Endothelium-derived nitric oxide: the endogenous nitrovasodilator in the human cardiovascular system
Texte intégral
Figure

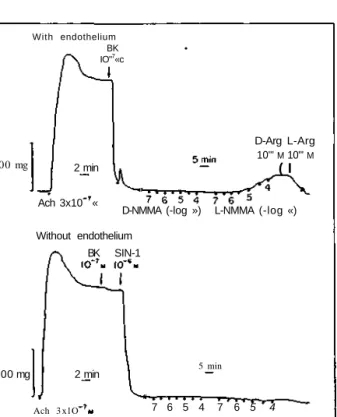
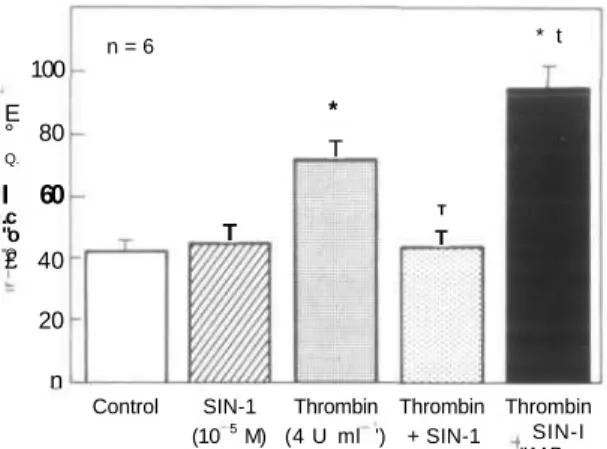
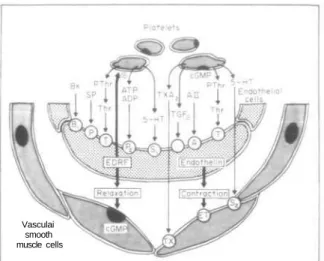
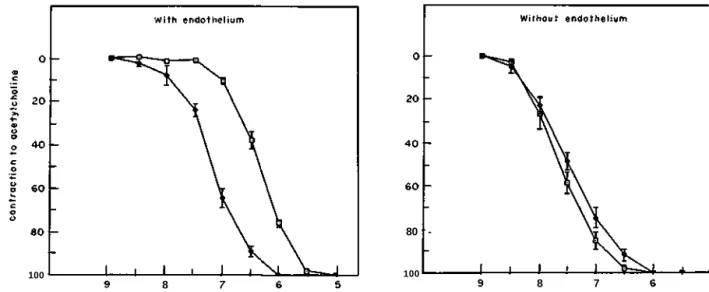
Documents relatifs
Put differently, gain score analyses and analyses using the pretest data as a covariate yield the same, correct estimation of the treatment effect on average, but ANCOVA has
Ainsi, pour limiter la durée du calcul d’optimisation et adapter le résultat au but recherché (la simulation des essais de fatigue), nous avons choisi d’utiliser quatre
Les matériaux ont été compactés statiquement pour obtenir des éprouvettes cylindriques 5×10 cm pour les mesures de conductivité hydraulique au triaxial et 1,5×2 cm
apices quite often not or only slightly digitate (Fig. The life cycle appears to be variable. In BRIP 7543/6045 and BRIP 6063/7548, spermogonia were associated with aecia or with
Depuis leur création dans les années quatre-vingt, les contrats aidés ont régulièrement changé de nom ou de modalités, ciblant plus ou moins et successivement le secteur marchand
Offres d’emploi collectées par Pôle emploi rapportées aux entrées à Pôle emploi (séries CVS) Évolution trimestrielle des entrées à Pôle emploi (CVS, %) Évolution
A labora- tory flow-through experiment was performed to test whether these suspended particles have an influence on the fitness of Daphnia hyalina from Lake Brienz, measured as
