Master 2 Recherche | Parcours Biodiversité, Ecologie et Evolution
MASTER SET
Sciences de l’Environnement Terrestre
Année 2015 - 2016
Analyse qualitative et quantitative de la
production de semences du sapin pectiné
(Abies alba) en région méditerranéenne
française et détermination des facteurs
environnementaux associés
Présenté par Célia MARTINEZ
Dr Christian Pichot et Dr Thomas Boivin, Maîtres de stage
Remerciements
Tout d’abord ce stage n’aurait pas été possible sans les données et les financements des projets ECOGER, ANR DRYADE et GRAAL, ainsi que les subventions de fonctionnement de l’état et du ministère de l’agriculture et de la pêche.
Je tiens à remercier dans un premier temps mes deux tuteurs, Christian Pichot et Thomas Boivin, pour m’avoir fait découvrir ce sujet d’étude et pour le temps qu’ils m’ont consacré tout au long de cette période. Je leur suis également reconnaissante pour m’avoir transmis un peu de leurs connaissances, pour les conseils toujours avisés de Thomas et l’aide précieuse en statistiques de Christian.
Merci à Eric Rigolot de m’avoir accueilli au sein de l’unité et à l’ensemble de l’équipe Ecologie des Forêts Méditerranéennes pour leur accueil et leur bonne humeur toujours au rendez-vous et plus.
Je remercie Alex Baumel, coordinateur du Master 2 BEE, pour m’avoir accompagnée dans la recherche de stage, ainsi que toute l’équipe pédagogique du Master 2 BEE pour m’avoir préparée à cette expérience.
Merci à Mehdi Pringarbe de m’avoir montré les manipulations en laboratoire et d’y avoir passé du temps en me faisant partager ses goûts en musique classique, et Alain Chalon, toujours disponible pour faire une petite radiographie à rayon X de dernière minute.
Je remercie également Anne Latreille, doctorante, pour son aide et ses conseils avisés, ainsi que tous les autres doctorants et stagiaires pour votre humour et votre bonne humeur.
Merci également à Clara Tallieu, stagiaire et voisine, d’avoir partagé ce bureau avec moi pendant six mois, d’avoir refait le monde pendant le déjeuner, d’être toujours là pour m’écouter et pour sa bonne humeur de tous les jours.
Merci à Juliette Cornille, notre petit rayon de soleil, qui a partagé notre bureau dans les moments les plus difficiles de ce stage sans se défaire de son sourire.
Spéciale dédicace à Maurane Buradino, pour avoir toujours le mot pour rire dans n’importe quelle situation.
Table des matières
I. Introduction 1
II. Matériels et Méthodes 4
1. Le système modèle : le sapin pectiné 4
2. Les dispositifs expérimentaux 6
a) Suivis de peuplements à l’échelle régionale 6
b) Suivis des individus répartis sur un gradient altitudinal 6
3. Caractérisation de variables de reproduction et de l’environnement 7
a) Les variables individuelles de reproduction 7
b) Les variables topographiques 7
c) Les variables climatiques 7
4. Estimations qualitatives et quantitatives de la production de graines 8
a) Echantillonnage 8
b) Extraction et tri des graines 8
c) Estimation de la qualité des graines par radiographie digitale à rayon X 8
5. Analyses statistiques 9
a) Analyse de la variabilité de la quantité et de la qualité de la production de graines 9
b) Analyse des effets des facteurs environnementaux 10
III. Résultats 12
1. Caractérisation climatique des sites étudiés 12
a) Dispositifs régionaux 12
b) Dispositifs en gradients altitudinaux 13
2. Etude de la variabilité de la quantité et de la qualité de graines produites 14
a) Dispositifs régionaux 14
b) Dispositifs en gradients altitudinaux 15
3. Influence des facteurs climatiques sur les variables étudiées 17
IV. Discussion 19
V. Bibliographie 23
Table des annexes 25
1 I. Introduction
Les écosystèmes forestiers sont aujourd’hui touchés par les modifications du climat à l’échelle mondiale. L’augmentation de la fréquence et de l’intensité des sécheresses et des stress pour la physiologie des arbres qui en résultent pourrait modifier fondamentalement la composition, la structure et la biogéographie des forêts dans de nombreuses régions (Allen et
al. 2010). Les conditions climatiques prévues au cours des prochaines décennies devraient
induire des modifications dans la plasticité et l’évolution des espèces et conduire à des changements dans leur distribution géographique. Les populations en marge de l’aire de distribution, qui sont particulièrement sensibles aux changements de leur environnement, sont attendues pour être les premières affectées (Morin et al. 2008). Les impacts du changement climatique ont été étudiés sur de nombreux processus biologiques tel que la phénologie (Parmesan et Yohe 2003; Root et al. 2003) ou la croissance (Cailleret et Davi 2011). Les composantes de la dynamique spatiale et temporelle telles que la reproduction ont très probablement un rôle clé dans la réponse de ces espèces aux changements climatiques, qui réside principalement dans leur capacité de d’adaptation et/ou de migration (Clark 1998; Sagnard et al. 2002).
La production de semences est déterminante pour la colonisation de nouveaux milieux mais aussi la survie à long terme des populations d’arbres (Sagnard et al. 2002). Acquérir des connaissances sur les facteurs qui contrôlent la production de semences, en quantité et en qualité, est cruciale aujourd’hui pour prévoir la distribution potentielle des espèces au cours des prochaines décennies. Cette production joue un rôle significatif dans les processus de régénération naturelle (Mencuccini, et al. 1995), elle influe sur le fonctionnement des écosystèmes, peut être assimilée à un puit de carbone non négligeable et constitue un flux de matière important (Fenner 1987).
La production des semences chez les conifères implique une longue séquence d’évènements développementaux qui peut durer jusqu’à trois ans (Tomlinson et Takaso 2002). Ainsi, la formation des graines résulte du développement des ovules après fécondation, les graines contiennent un embryon, accompagné ou non dans un tissu de réserve, qui est entouré par des téguments protecteurs (Côme 1982). La transformation des ovules en graines est régulée par des facteurs endogènes et par les conditions environnementales (Owens 1991; Philippe et
al. 2006).
Les arbres forestiers peuvent produire des quantités de graines irrégulières à l’échelle interannuelle (Philippe et al. 2006). Par exemple, la production de graines de l’épinette de Norvège diminue chaque année à cause des changements climatiques (Mencuccini et al. 1995).
2
Des variations dans le nombre de graines produites peuvent apparaître d’une à l’autre et certaines études évoquent une tendance modeste pour qu’une année de production abondante de semences soit suivie par une année de production anormalement faible (Greene et Johnson 2004). Les facteurs déterminant cette production sont souvent difficiles à démêler. Ainsi des facteurs biotiques et abiotiques peuvent perturber ce développement (Philippe et al. 2006).
Du fait de leur longévité, les arbres ont la capacité de participer à de nombreux épisodes de reproduction au cours de leur vie, leur fertilité et donc leur succès reproducteur peuvent varier en fonction des conditions climatiques. De plus, les variations interannuelles de la fertilité sont plus importantes pendant les années où la floraison a été mauvaise et pour les jeunes arbres (Kang et al. 2003). La quantité de semences disponibles dépend aussi du nombre et de la taille des cônes (Calama et Montera 2007) mais la production de graines peut également être réduite avec l’augmentation de l’altitude. Ces variations résultent souvent d’un ensemble de facteurs climatiques et édaphiques (Mencuccini et al. 1995), la température, les précipitations et l’humidité relative de l’air étant des facteurs susceptibles de contraindre ou de favoriser plusieurs phases du cycle de reproduction. Par exemple on associe souvent de faibles précipitations printanières ou estivales à de bonnes floraisons (McNeill 1954; Matyas et Giertych 2013) ou encore de fortes sécheresses peuvent provoquer des anomalies du développement embryonnaire (Owens et Blake 1986). Des facteurs biotiques peuvent aussi intervenir pendant le développement des graines. Les dégâts sont causés par des insectes ravageurs attaquant après la fécondation ou par le développement de larves issues d’œufs pondu avant ce stade, comme le genre Megastigmus. Ils peuvent causer des destructions directes, lorsque les larves se nourrissent de la graine, ou par des dégâts indirect, lorsqu’elles dévorent les tissus du cône (Turgeon et al. 1994; Boivin et al. 2016).
