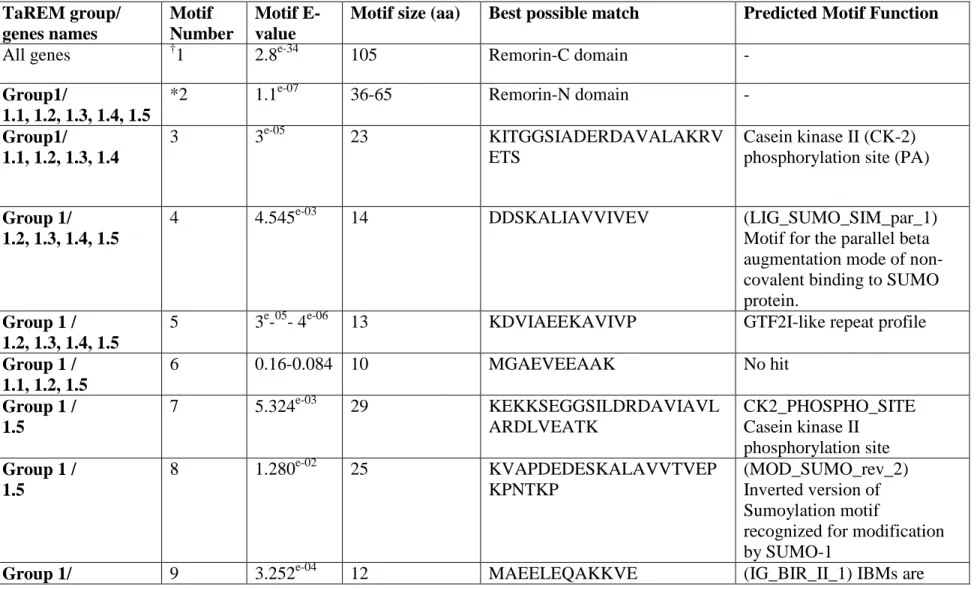
Table S2. Identificaton of consensus sequence of TaREM using MEME and their function by prosite and Eukaryotic Linear Motif resource programs.
Texte intégral
Figure
Documents relatifs
The principal underlying differ- ence between the two methods is that DILIMOT masks all but one arbitrarily selected homologous protein prior to motif discovery, whereas
In the last months the WP3 group, considering the present status of the R&D programs at different laboratories, and following the requirements of a XADS accelerator
Jean-Jacques Rousseau‟s critical rewriting in his Nouvelle Héloïse of two foundational medieval works – the letters of Abélard and Héloïse, and Petrarch‟s Canzoniere –
Recovered motifs from multi-camera – For space rea- sons, Fig. 20 shows only the 6 most probable motifs recovered by our algorithm when setting the prior on the average maximum
The original analysis by TIGR provided 625 function predictions (Bult et al., 1996) (37% of the chromosomal genome), with the update increasing them to 809 assignments (Kyrpides et
Finally, the catalytic properties of the mutant enzyme were examined by testing its ability to form a covalent complex with the FC-mini-RNA substrate analog (Fig. 4C) and to
We propose a verification approach guided by the requirement relationships, such as ``verify'', with the model elements, to verify functional requirements with state machine
Entringer numbers and case when q is a power of 2 Formulating a conjecture analogous to Conjecture 1 for powers of 2 requires to define, following Arnold [Arn91], a sequence
