Diversity within American cassava germ plasm based on RAPD markers
Texte intégral
Figure



Documents relatifs
We aimed to determine the diagnostic performance of several proxy meth- ods: the spectrophotometer, iCheck Carotene, and near-infrared spectroscopy using both a desktop (dNIRS) and
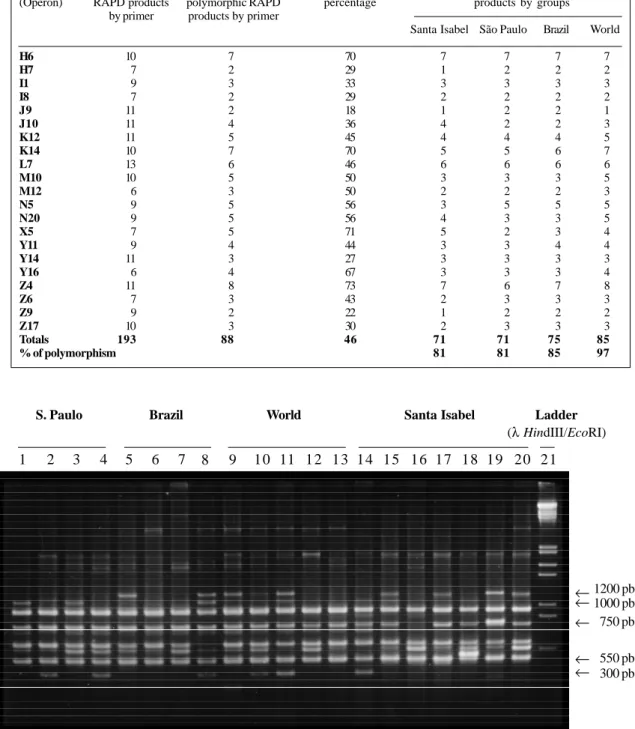
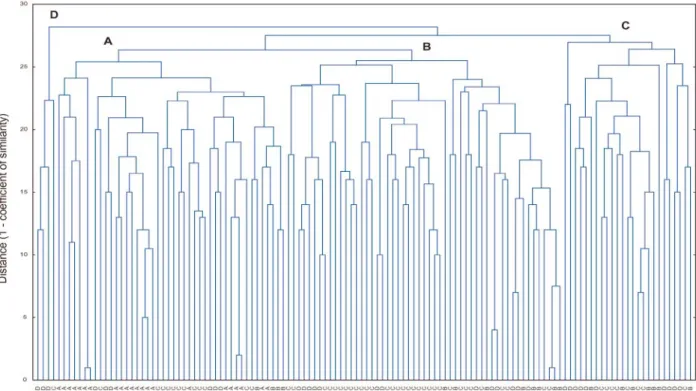
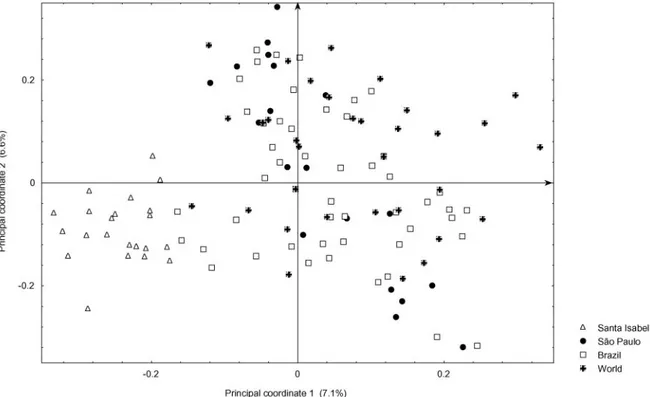
Trials with 50 cassava genotypes (Manihot esculenta Cranzt) from the genetic diversity collection of CIAT (Cali, Colombia) were used to assess the diversity of
178 cassava genotypes (Manihot esculenta Cranzt) from the genetic diversity collection of CIAT (Cali, Colombia) were grown and harvested to assess the diversity of cyanide contents
Cereal and grass forages Legume forages Forage trees Aquatic plants Other forage plants Cereal grains and by-products Legume seeds and by-products Oil plants and by-products Fruits
In common carp (Cyprinus carpio L.) a diet containing 47% cassava root meal has a slightly lower energy digestibility (87 vs. 90%) than diets containing maize or wheat starch..
The case study 3 of RTB project on cassava has for objective to ensure that products resulting from new varieties and from both current and new processing technologies meet
With the support of HarvestPlus, CIAT and other institutions developed and implemented a rapid cycling recurrent selection scheme to increase total carotenoids content (TCC) and
The Distance Kernel Method (DKM) overcomes this issue by clustering the realizations in a multidi- mensional space based on the flow responses obtained by means of an