Les mécanismes de production de semences a été principalement étudiés dans le cadre de l’amélioration de la production en vergers à graines (Philippe et al., 2006), et dans la plupart des études les travaux sur les facteurs influençant la fructification se sont focalisées sur la quantité de graines (Koenig et Knops 2000; Calama et Montera 2007), alors que la qualité des graines et leur potentiel germinatif restent un élément majeur du succès reproducteur d’un individu. Des approches uniquement quantitatives de la reproduction des arbres ne sont donc pas suffisantes pour comprendre et prédire la dynamique spatio-temporelle des arbres, mais l’accès non destructif aux données qualitatives nécessite des outils méthodologiques souvent sophistiqués et coûteux (radiographie à rayons RX par exemple).
Dans ce contexte, le but de cette étude est de déterminer tout d’abord comment varie, quantitativement et qualitativement, et dans le temps et dans l’espace, la production de
3
semences dans des populations d’arbres situées en marge sud de l’air de distribution de l’espèce. Puis nous recherché les facteurs environnementaux (biotiques et abiotiques) explicatifs de ces variations.
L’espèce Abies alba a été choisie comme modèle d’étude pour répondre à cette problématique. Le sapin pectiné est présent dans les régions montagneuses de l’Europe. Son aire de distribution s’étend du Nord de l’Allemagne (52°N) au sud de l’Italie (38°N), de l’est de la Roumanie (22°E) au Pyrénées occidentales en France (03°W). Dans la partie ouest de son aire de distribution Abies alba fait face à des variations biogéographiques (de l’intérieure des Alpes vers l’extérieure), des variations bioclimatiques et différentes conditions édaphiques (Ozenda 1981). Son aire de répartition est très fragmentée dans les Alpes méditerranéennes et constitue seulement 3,5% de la superficie totale que recouvre le sapin pectiné en France (Sagnard et al. 2002). Dans cette région, les populations de sapin pectiné sont soumises à des contraintes de température et d’humidité importantes, dont les conséquences en termes de croissance, de survie et de dynamique sont déjà significatives (Restoux 2009).
Dans un premier temps nous nous sommes intéressés aux différentes variations spatiales et temporelles de la production de semences dans les populations méditerranéennes de sapin pectiné. Puis dans un second temps nous nous sommes intéressés aux facteurs individuels et environnementaux, comme la température, les précipitations et l’humidité relative de l’air, pouvant expliquer ces variations. Pour répondre à notre problématique nous avons développé une approche à la fois quantitative et qualitative reposant sur des historiques de production de semences, ainsi que des données individuelles (arbres semanciers), climatiques et topographiques. Les données issues de deux types de dispositifs ont été étudiées. Un premier dispositif régional a permis d’étudier les variations de la production de semences entre peuplements, répartis dans toute la région du Languedoc Roussillon, de 2004 à 2009. Le second dispositif a permis l’étude de la production de semences des arbres de trois gradients altitudinaux sur trois sites en région Provence-Alpes-Côte d’Azur, le mont Ventoux, la forêt de la Vésubie et la forêt de l’Issole, ceci entre 2007 et 2013. Nous avons également fait l’acquisition de données topographiques et climatiques pour répondre à l’hypothèse concernant l’impact de ces facteurs sur la production de semences.
4 II. Matériels et Méthodes
1. Le système modèle : le sapin pectiné
Le sapin pectiné est un conifère de la famille des Pinaceae du genre Abies. Il est le plus grand arbre du genre Abies en Europe et peut ainsi atteindre jusqu’à 60-65m de hauteur et jusqu’à 150 à 200cm de diamètre. Il tolère un large éventail de conditions de sol, tant pour la teneur et la disponibilité en éléments nutritifs que pour le pH mais il est plus exigeant en terme d’humidité et de température (Wolf 2003). Il est notamment très sensible aux sécheresses estivales, aux faibles humidités et aux gelées de printemps (Sagnard et al. 2002)
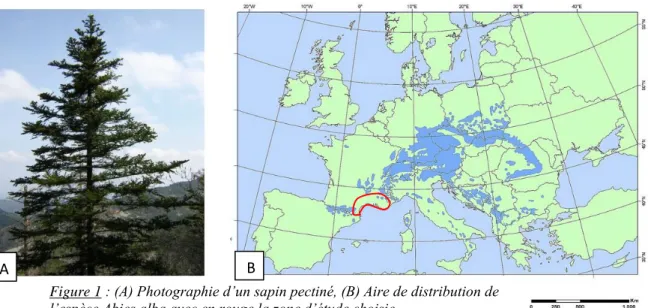
Le sapin pectiné est présent dans les régions montagneuses de l’Europe. Son aire de distribution (figure 1B) s’étend de Varsovie en Pologne comme limite nord, jusqu’en méditerranée comme limite sud (Sagnard et. al. 2002).
C’est une espèce monoïque, les inflorescences femelles et mâles sont présentes simultanément sur un même individu et anémophile (dispersion du pollen par le vent). Le cycle de vie d’A. alba comprend une longue période juvénile dans laquelle il ne produit pas de graines. Son âge de première maturité se situe entre 25 et 35 ans pour les arbres isolés et entre 60 et 70 ans pour les arbres en pleine forêt.
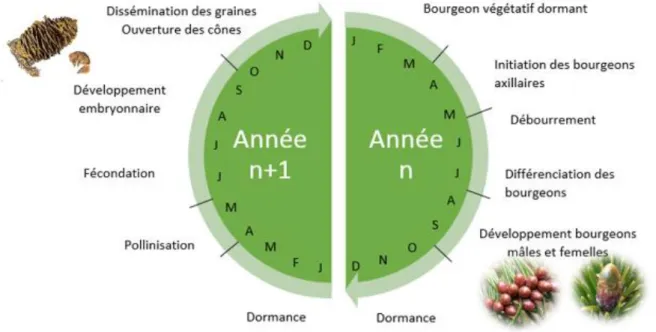
Son cycle reproducteur (Fig. 2) dure environ 1 an et demi, il débute pendant l’année n au printemps avec l’initiation florale et se termine en automne pendant l’année n+1 avec l’ouverture des cônes et la libération des graines. Pendant l’année n l’initiation florale est suivie du débourrement puis de la différenciation florale, cette étape est sensible au climat, des températures élevées pendant les premiers stades de formation des ébauches florales peuvent favoriser la floraison (Matthews 1963). Inversement, de très basses températures peuvent
Figure 1 : (A) Photographie d’un sapin pectiné, (B) Aire de distribution de l’espèce Abies alba avec en rouge la zone d’étude choisie.
5
endommager les bourgeons mâles pendant leur développement qui se termine en septembre de l'année n. Après la phase de dormance le gamétophyte femelle finit son développement pendant les mois de mai et juin. Juste avant la fécondation, la phase de pollinisation peut être perturbée par de fortes précipitations qui lessiveront les cônes ou par de faibles températures qui sont susceptibles de détruire les jeunes inflorescences (Philippe et al. 2006). Tout comme les graines, les cônes ont presque atteint leur taille définitive au moment de la fécondation (année n+1). C’est pendant les mois de juin, juillet et aout que la graine se développera et ce stade peut ainsi être perturbé par des sécheresses estivales dues à de faibles précipitations et de fortes températures. Dans les cônes récoltés, on peut trouver quatre types de graines. Tout d’abord les graines de petites tailles qui sont avortées et qui résultent d’un avortement précoce dû à une absence de pollinisation. Deuxièmement, les graines vides qui sont le résultat d’un avortement embryonnaire post-pollinisation dû à une incompatibilité embryon/endosperme généralement lié à de l’auto-fécondation. Troisièmement, les graines saines correspondent aux graines ayant eu un développement embryonnaire complet (Philippe et al. 2006). Et quatrièmement, les graines parasitées qui présentent des larves de Megastigmus se nourrissant de la graine pour son développement (Turgeon et al. 1994 ; Boivin et Auger-Rozenberg 2016).
