www.elsevier.com / locate / cardiores
S
tatins (HMG-CoA reductase inhibitors) reduce CD40 expression in
human vascular cells
a c a ,
*
aFlore Mulhaupt , Christian M. Matter , Brenda R. Kwak
, Graziano Pelli ,
a a b c a
¨
Niels R. Veillard , Fabienne Burger , Pierre Graber , Thomas F. Luscher , Franc¸ois Mach
a
Cardiology Division, Department of Medicine, University Hospital, Geneva Medical School, Foundation for Medical Research, 64 Avenue Roseraie, 1211 Geneva 4, Switzerland
b
Serono Pharmaceutical Research Institute, Geneva, Switzerland c
Division of Cardiology, Cardiovascular Research, Institute of Physiology and Division of Cardiology, Cardiovascular Center, University Hospital, Zurich, Switzerland
Received 2 December 2002; accepted 30 May 2003
Abstract
Objective: HMG-CoA reductase inhibitors (statins) possess anti-inflammatory and immunomodulatory properties that are independent
of their lipid-lowering action. As the CD40–CD40L signaling pathway is implicated in the modulation of inflammatory responses between vascular cells, involving adhesion molecules, pro-inflammatory cytokines, chemokines, we sought to investigate the potential role of statins in regulating the expression of CD40. Methods and Results: Using Western blot, flow cytometry and immunohistochemistry analyses, we observed that four different statins reduced IFN-g-induced CD40 expression in human vascular cells (endothelial cells, smooth muscle cells, macrophages and fibroblasts). This effect was dose-dependent (from 5 mM to 80 nM) and reversed by addition of
L-mevalonate. Activation of vascular cells by human recombinant CD40L, as measured by ELISA for IL-6, IL-8 and MCP-1, was strongly
reduced when cells were treated with statins. Immunostaining of human carotid atherosclerotic lesions of patients subjected to statin treatment revealed less CD40 expression on a ‘per vascular cell’ basis compared to control patients. Although many pleiotropic effects of statins are mediated by nitric oxide synthase (NOS)- or peroxisome proliferator-activated receptor (PPAR)-dependent signaling pathways, we observed similar statin-induced reduction of CD40 expression using NOS inhibitors or different PPAR ligands. Conclusion: Statins decrease CD40 expression and CD40-related activation of vascular cells. These effects are partially reversed by the HMG-CoA reductase product L-mevalonate and are mediated by NOS- or PPAR-dependent pathways. Altogether, these findings provide mechanistic insight
into the beneficial effects of statins on atherogenesis. They also provide a scientific rationale for the use of statins as immunomodulators after organ transplantation.
2003 European Society of Cardiology. Published by Elsevier B.V. All rights reserved.
Keywords: Atherosclerosis; Cytokines; Immunology; Lipid metabolism; Receptors
1 . Introduction that these drugs greatly reduce cardiovascular-related
morbidity and mortality in patients with and without Statins are effective lipid-lowering agents, extensively coronary disease [2,3]. Recently, in vitro and in vivo used in medical practice [1]. They are competitive in- findings have indicated that statins, beside their lipid-hibitors of 3-hydroxy-3-methylglutaryl coenzyme A lowering effects, possess anti-inflammatory properties[4]. (HMG-CoA) reductase, an enzyme crucial for cholesterol In addition, we recently demonstrated that these drugs synthesis. Several large clinical trials have demonstrated might also be recognized as a new type of
immuno-suppressor[5].
Atherosclerosis is considered as an immuno-inflamma-*Corresponding author. Tel.: 7233; fax:
141-22-382-tory disease [6,7] and increasing evidence suggests a
7245.
E-mail address: [email protected] (B.R.
Kwak). Time for primary review 32 days.
0008-6363 / 03 / $ – see front matter 2003 European Society of Cardiology. Published by Elsevier B.V. All rights reserved. doi:10.1016 / S0008-6363(03)00515-7
756 F. Mulhaupt et al. / Cardiovascular Research 59 (2003) 755–766
central role for the CD40–CD40L signaling pathway in the bodies for IL-6, IL-8 and MCP-1 were obtained from pathogenesis of this disease [8,9]. Recently, two reports R&D.L-Mevalonate was purchased from Sigma (St. Louis,
demonstrated that hypercholesterolemia, one of the major MO, USA). Human recombinant CD40 ligand (rCD40L) cardiovascular risk factors, increases CD40 and CD40L was generated as described previously[17].L-NAME was
expression on platelets and monocytes [10] as well as purchased from Sigma. 15d-PGJ2 was obtained from plasma levels of soluble CD40L [11]. In addition, it has Calbiochem (La Jolla, CA, USA), ETYA from Sigma, been shown that blocking CD40–CD40L interactions WY14643 from Biomol (Plymouth Meeting, PA, USA), significantly prevents the development of atherosclerotic and Troglitazone a gift from Park Davis Pharmaceuticals plaques and reduces already pre-established lesions (Morris Plains, NJ, USA).
