place et coûteuse.
5.2 Actions sur la culture elle-même
Les principaux virus du melon étant non persistants, la durée nécessaire à leur transmission est de l’ordre de quelques secondes à quelques minutes. Les insecticides ne sont pas capables de tuer les pucerons en aussi peu de temps et ont même parfois tendance à
14
augmenter leur activité avant de les tuer (Harrewijn & Minks, 1987 ; Perring et al., 1993 ; Fereres, 2000). Ils ne sont donc pas efficaces contre la transmission de virus mais limitent les dégâts directs provoqués par la pullulation de pucerons et réduisent le risque de dissémination de virus. Néanmoins, des cas de résistance d’Aphis gossypii à plusieurs insecticides ont été observés (Furk & Hines, 1993 ; Hullé et al., 1999), le nombre d’insecticides autorisés est en diminution et ils peuvent causer des dommages collatéraux sur l’entomofaune auxiliaire (pollinisateurs…).
La pulvérisation d’huiles minérales sur les cultures a également été testée et a montré certains avantages. Elle a pour but de protéger la culture des piqûres de pucerons, et donc d’éviter la transmission de virus d’une plante infectée à une plante saine, en retardant la pénétration du stylet du puceron dans le végétal (Powell et al., 1998). Quand la pression puceron est faible, cette méthode sur cultures de melon permet la réduction de l’incidence et de la transmission de virus (CMV et WMV). Lorsqu’elle est forte, l’infection est seulement retardée de quelques jours (Umesh et al., 1995). L’utilisation d’huiles minérales nécessite de nombreuses pulvérisations et du matériel adapté pour protéger la culture tout au long de sa croissance.
Ces techniques déjà testées ne donnent pas entière satisfaction. Prises indépendamment elles ont une efficacité partielle, des avantages, mais aussi des inconvénients, elles sont notamment coûteuses et chronophages.
Un contrôle efficace des viroses peut être envisagé en associant plusieurs techniques. Le choix de celles-ci va dépendre du coût et du temps qu’elles demandent au producteur pour être mises en place. Une des techniques les plus avantageuses (simple à mettre en place et peu coûteuse) est l’utilisation de variétés de melon résistantes.
6. La lutte génétique : le gène Vat chez le melon
L’utilisation de plantes résistantes est une méthode essentielle en protection intégrée des cultures : elle permet de contrôler les populations de ravageurs tout en étant sans danger pour l’environnement et la santé humaine (Dogimont et al., 2010). Les processus permettant d’identifier les gènes responsables de la résistance et leur incorporation dans les variétés cultivées est un travail de longue haleine et plus ou moins efficace. A la fin du XXe siècle, près de 200 variétés résistantes aux insectes étaient cultivées dans le monde, dont 25% résistant aux pucerons (Dedryver et al., 2010).
15 6.1 Le gène Vat
Depuis les années 1990, des variétés de melon possédant le gène Vat, qui leur confère une résistance au puceron A. gossypii, ainsi qu’une résistance aux virus : CMV, WMV et ZYMV lorsqu’ils sont portés par A. gossypii (Hullé et al., 1999), sont cultivées en France. Ce gène est présent dans 80% des cultivars français.
Le gène Vat a trois différents effets :
L’antixénose : l’insecte n’accepte pas la plante pour s’alimenter et se reproduire même en l’absence de choix (Pitrat & Lecoq, 1982),
L’antibiose : il induit une réduction du potentiel biotique de l’insecte, sa fécondité est réduite de 80 à 90% en 3 jours (Thomas et al., 2008),
La résistance complète et spécifique à la transmission de virus non-persistants par cette espèce de puceron (Lecoq et al, 1979 ; Lecoq et al, 1980).