6
2. Les dispositifs expérimentaux
a) Suivis de peuplements à l’échelle régionale
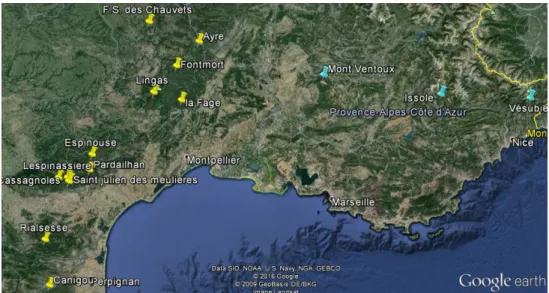
Dans le cadre d’une étude sur la qualité sanitaire de la production de graines d’A. alba, l’équipe d’Ecologie et dynamique des populations d’insectes forestiers de l’INRA d’Avignon a estimé différents paramètres faisant état de la qualité de la production des graines (la proportion de graines produites par cônes, le taux de graines saines, le taux de graines vides et les taux d’infestation par les insectes séminiphages du genre Megastigmus) (Candau et al. 2009). Cette étude a été menée, de 2004 à 2009, sur 11 peuplements situés en Languedoc Roussillon répartis aléatoirement dans les 5 départements de la région Tous les peuplements étudiés en Languedoc Roussillon sont des sapinières pures bordées par des peuplements de cèdres.
b) Suivis d’individus répartis sur un gradient altitudinal
Dans le cadre de projets Dryade et Ecoger au sein de l’URFM de l’INRA d’Avignon, 3 gradients altitudinaux ont été mis en place sur le versant nord de 3 massifs forestiers, le Mont Ventoux, la forêt de l’Issole et la forêt de la Vésubie. Ces trois sites sont gérés par l’Office Nationale des Forêts. Chaque site présente des zones de sapinières pures ainsi que des zones mixtes. L’essence dominante sur Issole et Vésubie est A. alba, associé à un peu de Larix decidua, de Picea abies et de Pinus sylvestris. On compte trois autres essences majoritaires sur le mont Ventoux : Fagus
sylvatica, Pinus sylvestris et Sorbus aria.
L’échantillonnage du gradient altitudinal du Mont Ventoux a été effectué le long d’un transect divisé en deux parties du fait de la topographie des lieux : un transect bas (de 970 à 1139m) et un transect haut (de 1241 à 1524m).
Figure 3 : Localisation des sites d’expérimentation en région méditerranéenne française, en jaune les suivis à l’échelle régionale, en bleu les suivis sur un gradient altitudinal.
7
En Issole et en Vésubie, l’échantillonnage a été effectué au sein de 5 placettes réparties sur les transects altitudinaux suivants : de 1092m jusqu’à 1568m (Issole) et de 1058m à 1501m (Vésubie).
3. Caractérisation de variables de reproduction et de l’environnement a) Les variables individuelles de reproduction
La création de 2 jeux de données a été nécessaire car les variables mesurées diffèrent entre les deux dispositifs.
Pour les peuplements étudiés à l’échelle régionale, les variables étudiées sont : le nombre de graines par cône, le taux de graines saines, le taux de petites graines, le taux de graines parasitées et le taux de graines vides.
Pour les peuplements et les arbres provenant des gradients altitudinaux, les variables étudiées sont : le taux de graines saines, le taux de graines vides et le taux de graines parasitées, la masse de mille graines (PMG), ainsi que la longueur et la masse de chaque cône.
L’âge, la circonférence et le nombre de cônes mâles de chaque arbre ont été obtenus à partir des bases de données d’archives sur ces mêmes sites créées dans le cadre d’autres projets de recherche à l’URFM.
b) Les variables topographiques
Ces variables correspondent à l’altitude de chaque peuplement et de chaque niveau altitudinaux de gradients mis en place. Dans le cadre d’un travail de thèse sur la diversité et l’adaptation locale des arbres à l’URFM (Anne Latreille), la densité des peuplements (nombre d’individus/ha) sur le Mont Ventoux, Issole et Vésubie ont aussi été calculées.
c) Les variables climatiques
Les données journalières de températures (minimale, maximale, moyenne), de pluviométrie et d’humidité relative ont été obtenues à partir de base de données Météo France. Les stations ont été choisies en fonction de leur distance aux sites de suivis. Parfois la station considérée comme étant la plus proche voisine est la même pour deux ou trois sites. C’est pourquoi les données de températures de chaque site ont nécessité une interpolation basée sur la méthode d’interpolation AURELHY (Analyse Utilisant le Relief pour l’Hydrométéorologie). Cette méthode utilise l’information « relief » pour améliorer la cartographie des précipitations et des températures en s’appuyant sur un modèle numérique de terrain fourni par l’I.G.N. (Institut Géographique National).
8
Des données climatiques provenant de micro stations météorologiques (HOBO) installées sur différents niveaux de chaque gradients altitudinaux ont permis de récolter des informations précises sur la température et l’humidité relative journalières le long de chaque gradient. Les données climatiques ont été choisies en fonction de périodes d’intérêt dans le processus de formation de la graine ont été distingué. Ainsi, les températures minimales (TN), les températures maximales (TX), les précipitations (RR) moyennes et l’humidité relative (HR) moyenne pendant la période de pollinisation des cônes (le mois de mai) ont été calculées pour chaque année, chaque site et chacun des niveaux altitudinaux des gradients. De la même façon les températures maximales et les précipitations moyennes pendant la période de développement de la graine ont été calculées (les mois de juin, juillet et août). L’intensité de la sécheresse a été quantifiée par le calcul du déficit de précipitations cumulée pendant l’été, ce déficit correspond aux millimètres d’eau nécessaires pour compenser les températures estivales plus fortes.
4. Estimations qualitatives et quantitatives de la production de graines a) Echantillonnage
Pour les deux dispositifs, les cônes ont été collectés à la fin du mois d’Août. A cette période, les graines ont atteint leur maturité mais n’ont pas encore été libérées des cônes. Le dispositif régional a été créé pour un suivi à l’échelle du peuplement contrairement au dispositif en gradients altitudinaux qui a été créé pour un suivi individuel, ce qui explique les différences de types d’échantillonnage entre les deux dispositifs.
Dans le dispositif régional, trois cônes ont été prélevés sur dix arbres choisis de façon aléatoire dans chaque peuplement. Les arbres récoltés diffèrent donc entre les années au sein d’un même peuplement.
Pour le dispositif en gradients altitudinaux, dix cônes ont été prélevés sur chaque arbre reproducteur sélectionné, qui a été identifié et suivi chaque année. Tous les arbres ne fructifient pas forcément tous les ans donc le prélèvement des cônes s’est fait en fonction des arbres reproducteurs chaque année, ainsi le nombre d’arbres prélevés peut différer entre année. Sur les dix cônes prélevés par arbre l’analyse des graines s’est faite uniquement sur trois cônes (un moyen, un gros et un petit), due à des contraintes temporelles et humaines pour traiter l’ensemble des récoltes.
9 Figure 4 : Lots de graines et mises en plaque avant la radiographie RX.
b) Extraction et tri des graines
Après la récolte des cônes, ceux-ci ont été désarticulés manuellement au laboratoire puis on en extrait les graines. Si les graines ne sont pas analysées tout de suite elles sont conservées dans un congélateur à -19°C. Le cadre de la présente étude a donné lieu à la réalisation d’un test de germination visant à évaluer l’effet de la décongélation/recongélation.
Une fois extraites, les graines provenant du dispositif à l’échelle régional ont été triées et comptées selon trois catégories : les petites graines, les grandes graines abimées et les grandes graines en bon état.