[12,13].Moreover, CD40 signaling has been implicated in
other chronic disorders such as rheumatoid arthritis, multi- 2 .2. Cells ple sclerosis and allograft rejection following organ
trans-plantation[14]. The investigation conforms with the principles outlined
CD40L (recently renamed CD154) and CD40 are mem- in the Declaration of Helsinki (Cardiovasc Res 1997;35:2-bers of the tumor necrosis factor (TNF) and TNF-receptor 3) as well as the Guide for the Care and Use of family, respectively. The original function of CD40L in T Laboratory Animals published by the US National Institute cell-dependent humoral immunity involves the activation of Health (NIH publication No. 85-23, revised 1996). and differentiation of B-lymphocytes, the switching of Human vascular ECs and SMCs were isolated from immunoglobulin classes as well as the formation of saphenous veins and cultured as described previously germinal center and memory cells. More recently, activa- [5,18].Primary human fibroblasts were isolated from nasal tion of vascular cells such as macrophages (MF), endo- polyps and cultured as previously described [19]. Cells thelial cells (ECs) and smooth muscle cells (SMCs) via were used at passages two to four for all experiments. CD40 signaling has been shown to induce inflammatory Human monocytes were obtained from peripheral blood of responses with expression of adhesion molecules and healthy donors as described previously [20]. They were secretion of pro-inflammatory cytokines, chemokines, ma- grown in RPMI 1640 medium containing 10% fetal calf trix metalloproteinases and tissue factor [15]. All these serum (FCS) for 10 days. The human Raji cell line was molecules are known to be crucial in the pathogenesis of obtained from ATCC (Rockville, MD, USA).
atherosclerosis as well as during graft-versus-host disease.
In view of the large clinical beneficial effects of statins 2 .3. Western blot analysis on cardiovascular morbidity and mortality, and the
emer-gence of numerous non-lipid lowering anti-inflammatory Cells were harvested in ice-cold radio-immuno-precipi-effects of these drugs, we sought to investigate whether tation assay (RIPA) solubilization buffer, and total amounts statins regulate the expression of CD40 in vascular cells. of protein determined using a bicinchoninic acid quantifi-cation assay (Pierce, Rockford, IL, USA). Western blotting was performed as described before[5].Blots were blocked
2 . Methods in 5% defatted dry milk / phosphate-buffered saline (PBS) /
0.1% Tween, incubated for 1 h at room temperature with
2 .1. Reagents primary antibody (1:40; rabbit polyclonal anti-CD40), or
mouse monoclonal anti-human b-actin (1:5000; Pharming-Human and mouse recombinant IFN-g were obtained en) as control of loading, followed by 1 h incubation with from Endogen (Cambridge, MA, USA) and R&D Systems secondary antibody (1:10,000; Jackson ImmunoResearch). (Abingdon, UK), respectively. The statins used in these Quantification was performed using AIDA software studies, i.e. atorvastatin, simvastatin, lovastatin and pravas- (Raytest, Urdorf, Switzerland).
tatin were obtained from commercial sources. Because
ECs lack lactonases, to process simvastatin and lovastatin 2 .4. Flow cytometry to their active forms, these agents were chemically
acti-vated before their use as previously described[16].Mouse Cells were incubated with FITC-conjugated specific anti-human CD40 monoclonal antibody fluorescein iso- antibody (60 min, 4 8C) and analyzed in a Becton Dickin-thiocyanate-conjugated (FITC) and mouse anti-human son FACScan flow cytometer as described [5]. At least HLA-DR monoclonal antibody (FITC) were obtained from 100,000 viable cells were analyzed per condition. Data Pharmingen (San Diego, CA, USA). Rabbit anti-human were analyzed using CELLQUEST software (Becton Dic-CD40 polyclonal antibody, anti-rabbit IgG FITC, and HRP kinson).
(horseradish peroxidase) goat anti-rabbit IgG were
pur-chased from Santa Cruz (Santa Cruz, CA, USA), Jackson 2 .5. Cytokine assays ImmunoResearch (West Grove, PA, USA) and Vector
sandwich-type enzyme-linked immunosorbent assay snap-frozen in liquid nitrogen. There was no significant (ELISA) according to the manufacturer’s instructions difference for age, gender, cardiovascular risk factors and (R&D). Experiments were performed in the presence of other medications between patients treated or not with polymyxin B (1 mg / ml). Optical density was measured at simvastatin (Table 1). Serial cryostat sections (5 mm) were 450 nm using a Dynatech plate reader (Chantilly, VA, cut, air dried onto microscope slides (Fisher Scientific, USA). The amount of IL-6, IL-8 and MCP-1 detected was Wohlen, Switzerland), and fixed in acetone at 220 8C for 5 calculated from a standard curve prepared with the recom- min. Sections were pre-incubated with blocking buffer binant protein. Samples were assayed in duplicate. (PBS / Tween with 8% normal horse serum) and then incubated successively with an anti-human CD40 antibody
2 .6. Fluorescence immunolabeling for 1 h. Thereafter, sections were incubated with
biotinylated secondary antibody (45 min; Vector Laborator-Monocytes / macrophages grown on coverslips were ies) followed by avidin–biotin–alkaline phosphatase com-rinsed and fixed for 15 min with paraformaldehyde (4%). plex (Vectastain ABC kit). Antibody binding was visual-Coverslips were rinsed and cells incubated successively in ized by alkaline phosphatase substrate (Vector Laborator-0.5 M NH Cl / PBS for 15 min and PBS supplemented with4 ies). Quantitative analyses for CD40 and CD68 expression 2% bovine serum albumin (Sigma) for a further 20 min. were performed with a computer-based color image analy-Human macrophages were incubated overnight with pri- sis system [21].