Comme dans la plupart des interactions plante-pathogène, la résistance serait due à la reconnaissance spécifique d’une protéine effectrice du puceron, qui déclencherait chez la plante hôte un signal activant rapidement ses défenses contre celui-ci (Dogimont et al., 2010). Le gène Vat induirait une réponse hypersensible. Lors d’une piqûre d’essai, ont lieux deux phénomènes principaux : la très rapide mise en place de dépôts de callose renforçant les parois cellulaires et la synthèse de phénols ayant un effet dissuasif sur les pucerons. Ces évènements perturberaient les activités normales de pénétration du stylet dans les cellules de la feuille et par la même occasion la transmission de virus non persistants par A. gossypii (Sarria Villada et al, 2009).
L’effet de Vat sur les épidémies virales est connu, mais peu d’articles y sont consacrés, seuls quelques résultats concernant son efficacité à retarder les épidémies de WMV ont été publiés (Lecoq & Pitrat, 1989 ; Lecoq, conversation personnelle).
6.2 La durabilité du gène Vat
La durabilité d’une résistance est la persistance de son efficacité dans le temps et dans l’espace (Johnson, 1981). Elle dépend de la capacité des populations de ravageurs à s’adapter aux variétés résistantes et de la pression de sélection exercée par les résistances sur les populations de ravageurs (Thomas, 2011).
16
Aphis gossypii a une reproduction parthénogénétique qui conduit à une faible
variabilité génétique, mais il fait preuve d’une grande plasticité phénotypique qui lui confère un fort potentiel adaptatif (Lombaert et al., 2009). Des contournements ont déjà été révélés sur le terrain, mais l’extension de ces biotypes n’a pas été observée (Dogimont et al., 2010).
Afin d’éviter un contournement rapide, il est nécessaire de renforcer la durabilité de cette résistance en associant la lutte génétique à une autre méthode de lutte intégrée. Cette méthode permettrait d’abaisser le niveau de la population aphidienne, réduisant ainsi le risque de contournement de la résistance.
7. L’aménagement de l’environnement parcellaire
7.1 Les avantages des associations de cultures
L’association de cultures consiste à cultiver plusieurs espèces végétales ou variétés sur la même parcelle et en même temps. Soit les cultures peuvent toutes être exploitées et commercialisées, soit la culture secondaire est seulement bénéfique au développement de la culture principale. Elle peut par exemple jouer un rôle protecteur face aux maladies, répulsif face aux ravageurs, ou constituer des réservoirs de biodiversité.
Dans un souci d’optimisation de l’utilisation de l’espace et de facilitation du travail de l’agriculteur, la culture secondaire est préférentiellement limitée en taille et disposée aux abords des parcelles pour ne pas gêner le passage des machines. L’idée de cultiver la bordure du champ en mettant en place une culture secondaire en tant que méthode de lutte culturale a été proposée dans les années 1950 (Fereres, 2000). L’aménagement de l’environnement parcellaire peut prendre plusieurs formes : des haies, des bandes enherbées, fleuries ou laissées en jachère…
Ces petites zones autour des cultures vont permettre de créer des habitats semi-naturels afin de lutter contre la diminution de la biodiversité et de l’abondance au sein des terres agricoles (Thomas & Marshall, 1999). En mettant à disposition, à proximité des cultures, des habitats, des refuges et des ressources alimentaires, ces zones favorisent la présence de pollinisateurs et d’ennemis naturels contribuant au contrôle biologique des ravageurs (Haaland et al., 2011 ; Pfiffner et Wyss, 2004). En effet, des parasitoïdes ou prédateurs des ravageurs des cultures peuvent y nicher et donc réduire la pression des ravageurs sur la culture principale.
17
7.2 L’aménagement de l’environnement parcellaire contre les pucerons et les virus
En plus de favoriser la présence des ennemis naturels des pucerons sur la parcelle, des hypothèses ont été formulées concernant le rôle de ces aménagements sur le contrôle des virus transmis par ces pucerons.
Hypothèse du « filtre à virus »
Pendant leur vol, les pucerons localisent les plantes hôtes grâce au contraste entre la couleur verte du feuillage et la couleur marron du sol nu : plus la couverture du sol est importante, plus la probabilité qu’un puceron se pose sur la culture est faible (Hook & Fereres, 2006). De plus, les pucerons ne peuvent distinguer les plantes hôtes des plantes non hôtes qu’après s’être posé dessus et avoir fait des piqûres d’essai sur les feuilles. Lorsque le puceron se rend compte que la plante n’est pas un hôte acceptable, il a tendance à quitter la zone, à la recherche d’une véritable plante hôte.