Les cônes provenant des dispositifs en gradients altitudinaux ont été pesés et mesurés avant d’être désarticulés. Pour chaque arbre la masse de 1000 graines a été mesuré mais les graines n’ont pas fait l’objet d’un tri préalable.
c) Estimation de la qualité des graines par radiographie digitale à rayons X
Après avoir été désailées, les grandes graines en bon état ont été disposées sur des plaques transparentes pour permettre leur radiographie (figure 4) (Faxitron-MX20 et scanner digital EZ20)
La lecture des images de radiographie à rayons X (logiciel Mesurim pro) permet d’identifier les trois catégories de grandes graines suivantes : les graines saines, les graines vides et les graines parasitées. Pour faciliter leur comptage, chaque type de graine est associé à une couleur (Figure 5). Les graines saines récupérées ont été ensuite placées dans un congélateur à -19°C pour être conservées.
10
5. Analyses statistiques
Les analyses statistiques ont toutes été réalisées avec le logiciel R 3.1 (GuR).
a) Analyse de la variabilité de la quantité et de la qualité de la production de graines La distribution de chaque variable a d’abord été étudiée. Pour les variables qui représentent des effectifs on utilise, soit un modèle linéaire généralisé qui suit une loi de Poisson, soit un modèle linéaire. Ce choix de modèle est basé sur l’observation de la distribution des résidus estimés, qui peut prendre la forme « < » (loi de Poisson, glm avec une fonction de lien « log ») ou « = » (loi normale). Pour les variables représentant des proportions, un modèle linéaire généralisé associé à la fonction binomiale a été utilisé.
Dispositifs régionaux :
Modèle 1 = variable choisie ~ sites*année Dispositifs en gradients altitudinaux :
Modèle 2 = variable choisie ~ sites*année + niveaux d’altitudes + individus
Pour chaque variable un test d’analyse de la variance (ANOVA) a été effectué et les prédictions (fonction « predict » de R) de chaque modèle ont permis l’observation graphique des variations entre années, sites et individus.
b) Analyse des effets des facteurs environnementaux
Deux types de modèles ont été utilisés pour déterminer les effets des facteurs environnementaux : un modèle linéaire (variables sous forme de comptages) et un modèle
Graine saine
Graine vide
Graine parasitée
Figure 5 : (A) Radiographie par rayon X, (B) Exemple de comptage de graines avec le logiciel Mesurim pro : en bleu les vides, en vert les saines et en rouge les parasitées (présence d’une larve)
11
linéaire généralisé (variables sous forme de pourcentages). L’effet aléatoire des variables sites et années a été pris en compte en recalculant le résultat du test de Fisher manuellement dans le cadre d’un modèle linéaire et avec l’utilisation de la fonction glmer pour les modèles linéaires généralisés (package lme4). Une analyse de variance, ANOVA, a permis de déterminer si l’un des facteurs avait un effet significatif, de plus la significativité des effets de chaque facteur a été étudiée à partir d’un test de Fisher (pour les modèles linéaires) et d’un test de Chi2 (pour les
modèles linéaires généralisés).
Dispositifs régionaux :
Modèle 3 = variable choisie ~ sites*année + RR(pollinisation) + RR(développement de la graine) + TN(polllinisation + TX(développement de la graine)
Dispositifs en gradients altitudinaux :
Modèle 4 = variable choisie ~ sites*année + niveaux d’altitudes + individus + HR(pollinisation) + HR(développement de la graine) + TN(polllinisation + TX(développement de la graine).
12 III. Résultats
1. Caractérisation climatique des sites étudiés a) Dispositifs régionaux.
Les profils climatiques des sites étudiés ont pu être définis à partir des diagrammes ombrothermiques (Fig. 6A et annexe 1) et d’une analyse en composantes principales des variables d’altitude, et de température et de précipitations au cours de différentes phases clés du développement des graines (Fig. 6B).
Les sites Lespinassière (n°8) et Cassagnoles (n°9) présentent deux profils similaires. Ce sont les sites qui ont subi les plus fortes précipitations pendant l’hiver (jusqu’à 160 mm de précipitation cumulée) et l’automne et le printemps (de 90 à 120 mm), mais ils présentent tout de même une période de sécheresse pendant les mois de juin et juillet. Les autres sites du dispositif expérimental régional n’ont pas subi de précipitations dépassant les 100 mm en moyenne au cours de l’année et ne sont donc pas caractérisés par les précipitations. Les sites Canigou (n°2), Espinouse (n°4), Pardailhan (n°10) et Rialsesse (n°11) sont corrélées avec la sécheresse estivale et présentent un déficit de précipitations cumulées moyen entre 28,15 mm et 90 mm. Le site Canigou est caractérisé par la plus grande période de sécheresse avec un déficit pouvant atteindre 90
mm et une sécheresse estivale durant en moyenne quatre mois (de juin à septembre). Espinouse
Figure 6 : (A) Exemple de diagramme ombrothermique du site Cassagnoles et (B) analyse en composantes principales des conditions climatiques de chaque site (RR = précipitations, TM = température moyenne, TN_polli
= température minimale pendant la pollinisation, TX_dev = température maximale pendant le développement de la graine et sécheresse = déficit cumulé de précipitations pendant l’été.
B A SITES N° Ayre 1 Canigou Cassagnoles 2 3 Espinouse 4 Fontmort 5 LaFage 6 Les Chauvets 7 Lespinasière 8 Lingas Pardailhan Rialsesse 9 10 11 Table 1 : Noms des sites et numéros correspondant sur les figures 6, 8 et 9.
13
est le second site le plus touché par la sécheresse avec 3 mois de sécheresse de juin à août et un déficit cumulé de 48,6 mm. Les sites Rialsesse et Pardailhan présentent eux deux mois de sécheresse, respectivement juin-juillet et juillet-août. Seuls les sites Ayre (n°1), Les Chauvets (n°7) et Lingas (n°9) ne présentent pas de sécheresse estivale malgré des précipitations en baisse en juillet (Fig. 6B, annexe 1). Les derniers sites du dispositif régional Fontmort (n°5) et La Fage (n°6) présentent en moyenne un seul mois de sécheresse en juillet (15 mm de déficit cumulé en moyenne). Pour finir la lecture de l’ACP montre que les sites Fontmort (n°5) et Lingas (n°9) sont les plus corrélés avec la variable altitude et sont ceux les moins corrélés aux variables climatiques (Fig. 6B et Annexe 1).
b) Dispositifs en gradients altitudinaux.
Les données de la station météorologique du mont Serein (1400 m, versant nord) indiquent que le mont Ventoux a reçu 1270 mm de précipitations annuelles en moyenne (de 2007 à 2013), dont l’essentiel s’est produit en automne. La température moyenne annuelle était de 8,3°C pendant la période et l’été est la saison la plus sèche avec l’observation régulière de sécheresses estivales.
Les données obtenues grâce aux hobos permettent de dire que la température moyenne diminue de 0,74°C quand l’altitude augmente de 100 mètres. En Vésubie, la sécheresse intervient généralement en août, et la température moyenne diminue de 0,63°C pour une élévation de 100 mètres. En Issole, la température moyenne décroît de 0,15°C pour 100 mètres d’élévation, et une période de sécheresse a été marquée en juillet. La figure 7A permet de constater cette corrélation négative entre la température et l’altitude sur les 3 sites, bien qu’une inversion des températures apparaisse entre 1101 mètres (niveau 1) et 1202 mètres (niveau 2) en Issole. Il n’y a par contre pas de corrélation entre l’humidité relative et l’altitude (Fig. 7B), le taux d’humidité
Figure 7 : Température moyenne en °C (A) et humidité relative en % (B) en fonction de l’altitude (1=niveau le plus bas, 5=niveau le plus haut)
14
reste constant sur les différents niveaux du site Ventoux mais varie légèrement, avec une amplitude de 10% (68 à 78%), indépendamment des niveaux d’altitude sur les sites Issole et Vésubie (Fig. 7 et Annexe 1)
2. Etude de la variabilité de la quantité et de la qualité de graines produites a) Dispositifs régionaux
Sur la période étudiée (2004-2009), le nombre de graines produites par cône montre une importante variabilité spatiale, les Chauvets (n°7) et Lingas (n°9) étant les sites qui se sont démarqués le plus du niveau global de production de graines par cône (Fig. 8A) avec une production de graines par cône minimale (212 et 229 graines). Une variabilité interannuelle de la production tous sites cumulés est significative en raison d’un fort accroissement du nombre moyen de graines par cône en 2007 (232 graines) et 2008 (297 graines).