mary rabbit polyclonal anti-CD40 (1:50) in 10% normal
goat serum (Sigma) / PBS. After rinsing, human macro- 2 .8. Statistical analysis phages were incubated with secondary antibodies
FITC-conjugated (1:800) for 3 h. All steps were performed at Data are presented as mean6S.E.M. Mean values room temperature and between incubation steps cells were between two groups were compared using a two-tailed rinsed with PBS. Finally, coverslips were mounted on Student’s t-test, after having performed an F-test for slides in Vectashield (Vector Laboratories). Cells were homogeneity of variances. If data failed to meet require-examined using a Zeiss Axiophot microscope. Replace- ments for use of the parametric t-test, a Mann–Whitney ment of the primary antibody with PBS / 10% normal goat U-test was used. Differences were considered statistically serum was used to control the specificity of the immuno- significant at P,0.05.
labeling of human macrophages.
2 .7. Immunohistochemistry 3 . Results
Surgical specimens of human carotid atheroma were 3 .1. Statins reduce CD40 expression on vascular cells obtained by protocols approved by the Review Committee
at the University Hospital of Geneva. All atherosclerotic To determine whether statins influence CD40 expres-carotid specimen analyzed were obtained from patients sion, human vascular endothelial cells (ECs) were acti-treated (for at least 3 months) or not with the simvastatin. vated with human recombinant IFN-g (1000 U / ml / 24 h) Materials were directly embedded in OCT compound or in the presence or absence of atorvastatin, lovastatin, pravastatin or simvastatin. As previously described, un-stimulated vascular cells showed little expression of CD40,
T able 1 which was greatly induced by IFN-g [18]. Western blot
Baseline characteristics of patients in the simvastatin and control groups
analysis (Fig. 1A) revealed that statin treatment reduced
Simvastatin (n518) Control (n514) both, basal CD40 expression as well as IFN-g-induced
CD40. Simvastatin appeared to be the most potent
inhib-Age (years) 68 65
Sex (F / M) 8 / 10 8 / 6 itor, followed by lovastatin, pravastatin and atorvastatin
Diabetes (%) 32 38 (Fig. 1A). Raji cells, known to express a large amount of
Smokers (%) 34 30
CD40, were used as positive control (Fig. 1A,lane 8). The
Hypertension (%) 73 68
reduction of CD40 expression by statins was
dose-depen-Medication (%)
dent (Fig. 1B). The concentration range (0.08 to 10 mM)
Aspirin 33 28
ACEIs 45 48 of statins used in these experiments was similar to earlier
b-Blockers 24 22 reports [5,22], and neither cell survival nor protein
syn-Anti-diabetic agents 33 38
thesis was affected at these concentrations (data not
Diuretics 32 21
shown). Interestingly, simvastatin treatment significantly reduced CD40 expression (Fig. 1B) at a concentration of
Total cholesterol (mmol / l) 5.3 (61.3) 6.4 (61.4)
LDL-c, 3.3 (61.1) 4.1 (61.2) only 80 nM, which corresponds to the effective serum
HDL-c 1.3 (60.3) 1.2 (60.3) levels seen with statin treatment in clinical practice [23]. Triglycerides 1.9 (61.4) 2.3 (61.4)
for the inhibition of the HMG-CoA reductase enzyme, under statin therapy expressed much less CD40 compared since addition ofL-mevalonate largely reversed the statin- to control patients (Fig. 4A,B). In addition, we observed
induced inhibition of CD40 expression (Fig. 1A). Similar that individual macrophages within lesions from statin-results were obtained for human vascular SMCs (Fig. 1C). treated patients showed markedly less CD40 expression These results were confirmed using flow cytometry analy- compared to individual macrophages in lesions of control sis (Fig. 1D). Western blot results were supported by patients (Fig. 4C). This reduction was mainly detected immunohistochemical analysis on human monocyte– within the shoulder region of the atherosclerotic plaques. macrophages (Fig. 2A–F) and on human fibroblasts (Fig.
2G–I), two other cell types present in atherosclerotic 3 .4. Statin reduction of CD40 expression is neither lesions. Taken together, these experiments demonstrate mediated via nitric oxide nor via PPARs
that statin treatment decreased CD40 expression on all
human vascular cells tested. Due to numerous recent reports demonstrating that
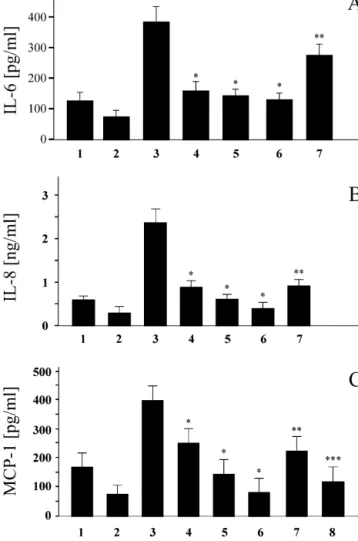
several anti-inflammatory effects of statins could be largely 3 .2. Statins inhibit functional CD40 signaling related to their ability to stabilize and increase nitric oxide synthase (NOS) [25], we investigated whether statins To examine functional consequences of statin treatment reduce CD40 expression via NOS pathways. To address on CD40 expression, we stimulated vascular cells with this question, we stimulated human ECs with recombinant human recombinant CD40 ligand (rCD40L) in the presence IFN-g and simvastatin in the presence or absence of the or absence of the statins. As presented inFig. 3for ELISA NOS inhibitor L-NAME. As shown in Fig. 5 by flow
experiments on ECs, rCD40L induced the secretion of the cytometry analysis, the level of CD40 expression after pro-inflammatory molecules IL-6, IL-8 and MCP-1, as statin treatment was similarly reduced with or without previously described [17,24]. When stimulated in the L-NAME treatment.