Ainsi, les pucerons transportant des virus non-persistants, peuvent perdre leur pouvoir virulifère lors de piqûres d’essais sur une culture secondaire à la fois non hôte pour les pucerons et les virus considérés (Fereres, 2000).
Hypothèse de la barrière physique
Si elle est assez haute, la culture secondaire placée aux abords de la parcelle peut représenter un obstacle entre le ravageur et la culture (Simons, 1957 ; Hook & Fereres, 2006). En présence de maïs ou de tournesol, les pucerons ont tendance à se poser plus sur la barrière que sur la culture d’intérêt (Simons, 1957).
Qu’il s’agisse du mécanisme du filtre à virus ou de la barrière physique, plusieurs études ont permis de mettre en évidence l’intérêt d’associer une culture principale à une culture secondaire pour lutter contre les virus transmis par les pucerons.
Une étude de Hussein & Samad (1993) a montré que les populations d’Aphis gossypii et la propagation du CVMV (Chilli Veinal Mottle Virus, virus de la marbrure des nervures du piment, virus non-persistant) sont significativement plus importantes en monoculture de piment qu’en culture associée piment/maïs ou piment/aubergine. De même, sur culture de courgettes, les plantes montrant des symptômes viraux sont significativement moins nombreuses lorsque que la culture est associée à une culture secondaire (Manandhar & Hook, 2011).
18
Une étude de Showler & Greenberg (2003) a mis en évidence que dans un champ de coton désherbé, les populations de ravageurs, dont Aphis gossypii, sont plus importantes qu’en présence de mauvaises herbes. De plus, ces dernières induisent la présence d’une plus grande population d’ennemis naturels.
7.3 Les bandes enherbées et fleuries
Contrairement à des aménagements pérennes tels que les haies, la mise en place de bandes annuelles aux abords des cultures offre une plus grande flexibilité à l’agriculteur (entretien). Dans le cadre de la lutte contre les pucerons et les virus qu’ils transmettent, le choix des espèces est très important et ne peut être laissé au hasard (jachère, végétation sauvage).
7.3.1 Critères de choix des espèces végétales utilisées
Alors que les bandes enherbées sont essentiellement constituées de graminées, les bandes fleuries ont une composition plus variée. Plus le nombre d’espèces végétales dans ces zones non-cultivées est important, plus la diversité et l’abondance des arthropodes sont grandes (Thomas, 1998).
La sélection des espèces végétales est très importante, toutes les fleurs ne conviennent pas (odeur répulsive, fleur non attractive visuellement…) (Pfiffner & Wyss, 2004). D’autres critères majeurs sont pris en compte dans le choix des espèces:
- la floraison, la plus longue possible, doit correspondre à la période de plantation et de développement de la culture,
- la présence dans la flore locale et la facilité d’approvisionnement en semences, - la nécessité d’être non hôte des pucerons ravageurs de la culture,
- la nécessité d’être non hôte des virus auxquels est sensible la culture,
- la capacité de survivre en sol agricole sans avoir tendance à envahir la culture. 7.3.2 Atouts supplémentaires des bandes fleuries
Le manque de ressources alimentaires dans les agrosystèmes peut être un obstacle pour l’efficacité des ennemis naturels à éliminer les ravageurs. La majorité des parasitoïdes, en plus de leurs proies, ont besoin de sucres (dont le nectar) pour couvrir leurs besoins énergétiques. Le nectar va augmenter leur durée de vie, leur fécondité et leur activité générale (Bianchi & Wäckers, 2008). Donc la mise en place d’une végétation complexe, riche en
19
espèces, avec une longue durée de floraison est favorable à leur installation (Pfiffner & Wyss, 2004).
La gestion de ces bandes doit se faire en fonction des objectifs établis en amont et des besoins des ennemis naturels pour se développer. Il est donc nécessaire de connaître les populations animales et végétales et leurs interactions pour mettre en place et gérer au mieux l’aménagement de l’environnement parcellaire.