Figure 9 : Variations du pourcentage de graines saines (noir), vides (rouge), parasitées (vert) et de petites graines (orange) en fonction de chaque site toutes années confondues (A) et en fonction de chaque année tous sites confondus (B). Prédictions obtenues à partir du modèle 1.
2004 2006 2007 2008 2009 Années
A B
1 2 3 4 5 6 7 8 9 10 11 Sites
Figure 8 : Prédictions (modèle 1) du nombre moyen (+ l’intervalle de confiance à 95%) de graines par cône en fonction de chaque site (A ; les noms des sites apparaissent dans le tableau 1) et en
fonction de chaque année tous sites confondus (B).
A B N o m br e d e g rai ne s pa r c ô ne N o m br e d e g rai ne s pa r cô ne 1 2 3 4 5 6 7 8 9 10 11 Sites 2004 2006 2007 2008 2009 Années
15
Les prédictions des effets montrent une importante variabilité inter-site pour chaque catégorie de graines (Fig. 9A, annexe 2). Par exemple, les sites Espinouse (n°4) et Fontmort (n°5) produisent en moyenne 10% de petites graines en plus que les autres sites, ce sont les Chauvets (n°7) et Lingas (n°9) qui produisent le plus de graines vides (45%), et les graines sont davantage parasitées (environ 15%) à Fontmort (n°5) et Lespinassière (n°8) qu’Ayre (n°1), Les Chauvets (n°7) et Rialsesse (n°11). Egalement, le site Fontmort (n°5) a produit 20% de graines saines en moins par rapport à la Fage (n°6) et Canigou (n°2). Pour chaque site le taux de graines vides reste le plus conséquent (entre 30 et 50%) et le taux de graines parasitées le plus faible (entre 0 et 20%)
Il existe aussi une variabilité inter-annuelle (figure 9B) visible sur le taux de graines saines avec une diminution de cette production de 20% en 2007 suivi d’une augmentation de 10% en 2009. Cette variation inter-annuelle est également significative pour le taux de petites graines produites avec une augmentation progressive à partir de 2007 (10%) jusqu’en 2009 (30%). Au cours du temps les graines vides sont toujours plus largement représentées que les autres catégories avec un pic en 2007 et l’intensité du parasitisme reste faible et stationnaire autour de 10%.
b) Dispositifs en gradients altitudinaux
Variabilité de la longueur et de la masse des cônes
La longueur des cônes ne varie pas significativement entre sites. Il existe une corrélation positive entre la longueur des cônes et l’altitude pour le site Issole mais cette variation n’est pas significative pour les sites Vésubie et Ventoux (Fig. 10). Une variabilité inter-annuelle est aussi significative mais seulement pour le site Ventoux avec une augmentation de la longueur des cônes de 120 mm à 160 mm entre 2007 et 2008 (Annexe 2). Il existe aussi une variabilité entre individus (p-value = 2,2e-16) 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5 Lo ng ue ur d es c ô ne s (m m )
Figure 10 : Prédictions de la longueur des cônes (en mm) en fonction des différents niveaux d’altitude en Issole (rouge), Vésubie (bleu) et Ventoux (noir). A partir du modèle 2
16
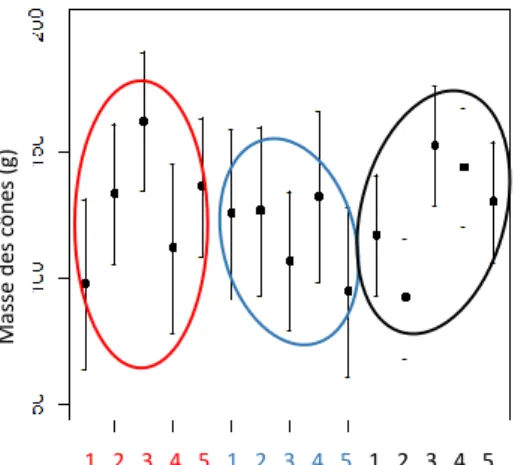
La masse des cônes varie significativement entre les individus (p-value=3,73e-06) mais elle ne diffère pas significativement entre les sites ni entre les différents niveaux d’altitude en Vésubie. Une tendance peut être observée en Issole car la masse des cônes augmente (de 98g à 162g) jusqu’au troisième niveau (à 1286 m) mais cette variation n’est pas significative. Au Ventoux, Pour finir la masse des cônes du site Ventoux présente variation significative entre le niveau 2 (1020 m) et le niveau 3 (1117 m) passant de 92g à 152 g (Fig. 11).
Variabilité du PMG
Le PMG représente la masse moyenne de mille graines (50,3 grammes en moyenne). L’étude des prédictions issues du modèle 2 montre qu’il n’y a pas de variations statistiquement significatives entre les différents niveaux d’altitude de chaque site. Egalement, il n’y a pas de variations statistiquement significatives entre années pour chaque site, ni de variabilité inter-sites.
Variabilité du taux de graines saines, vides et parasitées
Le taux de graines saines est toujours faible (<30%) mais ne varie pas significativement entre les sites ni entre les années (Fig. 12). Par ailleurs sur, le site Ventoux, le taux de graines saines varie significativement entre 1020 mètres (niveau 2) et 1117 mètres (niveau 3) passant de 10% à 25%. De la même façon le taux de graines vides varie significativement entre 1020 et 1117 mètre sur le mont Ventoux mais cette fois négativement passant de 90% à 70%. Le taux de graines vides varie aussi significativement entre le site Ventoux (75%) et les sites Issole (90%) et Vésubie (85%). Pour finir le taux de graines parasitées ne varie pas significativement entre les sites ni entre les différents niveaux d’altitude (Fig. 12).
Figure 11 : Prédictions de la masse des cônes en fonction des différents niveaux d’altitude en Issole (rouge), Vésubie (bleu) et Ventoux (noir). A partir du modèle 2. M as se d es c ô ne s (g ) 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
Figure 12 : Prédictions du taux de graines saines en fonction des différents niveaux d’altitude en Issole (rouge), Vésubie (bleu) et Ventoux (noir). A partir du modèle 2
G rai ne s sa ine s ( %) 1 2 3 4 5 1 2 3 4 5 1 2 3 4 5
17
3. Influence des facteurs climatiques sur les variables étudiées
Tous les sites d’étude du dispositif régional ne subissent pas les mêmes conditions pendant la période de pollinisation, notamment une forte augmentation des précipitations, de l’ordre de 100 mm, entre 2006 et 2009. Une diminution des températures maximales pendant la période de développement des graines est aussi observée. Ces variations suivent le même schéma pour tous les sites mais il y a une hiérarchisation des sites. Le site avec le pic de précipitation de plus faible ampleur correspondent à celui le plus au sud (Canigou), inversement les sites ayant subies la plus forte augmentation des précipitations pendant la pollinisation sont ceux situés le plus au nord dans les terres (Ayre et les Chauvets). Cette hiérarchisation est différente pour l’évolution de la température maximale lors du développement de la graine et l’évolution de la température minimale pendant la pollinisation. Par exemple le site Lingas présente les plus faibles températures (entre 15 et 20°C) et le site Espinouse subit les plus fortes températures (entre 25 et 30°C) pendant le développement des graines. Concernant les dispositifs altitudinaux, les trois sites suivent les mêmes évolutions climatiques pendant les périodes d’intérêts, ils ont tous les trois subit un pic de température pendant la pollinisation associé à une chute de l’humidité relative pendant la pollinisation en 2009. L’humidité relative pendant la période de développement de la graine reste relativement constante au cours du temps mais le pourcentage du site Vésubie reste supérieure (+4%). Inversement le site Vésubie est celui présentant les plus faibles températures pendant le développement de la graine au cours du temps (Annexe 3)
La comparaison de plusieurs modèles linéaires a permis de distinguer les facteurs ayant un impact sur chaque variable étudiée.