presence of the statins, the synthesis and secretion of all Furthermore, as statins have been recently reported to these pro-inflammatory proteins were largely reduced. As activate PPARs [26], we investigated whether PPAR previously described, statins also reduced constitutive ligands could regulate CD40 expression on human vascular cytokine secretion, these effects were however not statisti- cells. We did not detect any effect of either PPAR-a or cally significant. The statin-induced reduction in cytokine / PPAR-g ligands on CD40 cell surface expression following chemokine secretion was partially reversed by the addition IFN-g activation (Fig. 6). These findings indicate that of L-mevalonate (Fig. 3A–C). Specificity of these experi- neither NOS nor PPARs are involved in the reduction of
ments was confirmed by blocking CD40–CD40L signaling CD40 expression by statins on vascular cells. with the addition of an anti-CD40L antibody (Fig. 3C).
3 .3. Statin treatment decreases CD40 expression within 4 . Discussion human atherosclerotic tissue
Increasing evidence supports the central role of the In view of our findings that statins reduce CD40 CD40–CD40L signaling pathway in atherosclerosis. Acti-expression and CD40 signaling in vitro, we decided to vation of vascular cells via CD40–CD40L interactions has explore the expression of CD40 on vascular cells within been shown to induce inflammatory responses with expres-human atherosclerotic lesions obtained from patients sion of adhesion molecules, secretion of pro-inflammatory treated or not with simvastatin (20 mg / day for at least 3 cytokines, matrix metalloproteinases, tissue factor and months). Baseline patients characteristics were not statisti- chemokines [8,9,18], molecules considered as crucial cally different and are shown in Table 1.Immunohistoch- players in atherogenesis. Recent in vitro and in vivo emical in situ and Western blot analyses of human carotid findings suggest that statins also have anti-inflammatory arteries revealed that atherosclerotic lesions from patients properties[24,27].Statins inhibit HMG-CoA reductase that
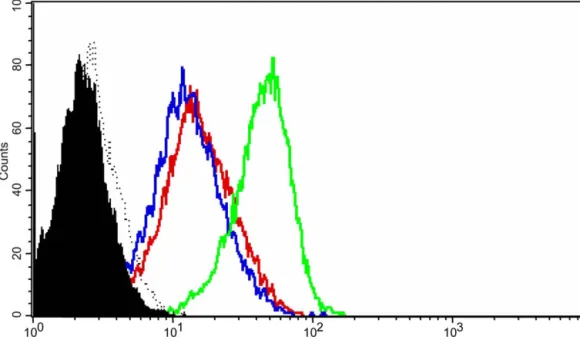
Fig. 1. Statins decrease IFN-g-induced CD40 expression on human endothelial cells. (A–C) Western blot analysis for CD40. (A) Human vascular ECs under unstimulated conditions (1); treated with IFNg (1000 U / ml, 24 h) alone (2); or in the presence of atorvastatin (10 mM) (3); lovastatin (10 mM) (4); pravastatin (20 mM) (5); simvastatin (10 mM) (6); or simvastatin (10 mM) andL-mevalonate (400 mM) (7); Raji cells under unstimulated condition as
positive control (8). *P,0.05 (compared to 2); **P,0.05 (compared to 6). Quantification of Western blots is expressed as ratio of CD40 / b-actin signal for each samples. (B) Human vascular ECs under unstimulated conditions (Ctrl), or treated with IFNg (1000 U / ml, 24 h) alone; or in the presence of simvastatin (5 to 0.08 mM). b-actin is shown as loading control. (C) Human vascular SMCs under unstimulated conditions (1), treated with IFNg (500 U / ml, 24 h) alone (2); or in the presence of simvastatin (10 mM) (3). b-actin is shown as loading control. (D) Flow cytometry analysis for CD40. Human vascular ECs treated with IFNg (1000 U / ml, 24 h) alone (green line); or in the presence of simvastatin (5 mM) (blue line). Expression of MHC class II for ECs stimulated with IFNg (1000 U / ml, 24 h) is shown as positive control (red line). Shown is a histogram representing cell numbers ( y-axis) vs. CD40 log fluorescence intensity (x-axis) for 20,000 viable cells, and ECs under unstimulated conditions are shown (solid histograms) as well as isotype controls (dotted lines). For all analysis, similar results were obtained in separate experiments using cells from four different donors.