20
OBJECTIFS DU TRAVAIL
L’essai Parcel-R couple deux méthodes de lutte contre les pucerons et les virus dont l’efficacité partielle a été démontrée par différentes études. Il associe l’utilisation de melons possédant le gène de résistance Vat à la mise en place d’aménagements de l’environnement parcellaire : le sol nu, la bande enherbée et la bande fleurie. Nous nous intéressons particulièrement aux effets de l’aménagement parcellaire sur les dynamiques de population de pucerons ailés ainsi que sur les épidémies virales sur melons possédant ou non le gène Vat.
La première hypothèse est que la mise en place d’un aménagement de l’environnement parcellaire favoriserait le développement de la biodiversité en créant des habitats et en apportant des sources alimentaires (nectar, pollen, proies…) à l’entomofaune auxiliaire (pollinisateurs, ennemis naturels des pucerons…). La prédation accrue, engendrée par une augmentation de l’abondance d’auxiliaires de culture, notamment des prédateurs des pucerons, tendrait à diminuer la proportion des principaux ravageurs (Aphis gossypii, Aphis
craccivora, Myzus persicae notamment) dans les piégeages et/ou modifierait la composition
de la population. Le développement de la biodiversité animale maintiendrait les ravageurs en dessous d’un seuil de nuisibilité pour la culture. Ce plus faible nombre de pucerons, confrontés à la pression exercée par la résistance génétique, diminuerait la probabilité d’apparition d’individus contournants.
La deuxième hypothèse est que la mise en place d’un aménagement de l’environnement parcellaire favoriserait la « purification » des pucerons en jouant un rôle de filtre à virus et/ou devrait stopper leur progression vers la culture par un effet de barrière physique (pour la modalité bande fleurie). Les pucerons n’arriveraient pas sur la parcelle ou arriveraient sans virus, retardant voire éliminant les infections virales.
Enfin, la connaissance de la composition des populations aphidiennes devrait permettre d’établir le lien entre la présence de certaines espèces de pucerons et l’initiation des épidémies des différents virus touchant le melon.
Les résultats permettront à terme de proposer des méthodes de lutte efficaces et durables.
21
MATERIELS ET METHODES
Cette partie met l’accent sur l’essai mené en 2013 dans le cadre du projet Parcel-R 2011-2014. Deux autres projets ont alimenté la base de données utilisée pour étudier la relation entre les épidémies virales et les dynamiques aphidiennes : Melon 2010 et VirAphid 2011-2013 (Lecoq et al, non publié ; Schoeny et al, non publié). Ils sont brièvement présentés en annexe I. La méthodologie utilisée pour établir les dynamiques de populations de pucerons ailés et les épidémies virales est la même pour tous les essais.
1. Dispositif expérimental de l’essai Parcel-R
1.1 Localisation
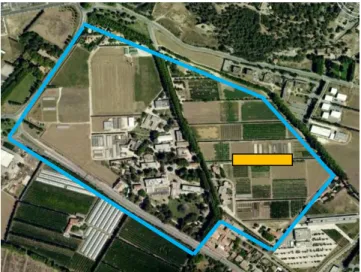
L’essai Parcel-R est implanté sur une parcelle du domaine expérimental INRA de Saint Paul à Montfavet (84140) (figure 9).
Des haies de cyprès sont implantées entre chaque parcelle expérimentale dans une orientation Est-Ouest pour protéger les cultures du vent, le mistral, qui sévit dans la région.
Domaine Saint Paul Essai Parcel-R
Figure 9: localisation de la parcelle expérimentale au sein du site de Saint Paul (Source : Google map, 2013)
22 1.2 Caractéristiques du dispositif
Deux facteurs expérimentaux sont étudiés dans cet essai :
Le type d’aménagement de l’environnement de la culture de melons, représenté par trois modalités : le sol nu (SN), les bandes enherbées (BE) et les bandes fleuries (BF).
