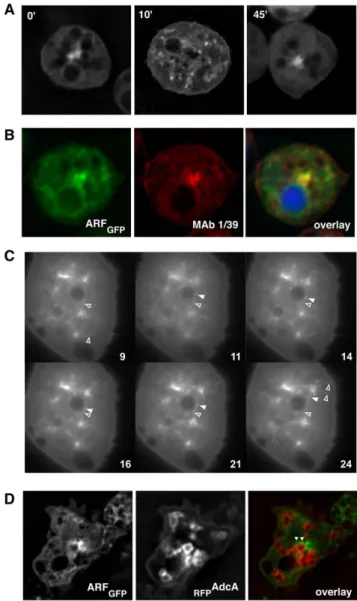
FYVE-dependent endosomal targeting of an arrestin-related protein in amoeba.
Texte intégral
Figure
Documents relatifs
The proof of the central limit theorem for the position of the tagged particle in the asymmetric exclusion process relies on the dual spaces and operators introduced in Section
We used kinetic data on three variables of the model: (i) previously published angiotensin- induced ERK phosphorylation data (Kim et al, 2005), (ii) original DAG accumulation and
Monsieur le médecin commandant Younes Moutaki Allah Professeur Assistant de la chirurgie cardio-vasculaire De l’hôpital militaire d’instruction Mohammed V Rabat. Nous vous
A l’avenir, de nouvelles études seront nécessaires afin d’évaluer la pertinence des différences prosodiques entre les variétés dialectales étudiées ici (alignement
Here we present a 2.75 Å crystal structure of ArrFL-1, a peptide analogue of the finger loop of rod photoreceptor arrestin, in complex with the prototypical GPCR rhodopsin..
Using YFP and Rluc fusion constructs expressed in COS-7 cells, BRET data revealed a pre-assembly of PAR2 with both G αi1 and Gαo and a rapid and transient acti- vation of these
These different results show that the epsins Ent3 and Ent5 are required for the endosomal sorting of internalized ubiquitylated cargos and that Btn3 is also involved in this
Further, the range of unstable wave number increases with an increase in the thickness of the porous layer. The film flow system is more unstable for a film over a thicker porous