760 F. Mulhaupt et al. / Cardiovascular Research 59 (2003) 755–766
Fig. 2. Statins reduce CD40 expression on human macrophages and fibroblasts. Immunohistochemical analysis of CD40 expression (green fluorescence). Macrophages (A–F) and fibroblasts (G–I). Cells were counterstained with Evans Blue (red color). (A) Cells under unstimulated conditions. (B) Cells treated with IFN-g alone (1000 U / ml, 24 h); or in the presence of simvastatin (5 mM) (C); simvastatin (1 mM) (D); or pravastatin (20 mM) (E). (F) Cells treated with IFN-g (1000 U / ml, 24 h) and stained with secondary antibody only (negative control). (G) Cells under control conditions; treated with IFN-g alone (500 U / ml, 24 h) (H); or in the presence of simvastatin (10 mM) (I). Scale bar represents 50 mm. Similar results were obtained in separate experiments using cells from three different donors.
transforms HMG-CoA into mevalonate. Indeed, as mevalo- CD40–CD40L in atherogenesis as well as the pleiotropic nate is not only the precursor of cholesterol but also of effects of statins in this disease, we sought to investigate in many non-steroidal isoprenoids, statins may regulate the vitro and in vivo the effect of statins on CD40 expression expression and activities of important molecules in the as well as on related CD40 activation in human vascular pathogenesis of atherosclerosis. Given the importance of cells.
showed that the reduction of CD40 expression by statins is not mediated via mevalonate synthesis, but are in agree-ment with three other recent reports[10,29,30].
All four statins used in this study reduce IFN-g-induced CD40 expression: simvastatin being the strongest, fol-lowed by lovastatin, pravastatin and atorvastatin. Although other in vitro studies have also revealed differences in potency of individual statins, these results should be interpreted with great care. One possible explanation for the strong effect of simvastatin on IFN-g-induced CD40 expression might be that this specific statin possesses additional anti-inflammatory effects through inhibitory actions in other signaling pathways. Alternatively, simply due to different pharmacokinetic properties simvastatin may be more effectively processed by cells in culture. It is of importance to note in this respect that in vivo in mouse models both atorvastatin and simvastatin are effective treatments against multiple sclerosis and rheumatoid arth-ritis [30,31].
CD40 signaling plays a critical role in the initiation of the immune response by inducing inflammatory responses with secretion of pro-inflammatory cytokines and chemokines. These chemokines probably attract and direct T lymphocytes and macrophages to the atheroma, thus sustaining chronic inflammation [32,33]. Indeed, lack of chemoattractants or their receptors is known to signifi-cantly diminish atherosclerotic lesion progression in mice [34,35]. We found that statin treatment influences func-tional cellular CD40 signaling by reducing the release of cytokines and chemokines. By secreting these pro-inflam-matory mediators and growth factors, vascular cells can
Fig. 3. Statins reduce CD40 function on CD40L-activated human
vascu-lar endothelial cells. IL-6 (A), IL-8 (B) and MCP-1 (C) releases measured promote the migration and proliferation of SMCs. In
by ELISA in supernatants of ECs exposed (24 h) to normal media (1); response to inflammatory stimulation, SMCs and MF simvastatin (10 mM) alone (2); activated by rCD40L (5 mg / ml) alone
secrete MMPs that will subsequently degrade matrix
(3); or in the presence of atorvastatin (10 mM) (4); lovastatin (10 mM)
compounds of the plaque as well as other inflammatory
(5); simvastatin (10 mM) (6), or simvastatin (10 mM) andL-mevalonate
mediators that inhibit collagen synthesis. As extracellular
(400 mM) (7); rCD40L (5 mg / ml) preincubated with polyclonal CD40L
antibody (8). Similar results were obtained in independent experiments matrix components are considered crucial determinants of
with ECs from four different donors. *P,0.05 (compared to 3); **P, fibrous cap integrity and stability, statin-induced reduction 0.05 (compared to 6); ***P,0.05 (compared to 3).
of CD40 signaling with consequent reduction in secretion of pro-inflammatory cytokines and chemokines may not Pro-inflammatory cytokines expressed within atheroma only exert beneficial actions on progression of atheros-provide a chemotactic stimulus to adherent leukocytes, clerotic plaque development but on the composition of the directing their migration into the intima. Our data show by plaque as well. Indeed, one of the major complications in different techniques that statin treatment largely reduces clinical cardiology is plaque rupture with local thrombus CD40 expression both on human ECs and on monocyte– formation, potentially leading to acute coronary macrophages, thereby potentially reducing EC–monocyte syndromes.
interactions. Similar results were obtained on murine Interruption of CD40–CD40L interactions by adminis-macrophages (data not shown). Importantly, we observed a tration of anti-CD40L blocking antibody or gene targeting reduction of CD40 expression at a concentration of 80 nM, has been shown to prevent atherogenesis and limit the which is within the range of effective serum concentrations progression of established atherosclerotic lesions in mice seen in clinical practice [23]. In addition, L-mevalonate [12,13]. Both strategies have resulted in a decrease in
largely reversed the statin-induced decreased in CD40 pro-inflammatory factors and an increase in collagen expression, indicating that indeed the inhibition of HMG- content of atherosclerotic lesions. Recently, it has been CoA reductase mediates the statin-induced repression of shown that in a short prospective treatment period statins CD40 on vascular cells. These results are partly in reduced the pro-inflammatory state of human atheromatous contradiction with the data from Wagner et al.[28],which lesions[36].Our results showing that patients under statin
therapy express less CD40-positive cells within atheros- deed, clinical studies have suggested a better outcome after clerotic plaques compared to control patients without statin cardiac transplantation in patients subjected to statin therapy. In addition, they show that individual macro- therapy[39,40].We recently demonstrated that statins are phages express less CD40, suggesting direct effects of involved in the immune response by inhibiting IFN-g
statins on macrophages in vivo. induced MHC-II [5],and thus might be used as
immuno-Most of the anti-inflammatory effects of statins have modulators. In addition, CD40 has been shown to play a been related to their ability to stabilize and increase NO crucial role in graft-versus-host disease [41,42]. Our synthase[4,22].This endogenous vasodilator molecule has current findings indicating that statins decrease CD40 by itself anti-inflammatory properties [37]. However, our expression on antigen presenting cells may therefore experiments illustrate that the reduction of CD40 expres- provide a new scientific rationale for their use as immuno-sion after statin treatment does not involve the NOS modulators following organ transplantation.