Le type de résistance du melon à A. gossypii : soit les melons sont résistants (R) car ils possèdent le gène Vat, soit ils sont sensibles (S) car ils en sont dépourvus . Les six traitements expérimentaux qui résultent du croisement des modalités de ces deux facteurs (SNS, SNR, BES,BER, BFS et BFR) sont comparés dans le dispositif suivant (Figure 10).
Felino et Cisco sont des variétés commerciales de melon, tolérantes à l’oïdium, qui servent de zones tampons entre les différentes modalités d’aménagement parcellaire. La variété Felino possède le gène Vat, la variété Cisco en est dépourvue.
1.3 Composition et gestion des aménagements parcellaires
La modalité sol nu (SN) est régulièrement désherbée mécaniquement au cours de l’expérimentation. Cette modalité sert de témoin, elle représente les abords des champs tels qu’ils sont généralement entretenus par les agriculteurs.
La modalité bande enherbée (BE) est un semis de ray-grass anglais (Lolium perenne, famille des Poacées).
Figure 5: les cinq espèces composant la bande fleurie. De gauche à droite : bleuet, gesse, marjolaine, pimprenelle et sainfoin (Source: INRA)
Figure 12: évaluation du pourcentage de couverture du sol le 24 mai 2013, jour de la plantation des melons.
A gauche, la bande fleurie Nord a atteint 100% de couverture. A droite, la bande enherbée Nord a une couverture du sol évaluée à 90%.
(Source : photos personnelles, 2013)
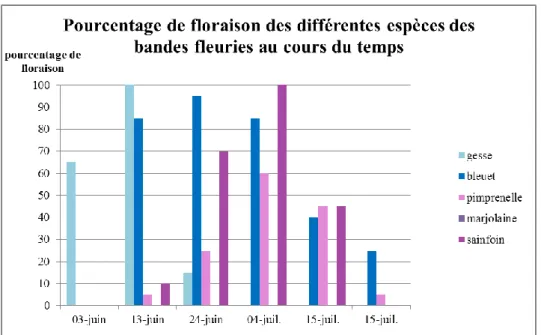
Figure 13 : suivi de la floraison des différentes espèces composant les bandes fleuries pendant la durée de la saison culturale
23
La modalité bande fleurie (BF) est composée d’un mélange de cinq espèces (figure 11):
le bleuet (Centaurea cyanus, famille des Astéracées),
la gesse (Lathyrus sativus, famille des Fabacées),
la marjolaine (Origanum majorana, famille des Lamiacées),
la pimprenelle (Sanguisorba minor, famille des Rosacées),
le sainfoin (Onobrychis viciifolia, famille des Fabacées).
Les modalités BE et BF sont semées le 21 mars (annexe II : itinéraire technique de l’essai Parcel-R 2013).
Le développement des bandes enherbées et fleuries est contrôlé à plusieurs reprises afin de déterminer la date optimale de la plantation des melons. Un quadrat de 1m² est positionné sur les bandes et permet d’évaluer le pourcentage de couverture du sol par la végétation. Une couverture proche de 100% est nécessaire (figure 12). Les melons ont été plantés le 24 mai (annexe II).
Pendant le développement de la culture, le stade phénologique des différentes espèces de la bande fleurie est évalué (échantillons de 10 individus par espèce, pris au hasard). La présence de fleurs, pour subvenir aux besoins des auxiliaires de culture, est nécessaire pendant toute la saison culturale (figure 13).
Les floraisons des différentes espèces végétales se sont échelonnées au cours de la culture du melon. La gesse était en fleurs dès la plantation. A partir de mi-juin, deux à quatre espèces végétales étaient fleuries en même temps. Contrairement aux années précédentes, la floraison de la marjolaine n’a été détectée au cours du suivi.
1.4 Données météorologiques
Elles proviennent de la station agro-climatique implantée sur le domaine expérimental St Paul de Montfavet. La base de données agro-climatique de l’Inra est gérée par l’Unité Agroclim basée à Montfavet et l’extraction de données se fait via l’interface web Inra Climatik. Les données relatives à la pluviométrie, aux températures et au vent ont été utilisées.