Hyperosmolar environment and intestinal epithelial cells: impact on mitochondrial oxygen consumption, proliferation, and barrier function in vitro
Texte intégral
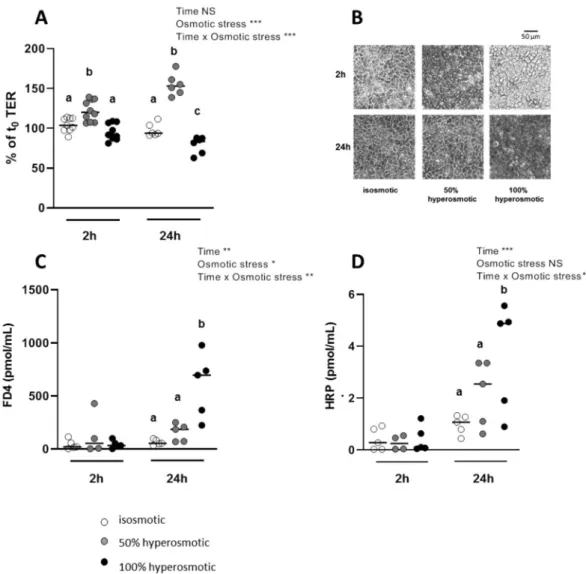
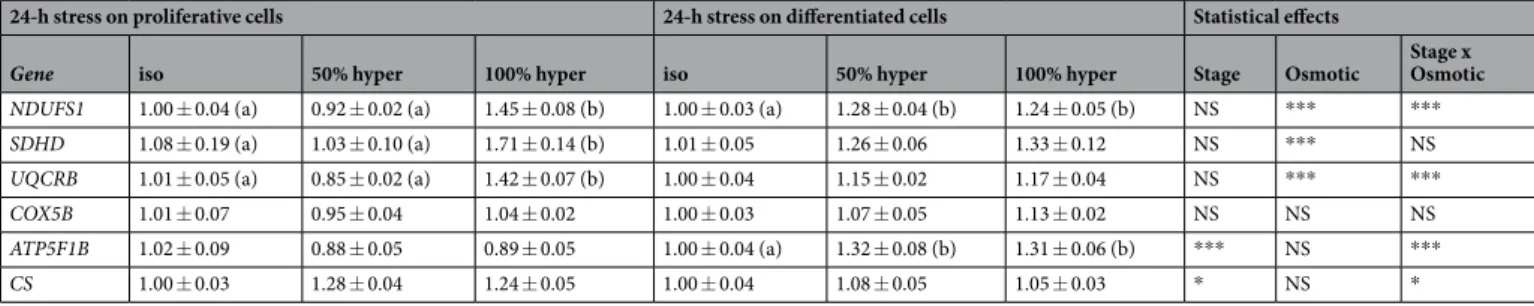
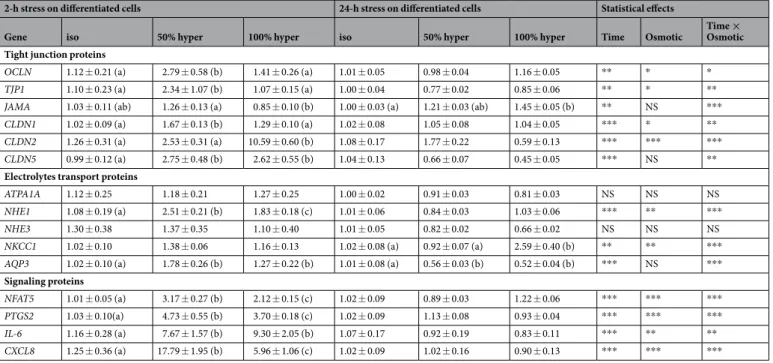
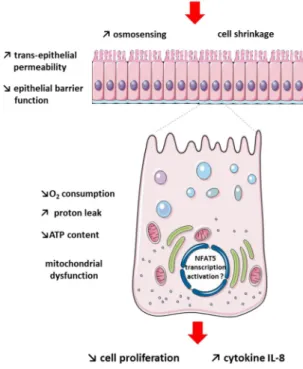
Figure
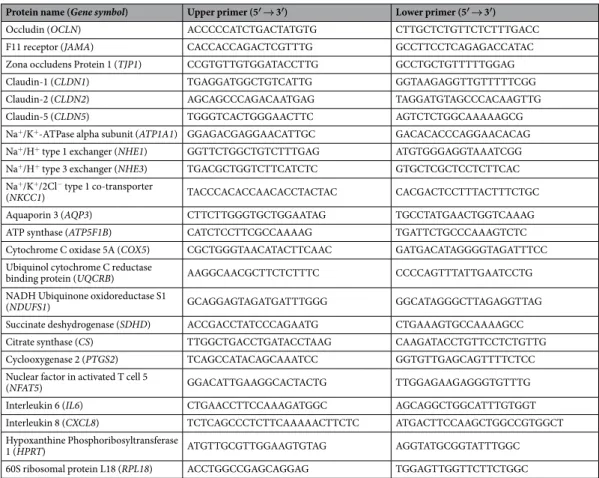
Documents relatifs
— Les consommations d'oxygène de deux espèces de Palourdes, Rudi- tapes decussatus et Ruditapes philippinarum, ont été mesurées sur de jeunes individus nés en
Stoichiometric coefficients and thermodynamic equilibrium constants for minerals and aqueous species were taken from the EQ3/6 database (Wolery, 1992), though only a
Using a homologous in vitro binding assay, we could show that, as predicted, the carboxyl-terminal domain of trypanosomal IF2 was able to specifically bind the elongator tRNA Met
Certain endpoints can have a direct bearing on chemical substances management (e.g., REACH criterion for very bioaccumulative substances: BAF >5000) as well as on the
Innovation: In human epithelial corneal and conjunctival cells of the ocular surface, we demonstrated the harmful impact of blue light on viability, redox state and
α la, α -lactalbumin; β lg, β -lactoglobulin; cAMP, cyclic adenosine 3 ′ ,5 ′ -monophosphate; CLA, conjugated linoleic acid; CMC, critical micelle concentration; DMEM, Dulbecco ’
But beyond the child itself who is immersed in mundane consumption practices and permanently confronted with commercial persuasive discourses, the field of child and teen
Ce que nous proposons dans cette dernière partie, c’est d’aller plus loin que les travaux précédents et de suggérer que la consommation, dans sa prise en compte stratégique