signaling pathway. Recently, statins have been reported to In summary, we show that statins decrease CD40 activate PPARs [26].In addition, we recently reported that expression in vascular cells in vitro as well as in vivo. By PPAR-g-induced repression of major histocompatibility reducing CD40 expression and signaling, statins may complex class II (MHC-II), another molecule involved in reduce inflammation and thus stabilize atherosclerotic the immune response [38].We did not detect an effect of plaques, features believed to account for the beneficial either PPAR-a or PPAR-g ligands on CD40 cell surface effects of statins on cardiovascular morbidity and mortali-expression following IFN-g activation, thus excluding also ty. Our findings not only provide an additional explanation these signaling pathways from mediating statin-induced for previously unknown non-lipid effects of statins in reduction in CD40 expression. atherogenesis, but also suggest their application in further One of the major problems following organ transplanta- immuno-inflammatory diseases in which CD40 is in-tion is the development of graft-versus-host disease. In- volved, such as multiple sclerosis, systemic lupus
erythe-Fig. 5. Statins effect on CD40 expression is not mediated through NOS. Flow cytometry analysis for CD40 expression. Human vascular ECs treated with IFNg (1000 U / ml, 24 h) alone (green line); or with simvastatin (5 mM) (blue line); or in the presence of simvastatin (5 mM) andL-NAME (5 mM) (red
line). Shown is a histogram representing cell numbers ( y-axis) vs. CD40 log fluorescence intensity (x-axis) for 20,000 viable cells, and ECs under unstimulated conditions are shown (solid histograms) as well as isotype controls (dotted lines). Similar results were obtained in independent experiments with ECs from three different donors.
Fig. 4. Statin therapy decreases CD40 expression on human atherosclerotic lesions. (A) Representative photomicrographs (3100) of CD40 expression (red color) within human carotid atherosclerotic lesions in situ. Atherosclerotic lesion in patient treated (1) or not (2) by statin therapy. L, lumen of the vessel. Quantitative analysis of CD40 expression on atherosclerotic lesions obtained from patients treated with statin (1) (n518), or without statin therapy (2) (n514) is shown on the right side of this panel. *P,0.05. (B) Western blot analysis of human carotid atherosclerotic tissues obtained from patients treated (1) or not (2) with statin therapy. Quantitative analysis of CD40 expression in patients treated with statin (1) (n512), or without statin therapy (2) (n510) is shown on the right side of this panel. *P,0.05. (C) Double immunofluorescence stain for macrophages (CD68) in green and CD40 in red in patient treated (1) or not (2) by statin therapy. *P,0.05. Similar results were obtained in separate experiments using tissues from five different donors.
764 F . Mulhaupt et al . / Cardiovascular Research 59 (2003) 755 – 766
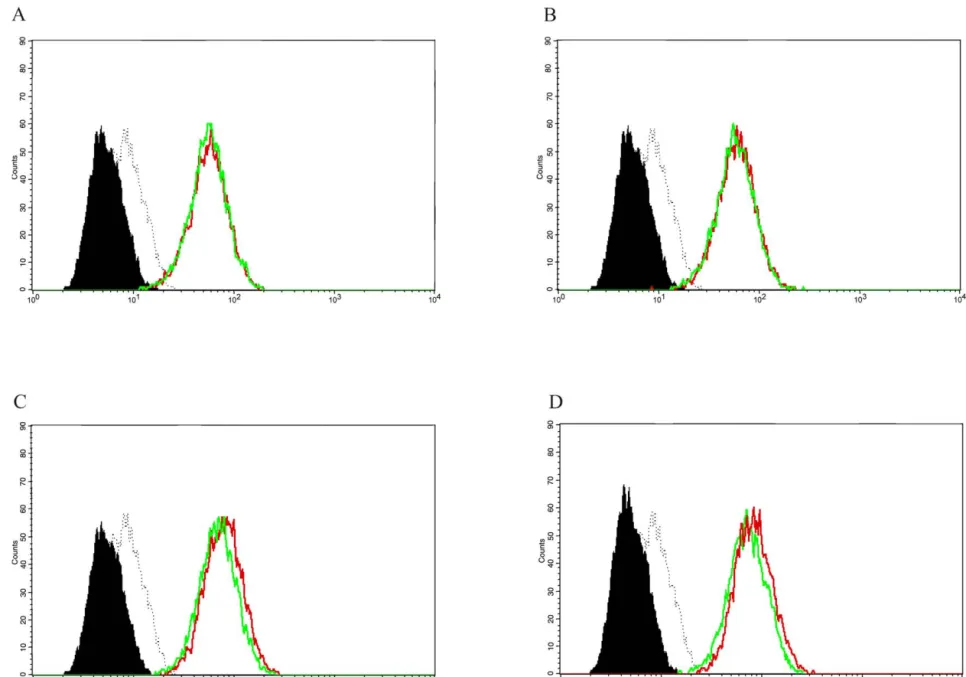
Fig. 6. Effects of PPAR ligands on IFN-g-induced CD40 expression on human endothelial cells. Flow cytometry analysis for CD40 expression. Human vascular endothelial cells (ECs) treated with IFN-g (500 U / ml, 24 h) alone (green line), or with the PPAR-g ligands (red lines) 15d-PGJ (10 mmol / l) (A), Troglitazone (50 mmol / l) (B); or with the PPAR-a ligands (red lines) ETYA (50 mmol / l)2 (C), and WY14643 (50 mmol / l) (D). Each panel is a histogram representing cell numbers ( y-axis) vs. log fluorescence intensity (x-axis) for 15,000 viable cells, and ECs under unstimulated conditions are shown (solid histograms) as well as isotype controls (dotted lines). Similar results were obtained in independent experiments with ECs from five different donors.