Dans le cas des dispositifs régionaux l’altitude à un effet significatifs sur trois des quatre variables (le nombre de graines par cône, le taux de petites graines et le taux de graines parasitées). La température maximale pendant le développement de la graine est le facteur qui influence le moins de variables, avec un effet négatif sur le nombre de graines produites par cône et un effet positif sur le taux de graines saines (Table 2). Le nombre de graines par cônes est influencé par trois facteurs, positivement par la température pendant la pollinisation et négativement pat l’altitude et la température pendant le développement de la graine. Le taux de petites graines est influencé positivement par tous les facteurs sauf la température du développement de la graine. Le taux de graines saines est influencé négativement par trois variables (les précipitations pendant la pollinisation et le développement de la graine et
18
l’altitude) tandis que la température de pollinisation n’a aucun effet et que la température pendant le développement de la graine a un effet positif. Le taux de graines vides est influencé par trois variables négativement alors que la température pendant le développement de la graine et l’altitude n’ont aucun effet. Pour finir le taux de graines parasitées est uniquement influencé positivement par trois facteurs (les précipitations pendant la pollinisation et le développement de la graine et les températures pendant la pollinisation).
Dans le cas des dispositifs en gradients altitudinaux la longueur des cônes est influencée (négativement) seulement par la température pendant la pollinisation (au seuil de 9%. Les variables les plus sensibles aux facteurs climatiques et à l’altitude sont les variables liées à la qualité des graines, c’est-à-dire que seul le taux d’humidité relative pendant la pollinisation n’a pas d’effet sur le taux de graines saines et le taux de graines vides, de même le seul facteur climatique qui n’influence pas le taux de graines parasitées est la température pendant la phase de développement des graines. Le taux de graines saines est influencé négativement par l’altitude et la température pendant le développement de la graine mais positivement par l’humidité relative de l’air pendant cette période. Le taux de graines vides et de graines parasitées sont tous deux influencés positivement par l’altitude et négativement par l’humidité relative du développement de la graine et la température pendant la pollinisation.
TN_polli HR_polli TX_dev HR_dev altitude
longueur cônes 0,09 0,19 0,26 0,65 0,11
masse cônes 0,61 0,04 0,94 0,16 0,08
PMG 0,71 0,52 0,98 0,99 0,18
graines saines 3,4 e-12 0,26 0,007 3,32 e-09 0,03
graines vides 3,53 e-13 0,61 0,01 2,2 e-16 0,03
graines parasitées 0,016 0,005 0,11 8,8 e-10 0,02
TN_polli RR_polli TX_dev RR_dev altitude Nbr graines/cône 4,4 e-04 0,55 1,08 e-06 0,41 2,6 e-14
petites graines 0,002 0,007 0,41 4,12 e-05 1,43 e-10
graines saines 0,6 9,53 e-13 3,7 e-10 0,01 1,7 e-04
graines vides 2,31 e-07 9,1 e-04 0,21 2,03 e-05 0,69
graines parasitées 0,06 0,13 0,38 0,03 0,014
Table 2 : Résumé des p-value obtenues à partir des modèles 3 et 4 et effets de facteurs environnementaux sur chaque variable. TN_polli et RR_polli = températures minimales et précipitations pendant la pollinisation, TX_dev et RR_dev = températures maximales et précipitations pendant la période de développement de la graines. En rouge les effets négatifs, en vert les effets
positifs et en gris les effets non significatifs.
Tableau 3 : Résumé des p-value obtenues à partir des modèles 3 et 4 et effets de facteurs environnementaux sur chaque variable. TN_polli et HR_polli = températures minimales et humidité relative pendant la pollinisation, TX_dev et HR_dev = températures maximales et humidité relative pendant la période de développement de la graines. En rouge les effets négatifs, en vert les effets
19 IV. Discussion
Nos résultats montrent une variation significative de la production de semences dans le temps et dans l’espace. La variabilité interannuelle est marquée par une augmentation de la production d’environ 60 graines par cônes, entre les années 2007 et 2008, et la variabilité inter-sites est marquée par une production de graines par cône plus faible d’environ 30 graines pour les sites Lingas et les Chauvets. De plus ces deux sites sont aussi ceux produisant le plus fort taux de graines vides. Au contraire, les variations entre gradients altitudinaux ou entre années ne sont pas systématiques. En revanche, il semble que l’altitude soit une source environnementale de variation de la production de graines, sur les plans qualitatif et quantitatif.
Variabilité interannuelle de la production de graines
La cyclicité de la production de graines est un facteur fréquemment étudié (Greene et Johnson 2004). Dans notre étude il existe une variation interannuelle de la production de graines par cônes, en 2007 une production de 240 graines par cônes en moyenne et en 2008 une production de 300 graines par cônes en moyenne. Sur 22 espèces nord-américaines, Greene et Johnson (2004) ne détectent qu’une tendance modeste pour qu’une année de bonne production de semences soit suivie d’une année de faible production. Ils expliquent cette variation interannuelle par un effet du climat, qui semble selon eux bien plus important que celui d’un épuisement de ressources. Greene et Johnson (2004) ont travaillé sur plusieurs espèces, des genres Pinus et Quercus, provenant du nord de l’Amérique situées au cœur de leur aire de distribution contrairement aux populations de sapin pectinés situés en marge de leur aire de répartition et pour lesquelles les conditions climatiques sont plus contraignantes. Il se trouve qu’une augmentation des précipitations au mois de mai a été observée ainsi qu’une chute des températures estivales (juin, juillet et août) l’année précédant celle de forte production de graines de sapin en 2008. Dans le cycle de reproduction d’Abies alba, pendant la première année de son cycle de reproduction, le mois de mai correspond à l’initiation des bourgeons axillaires (mai) et à la fin de la phase de différenciation florale et les mois de juin, juillet, août correspondent au début de la phase de développement des bourgeons mâles et femelles. Nous pouvons donc supposer que cette augmentation du nombre de graines soit associée à des conditions favorables à la production florale l’année précédente.
20 Sensibilité à l’altitude et aux facteurs climatiques
L’altitude a un effet négatif sur certaines variables, plus elle augmente plus le nombre de graines diminue ainsi que le taux de graines saines. Il se trouve que pour les deux dispositifs le taux de graines saines diminue avec l’altitude. Le second dispositif nous informe également que le taux de graines vides augmente avec l’altitude. Ces résultats corroborent d’autres travaux montrant que le taux de graines produites est corrélé négativement avec l’altitude. En effet, la production de graines de l’étude de l’épinette de Norvège déclinerait en moyenne de 100 graines par 100 mètres d’altitude (Mencuccini et al. 1995). Les fluctuations de la production de graines avec l’altitude pourraient ainsi refléter des variations spatiales de qualité des sols, de densités des peuplements et de conditions climatiques. Mencuccini et al. (1995) associent ces variations aux différences de fertilité du sol et aux caractères climatiques liés à l’altitude. Notre étude ne prend pas en compte la fertilité du sol mais nous avons montré, comme attendu, que les températures sont corrélées négativement à l’altitude le long des trois gradients. Cette interaction entre le nombre de graines par cône et le taux de graines saines et l’altitude peut s’expliquer par des températures plus froides en altitude. De plus il a été montré dans de précédentes études que le froid et la sécheresse peuvent provoquer des anomalies du développement embryonnaire (Owens & Blake 1986). Il est toutefois intéressant de noter que le taux de graines saines augmente de 15% entre 1020 et 1117 mètres au Mont Ventoux, suggérant que des facteurs autres que le climat, qui n’ont pas été pris en compte dans nos modèles, influencent la fructification du sapin sur ce site. On pourrait citer par exemple les conditions édaphiques, l’âge ou la densité des peuplements.
