Communication Biophysics
Texte intégral
Figure




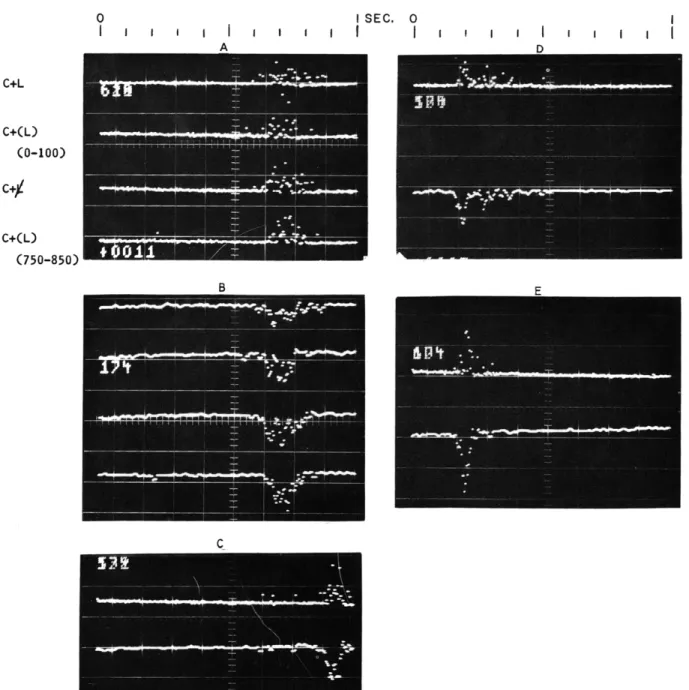
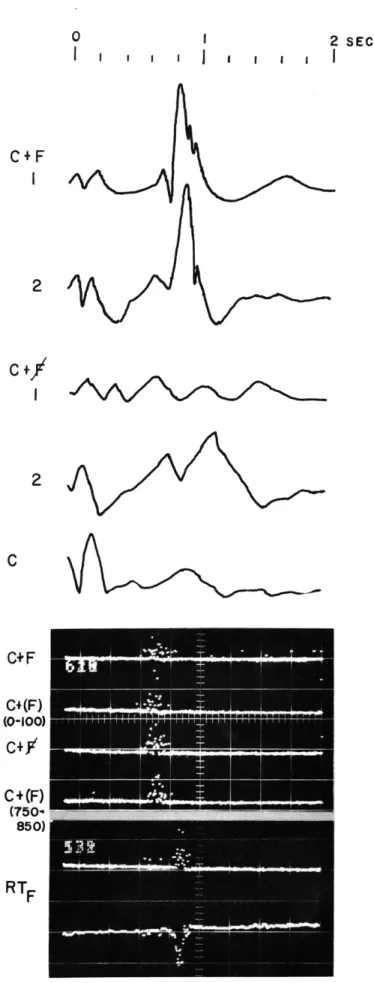
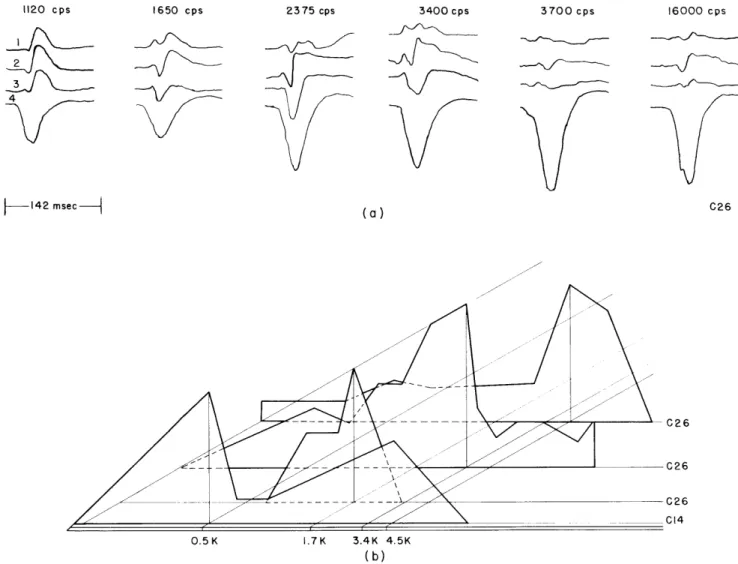
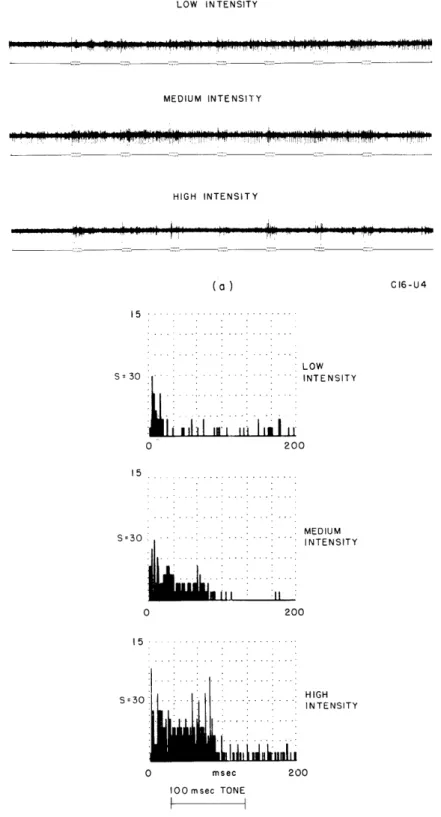
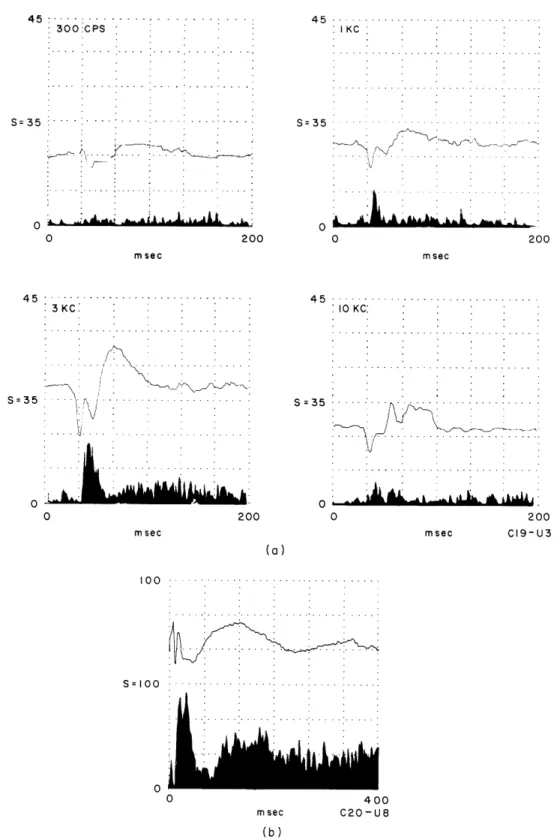
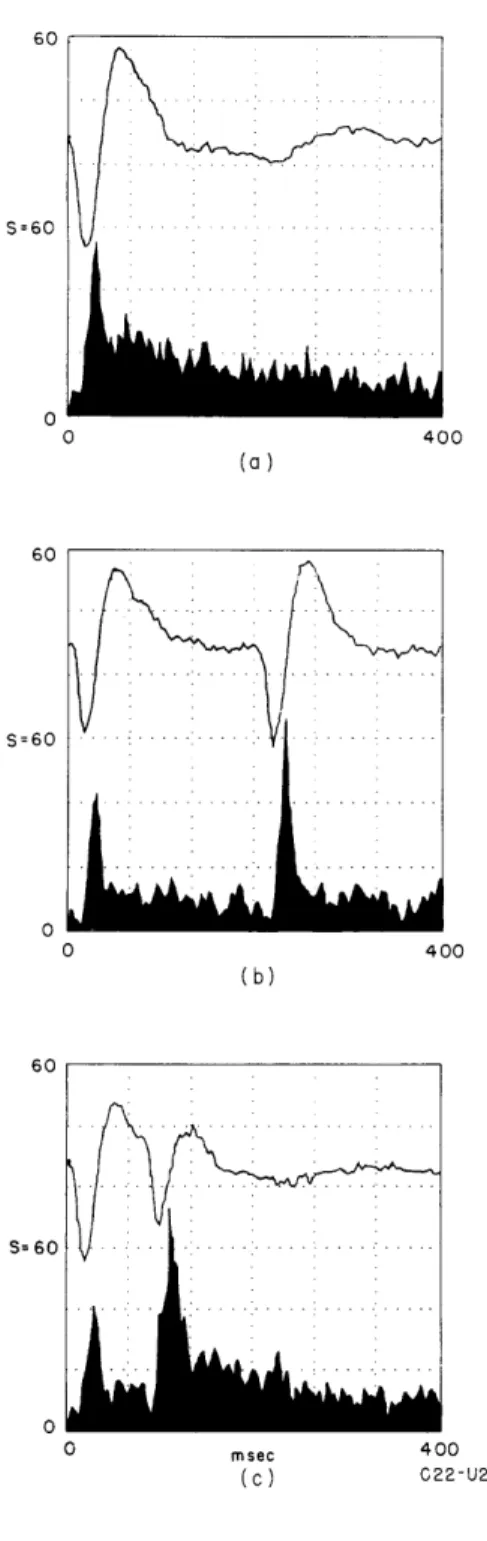
Documents relatifs
Un système mécanique oscillant est un système dont le mouvement du centre d’inertie G est périodique de va-et-vient au tour d’une position d’équilibre..
Sketch in the Brusin’s excavation site diary, illustrating the system of land consolidation and underpinning for the outer curtain wall (Courtesy of Museo Archeologico Nazionale
[r]
On emploie le ball socket pour connecter les chaînes d’isolateurs type rotule et logement de rotule avec les pinces d’ancrage et suspension.. Les dimensions du tige sont d’accord
2- relie les grandeurs à
Conclure : si la même charge peut être répartie de différentes façons et si le critère de dimensionnement de la poutre est la contrainte maximale, quelle sera la valeur de a
The paper was devoted to the analysis of the control properties of the structurally damped 1D wave equation on the interval I = (0, 1), the control acting at the point x = 1.. It
Les enzymes sont des protéines qui agissent comme catalyseurs dans les cellules des vivants en abaissant l’énergie et des particules de substrat se rassemblent de façon