is expressed on human vascular endothelial cells, smooth muscle
matosus, autoimmune diabetes mellitus, chronic arthritis
cells, and macrophages: implications for CD40–CD40 ligand
sig-and graft-versus-host disease.
naling in atherosclerosis. Proc Natl Acad Sci USA 1997;94:1931– 1936.
[19] W iszniewski L, Limat A, Saurat JH, Meda P, Salomon D.
Differen-A cknowledgements tial expression of connexins during stratification of human keratino-cytes. J Invest Dermatol 2000;115:278–285.
[20] K ol A, Sukhova GK, Lichtman AH, Libby P. Chlamydial heat shock
The authors thank Dr Ludovic Wiszniewski (Department
protein 60 localizes in human atheroma and regulates macrophage
of Pediatrics, Geneva University Hospital) for providing us tumor necrosis factor-alpha and matrix metalloproteinase expres-with primary human fibroblasts. This work was supported sion. Circulation 1998;98:300–307.
by grants from the Swiss National Research Foundation [21] A ikawa M, Voglic SJ, Sugiyama S et al. Dietary lipid lowering reduces tissue factor expression in rabbit atheroma. Circulation
(grants 3200-054965.98 / 1 and 3200-065121.01 / 1) to FM;
1999;100:1215–1222.
(3234-066311.01 / 1 and 3100-067777.02) to BRK; and
[22] L aufs U, La Fata V, Plutzky J, Liao JK. Upregulation of endothelial
(32-510069.97) to TFL. Additional support for this work nitric oxide synthase by HMG CoA reductase inhibitors. Circulation was obtained from the Leenaards Foundation (to BRK and 1998;97:1129–1135.
FM), the Swiss Heart Foundation of Cardiology and the [23] I llingworth DR, Erkelens DW, Keller U, Thompson GR, Tikkanen MJ. Defined daily doses in relation to hypolipidaemic efficacy of
Adumed Foundation (to CMM).
lovastatin, pravastatin, and simvastatin. Lancet 1994;343:1554– 1555.
[24] D iomede L, Albani D, Sottocorno M et al. In vivo anti-inflammatory
R eferences effect of statins is mediated by nonsterol mevalonate products.
Arterioscler Thromb Vasc Biol 2001;21:1327–1332.
[25] L aufs U, Liao JK. Direct vascular effects of HMG-CoA reductase [1] M aron DJ, Fazio S, Linton MF. Current perspectives on statins.
inhibitors. Trends Cardiovasc Med 2000;10:143–148. Circulation 2000;101:207–213.
[26] M artin G, Duez H, Blanquart C, Berezowski V, Poulain P, Fruchart [2] L aRosa JC. Statins and risk of coronary heart disease. J Am Med
JC, Najib-Fruchart J, Glineur C, Staels B. Statin-induced inhibition Assoc 2000;283:2935–2936.
of the Rho-signaling pathway activates PPARalpha and induces [3] V aughan CJ, Gotto Jr. AM, Basson CT. The evolving role of statins
HDL apoA-I. J Clin Invest 2001;107:1423–1432. in the management of atherosclerosis. J Am Coll Cardiol
[27] S parrow CP, Burton CA, Hernandez M et al. Simvastatin has 2000;35:1–10.
anti-inflammatory and antiatherosclerotic activities independent of [4] T akemoto M, Liao JK. Pleiotropic effects of
3-hydroxy-3-plasma cholesterol lowering. Arterioscler Thromb Vasc Biol methylglutaryl coenzyme A reductase inhibitors. Arterioscler
2001;21:115–121. Thromb Vasc Biol 2001;21:1712–1719.
¨
[28] W agner AH, Gebauer M, Gudenzorph B, Hecker M. 3-Hydroxy-3-[5] K wak B, Mulhaupt F, Myit S, Mach F. Statins as a newly
methylglutaryl coenzyme A reductase-independent inhibition of recognized type of immunomodulator. Nat Med 2000;6:1399–1402.