Il se trouve que dans notre dispositif régional, les deux sites les plus élévés, les Chauvets (1236 m) et Lingas (1275 m), se distinguent de par leur nombre de graines produites par cône le plus faible (215 graines) et par la plus forte production de graines vides (45%). Nos résultats montrent que l’altitude avait un effet négatif sur le nombre de graines par cônes mais elle n’a pas d’effet sur le taux de graines vides dans le cas de ces deux sites, il existe donc un autre paramètre expliquant le fait qu’ils se distinguent des autres. La température, l’humidité relative de l’air ou encore les précipitations sont des facteurs climatiques pouvant également influencer la fructification des conifères (Philippe et al. 2006; Roland et al. 2014). Les profils climatiques nous informent que ce sont les deux seuls sites qui ne subissent aucune période de sécheresse au cours de l’année. Malgré des précipitations annuelles de niveau moyen, l’absence de sécheresse estivale fait d’eux les sites les plus humides pendant l’été, c’est-à-dire pendant la période de développement de la graine (de juin à août).
21 Des facteurs biotiques peu influants/les insectes prédateurs des graines
Une des causes majeures de pertes de graines chez l’espèce Abies amabilis sont les dégâts occasionnées par les insectes (32 à 39%) pendant le développement de l’ovule puis de la graine (Owens et Morris 1998). En outre nos résultats montrent que l’altitude influence le taux de graines parasitées de façon positive. La larve qui parasite les graines d’A. alba est celle d’un hyménoptère ravageur du sapin du genre Megastigmus. En France on rencontre essentiellement deux espèces : M. suspectus qui est une espèce native présente partout en Europe et M. rafni qui est originaire d’Amérique du nord (Candau et al. 2009). Le niveau de parasitisme de nos peuplements reste globalement faible et stationnaire (entre 10 et 20 %) en région méditerranéenne par rapport à Owens et Morris (1998). Pour expliquer les variations dans le taux de parasitisme et cet effet de l’altitude observé un échantillonnage plus intensif des espèces de Megastigmus permettrait d’estimer si leur abondance est elle-même influencée par l’altitude. Cet effet de l’altitude pourrait être analysé plus en détail.
Interaction pollinisation et climat
Les conditions climatiques, en particulier les précipitations, peuvent aussi influer sur le succès de pollinisation, et donc le développement des graines saines, en perturbant la libération et l’acheminement du pollen vers les fleurs femelles et/ou en contrariant la capture du pollen et son transport (Philippe et al. 2006). Notre étude montre qu’une augmentation des précipitations pendant la période de pollinisation entraîne une augmentation du taux de petites graines et une diminution des taux de graines saines et vides. En effet, la pluie, en particulier les fines gouttelettes, peut chasser les grains de pollen vers le sol (lessivage) et réduire ainsi la quantité de pollen disponible sur les cônes femelles (Owens et Blake 1986). Ceci affecte donc directement le succès de la pollinisation et peut expliquer l’accroissement du taux d’avortement précoce des ovules chez le sapin. Il en résulte une chute du taux de graines saines disponibles. La baisse du taux de graines vides pourrait s’expliquer également par un lessivage accru du pollen, puisque ce type de graines résulte généralement d’auto-fécondation (Owens 1991; Philippe et al. 2006).
Des variables non expliquées
La qualité des graines est certes influencée par le climat, mais nos résultats indiquent que l’altitude ainsi que les facteurs climatiques pris en compte n’ont aucun effet sur la longueur ni sur la masse des cônes. Ces variables impliquent des processus physiologiques d’acquisition et d’exploitation des ressources par l’arbre qui ne sont pas abordées dans cette étude. Mais l’on
22
sait que le climat influence le succès du développement d’une graine, on peut donc supposer qu’une fois la maturation de la graine possible, il n’influe plus sur le stade final du développement. Sachant que le climat influence aussi le nombre de cônes produits (Mutke et
al. 2005) mais pas leur longueur ni leur masse on peut supposer que l’hypothèse précédente est
valable pour la structure reproductrice en elle-même : le cône.
Il est difficile de déterminer précisément quel facteur est responsable de la variation de de la production de graines et notamment les facteurs entrant en jeux lors des variations observées sur un gradient altitudinal. Nous avons montré que cette variation est due en partie aux variations de température et que la phase de pollinisation, qui joue un rôle important dans le taux de graines viables produit par un cône, est sensible aux changements climatiques. D’autres facteurs n’ont pas été étudiés dans cette étude, par exemple certaines études émettent l’hypothèse que ces variations peuvent être expliquées par une réduction de l’activité photosynthétique en haute altitude (Körner 2003). D’après Ayari et al. 2012, le nombre de cônes par arbre est déterminé par l’âge, la densité des peuplements et la taille de la couronne pour le pin d’Alep. Il serait donc intéressant, dans un travail futur, d’étudier si ces facteurs ont aussi un effet direct sur la production de graines. Enfin, cette étude dispose probablement d’une série temporelle de données encore trop courte pour pouvoir faire émerger de plus fortes tendances sur le plan statistique. Cette étude met en évidence que la phase de pollinisation est déterminante dans la qualité de la production de semences et c’est une période sensible aux variations climatiques. L’altitude semble aussi être un facteur limitant du nombre de graines par cône. Enfin, la température et les précipitations influencent davantage la qualité que la quantité des graines produites. Les effets des facteurs climatiques sur la production de semences étudiés ici mettent donc en lumière des facteurs environnementaux à considérer pour une meilleure compréhension et la prédiction de la dynamique spatio-temporelle du sapin pectiné en marge sud de son aire de répartition, dans un contexte de changement climatique.
23 V. Bibliographie
Allen, C.D., Macalady, A.K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., Kitzberger, T., Rigling, A., Breshears, D.D., Hogg, E.T. & others. (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. Forest ecology and management, 259, 660–684. Ayari, A., Zubizarreta-Gerendiain, A., Tome, M., Tome, J., Garchi, S. & Henchi, B. (2012) Stand, tree and crown variables affecting cone crop and seed yield of Aleppo pine forests in different bioclimatic regions of Tunisia. Forest Systems, 21, 128.
Boivin, T. & Auger-Rozenberg, M.A. (2016) Native Fruit, Cone and Seed Insects in the Mediterranean Basin, Springer, The Netherlands. T.D. Paine, & F. Lieutier (Eds.).
Cailleret, M. & Davi, H. (2011) Effects of climate on diameter growth of co-occurring Fagus sylvatica and Abies alba along an altitudinal gradient. Trees, 25, 265–276.
Calama, R. & Montera, G. (2007) Cone and seed production from stone pine (Pinus pinea L.) stands in Central Range (Spain). European Journal of Forest Research, 126, 23–35.
Candau, J.-N., Chalon, A. & others. (2009) L’impact des insectes parasites des graines sur les capacités de régénération naturelle des peuplements de cèdre de l’Atlas et de sapin pectiné en Languedoc-Roussillon. Clark, J.S. (1998) Why trees migrate so fast: confronting theory with dispersal biology and the paleorecord. The
American Naturalist, 152, 204–224.
Côme. (1982) Influence de la réfrigération et de la congélation sur la qualité et l’aptitude à la germination des graines. International Journal of Refrigeration, 5, 333–336.
Fenner, M. (1987) Seedlings. New Phytologist, 106, 35–47.
Greene, D.F. & Johnson, E.A. (2004) Modelling the temporal variation in the seed production of North American trees. Canadian Journal of Forest Research, 34, 65–75.
Kang, K.S., Bila, A.D., Harju, A.M. & Lindgren, D. (2003) Estimation of fertility variation in forest tree populations. Forestry, 76, 329–344.
