UNIVERSITE DE SHERBROOKE
Faculte des sciences appliqueesDepartement de genie civil
ETUDE DE LA DEGRADATION DES ACIDES RESINIQUES PAR LE CHAMPIGNON
PHANEROCHAETE CHRYSOSPORIUM
IVlemoire de maitrise es sciences appliquees Specialite: genie civil (environnement)
Pierre Jonathan HODGSON
RESUME
Le champignon de la poumture blanche du bois Phanerochaete chrysosporium (BKM-F-1767) a ete etudie dans Ie but d'evaluer sa capacite a degrader les acides resiniques (AR), en
particulier les acides dehydroabietique (ADA) et abietique (AA). Ces substances d'origine
naturelle sont contenues majoritairement dans 1'oleoresine des coniferes et sont liberees lors des operations de mise en pate. Les AR sont responsables en grande partie de la toxicite des effluents papetiers et bien qu'ils puissent etre enleves efficacement par les systemes de traitement aerobies conventionnels, ils peuvent cependant etre la cause de problemes d'operation des reacteurs anaerobies traitant certains effluents des industries de pates et papiers. La premiere partie de cette etude avait pour but de verifier la compatibilite de 1'AA et de 1'ADA avec P. chrysosporium. Differents parametres physiologiques du champignon tels que sa croissance, 1'activite en solution des enzymes ligninolytiques, la production de proteines extracellulaires et la consommation de glycerol et d'ammonium ont ete suivis pour des concentrations variables en AR. Les resultats montrent que des concentrations en AA et en ADA jusqu'a 800 mg/L (concentration excedant par un facteur de 10 a 20 les concentrations typiques mesurees dans un effluent reel) ont modifie 1'evolution des parametres cites ci-haut sans toutefois menacer la viabilite du champignon. La seconde partie de cette etude a pennis, a 1'aide d'analyses effectuees par chromatographie en phase gazeuse avec detection par spectrometrie de masse, de mettre en
evidence un enlevement de 48 % de 1'ADA (concentration initiale de 100 mg/L) apres 10 jours
d'incubation dans un milieu de culture a base de glycerol sous atmosphere initial constitue d'oxygene pur. L'incubation abiotique de 1'AA dans ces conditions physico-chimiques a ete la cause de sa disparition complete, rendant impossible toute conclusion sur 1'enlevement de cet AR par P. chrysosporium. Finalement, 1'effet de 1'enlevement fongique de 1'ADA a ete verifie en effectuant des tests de toxicite anaerobie (TTA) sur Ie contenu final des cultures. II en est ressorti que Ie traitement de 1'ADA (200 mg/L) par P. chrysosporium a permis une reduction de 1'inhibition de la methanogenese causee par cet AR et a ainsi permis a la biomasse anaerobie d'effectuer un enlevement accru de la demande chimique en oxygene.
REMERCIEMENTS
Je tiens a souligner en premier lieu Ie support inconditidnnel que mes parents m'ont apporte au cours de ma maitrise et de toutes les annees d'etude precedentes. Je tiens a remercier tres sincerement MM. Roland Leduc de 1'Universite de Sherbrooke, Serge Guiot de Institut de
recherches en biotechnologie (IRB/CNRC) et Marcel Asther du Laboratoire de Biotechnologie
des Champignons Filamenteux (LBCF/INRA) qui ont supervise et mis sur pied ce projet de
recherche. Une place toute speciale doit etre reservee a Mme Chantal Laugero pour ses precieux conseils et son travail de supervision directe lors des travaux en laboratoire.
Je remercie egalement 1'equipe de chimie analytique de FInstitut de recherches en
biotechnologie (IRB/CNRC) pour 1'aide technique et scientiflque apportee lors de ce projet: Jalal
al Hawari, Chantal Beaulieu, Stephane Deschamps et Alain Comveau. La contribution d'Alain Patoine pour la mise au point. Ie rodage et 1'enseignement de la methode de dosage des acides resiniques doit etre soulignee avec insistance. Je desire egalement remercier M. Denis Rho (IRB/CNRC) pour son soutien au cours de la redaction de ce memoire.
Je tiens a remercier d'une fa9on toute personelle les membres du groupe de Bioingenierie
de 1'enviroimement de FIRB/CNRC (Sylvie Rocheleau, Jean-Claude Frigon, Rachid el
Mamouni), du LBCF/INRA (Michel Delattre, Pierre Frasse, Serge Moukha, Mireille Haon, Jean-Claude Sigoillot, Eric Record) et du Laboratoire de genie de 1'environnement de 1'Universite de Sherbrooke (Serge Bembe) pour leurs precieux conseils mais surtout pour les agreables moments passes en leur compagnie.
Ce projet de recherche a ete finance en partie par Ie Ministere quebecois des affaires intemationales, 1'Association pour la recherche intemationale en biotechnologie (ARIBIO), Ie CRSNG par 1'entremise de la subvention de recherche OGP 0120775 ainsi que Ie Programme de cooperation franco-quebecois scientifique et technologique.