CD40 expression by atorvastatin in human endothelial cells. Ar-[6] L ibby P, Ridker PM, Maseri A. Inflammation and atherosclerosis.
terioscler Thromb Vasc Biol 2002;22:1784–1789. Circulation 2002;105:1135–1143.
¨
[29] S chonbeck U, Gerdes N, Varo N et al. Oxidized low-density [7] G lass CK, Witztum JL. Atherosclerosis. The road ahead. Cell
lipoprotein augments and 3-hydroxy-3-methylglutaryl coenzyme A 2001;104:503–516.
reductase inhibitors limit CD40 and CD40L expression in human [8] S chonbeck U, Libby P. The CD40 / CD154 receptor / ligand DYAD.
vascular cells. Circulation 2002;106:2888–2893. Cell Mol Life Sci 2001;58:4–43.
¨
[30] Y oussef S, Stuve O, Patarroyo JC et al. The HMG-CoA inhibitor, [9] L utgens E, Daemen MJ. CD40–CD40L interactions in
atherosclero-atorvastatin, promotes a Th2 bias and reverses paralysis in central sis. Trends Cardiovasc Med 2002;12:27–32.
nervous system autoimmune disease. Nature 2002;420:78–84. [10] G arlichs CD, John S, Schmeisser A et al. Upregulation of CD40 and
[31] L eung BP, Satta N, Crilly A et al. A novel anti-inflammatory role for CD40 ligand (CD154) in patients with moderate
hypercholes-simvastatin in inflammatory arthritis. J Immunol 2003;170:1524– terolemia. Circulation 2001;104:2395–2400.
1530. [11] C ipollone F, Mezzetti A, Porreca E et al. Association between
[32] L uster AD. Chemokines—chemotactic cytokines that mediate in-enhanced soluble CD40L and prothrombotic state in
hypercholes-flammation. N Engl J Med 1998;338:436–445. terolemia. Effects of statin therapy. Circulation 2002;106:399–402.
[33] M ach F. The role of chemokines in atherosclerosis. Curr Atheroscler [12] M ach F, Schonbeck U, Sukhova GK, Atkinson E, Libby P.
Rep 2001;3:243–251. Reduction of atherosclerosis in mice by inhibition of CD40
sig-[34] L u B, Rutledge BJ, Gu L et al. Abnormalities in monocyte nalling. Nature 1998;394:200–203.
recruitment and cytokine expression in monocyte chemoattractant [13] L utgens E, Gorelik L, Daemen MJ et al. Requirement for CD154 in
protein 1-deficient mice. J Exp Med 1998;187:601–608. the progression of atherosclerosis. Nat Med 1999;5:1313–1316.
[35] G u L, Okada Y, Clinton SK et al. Absence of monocyte chemoat-[14] F oy TM, Aruffo A, Bajorath J, Buhlmann JE, Noelle RJ. Immune
tractant protein-1 reduces atherosclerosis in low density lipoprotein regulation by CD40 and its ligand GP39. Annu Rev Immunol
receptor-deficient mice. Mol Cell 1998;2:275–281. 1996;14:591–617.
[15] S chonbeck U, Mach F, Libby P. CD154 (CD40 ligand). Int J [36] C risby M, Nordin-Fredriksson G, Shah PK et al. Pravastatin Biochem Cell Biol 2000;32:687–693. treatment increases collagen content and decreases lipid content, [16] B lum CB. Comparison of properties of four inhibitors of 3-hydroxy- inflammation, metalloproteinases, and cell death in human carotid 3-methylglutaryl-coenzyme A reductase. Am J Cardiol 1994;73:3D– plaques: implications for plaque stabilization. Circulation
11D. 2001;103:926–933.
[17] M azzei GJ, Edgerton MD, Losberger C et al. Recombinant soluble [37] D e Caterina R, Libby P, Peng HB et al. Nitric oxide decreases trimeric CD40 ligand is biologically active. J Biol Chem cytokine-induced endothelial activation. Nitric oxide selectively 1995;270:7025–7028. reduces endothelial expression of adhesion molecules and proinflam-[18] M ach F, Schonbeck U, Sukhova GK et al. Functional CD40 ligand matory cytokines. J Clin Invest 1995;96:60–68.
766 F. Mulhaupt et al. / Cardiovascular Research 59 (2003) 755–766
[38] K wak BR, Myit S, Mulhaupt F et al. PPARgamma but not [41] S himizu K, Schonbeck U, Mach F, Libby P, Mitchell RN. Host PPARalpha ligands are potent repressors of major histocompatibility CD40 ligand deficiency induces long-term allograft survival and complex class II induction in atheroma-associated cells. Circ Res donor-specific tolerance in mouse cardiac transplantation but does 2002;90:356–362. not prevent graft arteriosclerosis. J Immunol 2000;165:3506–3518. [39] K obashigawa JA, Katznelson S, Laks H et al. Effect of pravastatin [42] L arsen CP, Elwood ET, Alexander DZ et al. Long-term acceptance on outcomes after cardiac transplantation. N Engl J Med of skin and cardiac allografts after blocking CD40 and CD28
1995;333:621–627. pathways. Nature 1996;381:434–438.
[40] W enke K, Meiser B, Thiery J et al. Simvastatin reduces graft vessel disease and mortality after heart transplantation-a four year random-ized trial. Circulation 1997;96:1398–1402.