Koenig, W.D. & Knops, J.M. (2000) Patterns of annual seed production by northern hemisphere trees: a global perspective. The American Naturalist, 155, 59–69.
Körner, C. (2003) Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems ; with 47 Tables, Springer Science & Buisness Media.
Matthews, J.D. (1963) Factors affecting the production of seed by forest trees. , 24, 1–13. Matyas, C. & Giertych, M. (2013) Genetics of Scots Pine, Elsevier.
McNeill, W.M. (1954) Observations on cone and seed production in plantations of Scots pine in Scotland.
Forestry, 27, 122–133.
Mencuccini, M., Piussi, P. & Sulli, A.Z. (1995) Thirty years of seed production in a subalpine Norway spruce forest: patterns of temporal and spatial variation. Forest Ecology and Management, 76, 109–125.
Morin, X., Viner, D. & Chuine, I. (2008) Tree species range shifts at a continental scale: new predictive insights from a process-based model. Journal of Ecology, 96, 784–794.
Mutke, S., Gordo, J. & Gil, L. (2005) Variability of Mediterranean Stone pine cone production: Yield loss as response to climate change. Agricultural and Forest Meteorology, 132, 263–272.
Owens, J.N. (1991) Flowering and seed set. Physiology of trees. Wiley, New York, 247–271. Owens, J.N. & Blake, M.D. (1986) Production de Semences Forestières.
24 Owens, J.N. & Morris, S.J. (1998) Factors affecting seed and cone development in Pacific silver fir (Abies amabilis). Canadian journal of forest research, 28, 1146–1163.
Ozenda, P. (1981) Carte de La Végétation de La France Au 200 000e. Végétation Des Alpes Sud-Occidentales:
Notice Détaillée Des Feuilles 60 Gap-61 Larche-67 Digne-68 Nice-75 Antibes., Editions du Centre national de la
recherche scientifique.
Parmesan, C. & Yohe, G. (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature, 421, 37–42.
Philippe, G., Baldet, P., Héois, B. & Ginisty, C. (2006) Reproduction Sexuée Des Conifères et Production de
Semences En Vergers À Graines., Cemagref. Juliette Baudel.
Restoux, G. (2009) Variabilité Spatio-Temporelle de La Reproduction Chez Une Espèce Pérenne Monoïque: Le
Cas Du Sapin Pectiné, Abies Alba Miller, En Limite Sud de Son Aire de Répartition. Paul Cezanne - Aix
Marseille III, Ecologie des forêts méditerranéennes - UR629 INRA Avignon.
Roland, C.A., Schmidt, J.H. & Johnstone, J.F. (2014) Climate sensitivity of reproduction in a mast-seeding boreal conifer across its distributional range from lowland to treeline forests. Oecologia, 174, 665–677.
Root, T.L., Price, J.T., Hall, K.R., Schneider, S.H., Rosenzweig, C. & Pounds, A.J. (2003) Fingerprints of global warming on wild animals and plants. Nature, 421, 57–60.
Sagnard, F., Barberot, C. & Fady, B. (2002) Structure of genetic diversity in Abies alba Mill. from southwestern Alps: multivariate analysis of adaptive and non-adaptive traits for conservation in France. Forest Ecology and
Management, 157, 175–186.
Tomlinson, P.B. & Takaso, T. (2002) Seed cone structure in conifers in relation to development and pollination: a biological approach. Canadian Journal of Botany, 80, 1250–1273.
Turgeon, J.J., Roques, A. & Groot, P.D. (1994) Insect fauna of coniferous seed cones: diversity, host plant interactions, and management. Annual review of entomology, 39, 179–212.
Wolf, H. (2003) EUFORGEN Technical Guidelines for Genetic Conservation and Use for for Silver Fir (Abies
Alba). Bioversity International.
Logiciel statistique :
R Development Core Team (2008). R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3-900051-07-0, URL http://www.R-project.org.
Site internet :
25 Table des annexes
ANNEXE 1 : Diagrammes ombrothermiques.
ANNEXE 2 : Etude de la variabilité de la production de graines à partir des prédictions des modèles 1 et 2.
ANNEXE 3 : Evolution des températures, des précipitations et de l’humidité relative pendant la phase de pollinisation et pendant le développement de la graine.
26 ANNEXE 1 : Diagrammes ombrothermiques
27
28 2004 2006 2007 2008 2009 1 2 3 4 5 6 7 8 9 10 11 Sites 2004 2006 2007 2008 2009 1 2 3 4 5 6 7 8 9 10 11 Sites 2004 2006 2007 2008 2009 1 2 3 4 5 6 7 8 9 10 11 Sites
ANNEXE 2 : Etude de la variabilité de la production de graines à partir des prédictions des modèles 1 et 2
Nombre de graines par cône
Variable petites graines
29 1 2 3 4 5 6 7 8 9 10 11 Sites 2004 2006 2007 2008 2009 2006 2007 2008 2011 2012 2013 2007 2008 2011 2007 2008 2011 2004 2006 2007 2008 2009 1 2 3 4 5 6 7 8 9 10 11 Sites
Variable graines vides
Variable graines parasitées
30 ANNEXE 3 : Evolution des températures, des précipitations et de l’humidité relative
pendant la phase de pollinisation et pendant le développement de la graine.
SITES Ayre Canigou Cassagnoles Espinouse Fontmort La Fage Les Chauvets Lespinasière Lingas Pardailhan Rialsesse SITES Ayre Les Chauvets Canigou Cassagnoles Lespinasières Espinouse Pardailhan Fontmort Lingas La Fage Rialsesse
31
Ventoux
Issole
32 Résumé
Les écosystèmes forestiers sont aujourd’hui touchés par les conséquences des changements globaux en cours. Les conditions climatiques prévues au cours des prochaines décennies devraient induire des modifications dans la plasticité et l’évolution des espèces et conduire à des changements dans leur distribution géographique. Les populations d’arbres établis en marge de leur aire de distribution sont particulièrement sensibles aux modifications de leur environnement et sont déjà contraintes ces changements en cours. Leur succès reproducteur est une composante majeure de leurs dynamiques spatio-temporelles dans ce contexte d’environnement changeant. Nous cherchons ici à identifier les facteurs environnementaux impliqués dans la variabilité de la production quantitative et qualitative de semences du sapin pectiné (Abies alba), en limite sud de son aire de distribution (pourtour méditerranéen français). L’analyse de données historiques de production de semences confrontées à des facteurs climatiques, topographiques et biotiques suggère que la période de pollinisation est une phase sensible aux variations climatiques et elle détermine en partie la qualité de la production de semences. L’altitude semble être un facteur limitant du nombre de graines par cône. La température et les précipitations influencent davantage la qualité que la quantité des graines produites. Cette étude met en évidence les facteurs à prendre en considération pour une meilleure compréhension de la dynamique spatiale et temporelle des arbres en marge de leur aire de distribution
Mots-clés : production de graines, qualité, changement climatique, altitude, pollinisation.
Today, forest ecosystems receives the consequences of global climate change. Climatic conditions expected in the coming decades should lead to changes in species plasticity and evolution, and lead to changes in geographical distribution. Trees populations, which are fixed on the fringe of their distribution area, are particulary sensitive to changes in their environment and they are already forced by this modifications. Their reproductive success is an essential part of their spatial and temporal dynamics in this context. In this study, we search to indentify what environmental factors are involved in seed production variability quantitative and qualitative of silver fir (Abies alba) on the south fringe of this distribution area (french mediterranean region). Analysing of seed production historic data facing climatic, topographic and biotic factors suggest pollination period is a delicate phase with climate modifications and it decides, in part, the seed production quality. The elevation seems to be a limitant factor for the seed production per cone. Temperature and precipitation affect more the quality than the quantity of seed production. This study bring to light factors must be taken to a better understanding of the spatial and temporal dynamic of trees on the fringe of ditribution area.