LISTE DES ABREVIATIONS:
AA: acide abietique
ADA: acide dehydroabietique
AGV: acide gras volatil
AR: acide resinique
ARM: ester methylique d'acide arachidique
AVE: alcool veratrylique
BPC: biphenyl polychlore
BSTFA: N-0-bis-trimethylsyliltrifluoroacetamide (agent de derivation)
CLHP: chromatographie en phase liquide a haute performance CPG: chromatographie en phase gazeuse
CPG/SM: chromatographie en phase gazeuse a detection par spectrometrie de masse CTMP: mise en pate par voie chimico-thermomecamque
DCO: demande chimique en oxygene Cm: coefficient d'extinction molaire
HAP: hydrocarbure polycyclique aromatique
I^m; inhibition de 1'activite methanogenique
IpTnr inhibition de la production totale de methane
kDa: kiloDalton
LiP: peroxydase de lignine
MeOH: methanol
MnP: peroxydase de manganese MPA: acide o-methyl podocarpique MTBE: methyl-ter-butyl ether
MVS: matieres volatiles en suspension
OHPA :bacteries acetog6niques productrices d'H^.
PCP: pentachlorophenol
r/min: rotation par minute
TAH: test d'activite hydrogenotrophique
TABLE DES MATIERES
l.INTRODUCTION... 1 1.1 PROBLEMATIQUE... 1 1.2 COUPLAGEFONGIQUEETBACTERIEN... 2 1.3 OBJECTIFS ... 2 1.4 PLAN DUMEMOIRE ... 2 2. REVUE DE LITTERATURE ... 5 2.1 LETRAITEMENTANAEROBIE... 6 2.1.1 Principesetmecanismes... 6 2.1.2 Avantagesetdesavantagesdutraitementanaerobie... 6 2.2 LESACIDESRESINIQUES... 92.2.1 Description, provenance et structure ... 9
2.2.2 Toxicite des acides resiniques ... 11
2.2.3 Traitement aerobie d'effluents contenant des acides resiniques... 12
2.2.4 Traitement anaerobie d'effluents contenant des acides resiniques... 13
2.2.5 Modification et pretraitement physico-chimique des acides resiniques... 15
2.2.6 Degradation biologique des acides resiniques par des cultures pures ... 15
2.3 PHANEROCHAETECHRYSOSPOmUM... 18
2.3.1 Origine et description... 18
2.3.2 Description de la structure de la lignine... 19
2.3.3 Mecanismes d'action des lignine et manganese peroxydases ... 20
2.3.4 Role de 1'acide oxalique ... 23
2.3.5 Production du peroxyde d'hydrogene ... 24
2.3.6 Conditions de production des enzymes ligninolytiques... 24
2.4 APPLICATIONS POTENTIELLES DE P. CHRYSOSPORIUM... 26
2.4.1 Degradation de composes recalcitrants par P. chrysosporium... 26
2.4.2 Utilisations de P. chrysosporium par les industries papetieres... 28
3. MATERIEL ET METHODES... 32 3.1 SOUCHESMICROBIENNES... 32 3.1.1 Phanerochaete chrysosporium... 32 3.1.2 Bacteries anaerobies ... 32 3.2 PRODUITSCHIMIQUES SPECIALISES...32 3.3ESSAISFONGIQUES ... 32
3.3.1 Milieu de culture fongique ... 32
3.3.2 Protocole general de demarrage de cultures fongiques... 33
3.3.3 Essai de toxicite des acides resiniques surP. chrysosporium... 34
3.3.4 Essai de degradation des acides resiniques par P. chrysosporium... 34
3.3.5 Essai de degradation des acides resiniques avec suivi de latoxicite anaerobie... 35
3.4 METHODES ANALYTIQUES POUR LESUIVIDES CULTURES FONGIQUES... 35
3.4.1 Estimation de la biomassefongique... 35
3.4.2 Dosage des activites enzymatiques ligninolytiques... 35
3.4.3 Dosage des proteines extracellulaires... 36
3.4.4 Dosage de Fammonium... 36
3.4.5 Dosage duglycerol...>... 36
3.4.6 Preparation des echantillons pour Ie dosage des acides resiniques ... 37
3.4.7 Dosage des acides resiniques... 38
3.5 EVALUATION DU TRAITEMENT FONGIQUE PAR LE TEST DE TOXICITE ANAEROBIE... 39
3.5.1 Description du test de toxiciteanaerobie... 39
3.5.2 Protocole de preparation des echantillons soumis au TTA... 40
3.5.3 Protocole de demarrage du test de toxicite anaerobie... 41
3.5.4 Reactifs et solutions du TTA... 42
3.5.5 Suivi du test de toxicite anaerobie... 45
1^ TEST D'ACTIVITEHYDROGENOTROPHIQUE... 46
3.6.1 Description du test d'activitehydrogenothrophique... 46
3.6.2 Procedure de demarrage du TAH ... 46
3.7.1 Estimation de la biomasseanaerobie... 47
3.7.2 Dosage de la demandechimique en oxygene... 47
3.7.3 Dosage des acides gras volatils... 48
3.7.4 Mesure de la production de biogaz... 48
3.7.5 Analyse de la composition dubiogaz... 48
4. RESULTATS ET DISCUSSION... 50
4.1 ESSAI DE TOXICITE DES ACIDES RESINIQUES SURP. CHRYSOSPORIUM... 50
4.1.1 Effet sur la croissance fongique etl'excretion de proteines... 50
4.1.2 Effet des acides resiniques sur la consommation de nutriments... 51
4.1.3 Effet des acides resiniques sur la production d'enzymes ligninolytiques ... 51
4.1.4 Discussion du test de toxicite fongique des acides resiniques... 54
4.2 EXPERIENCE DE DEGRADATION DES ACIDES RESINIQUES PAR P. CHRYSOSPORIUM. ... 57
4.2.1 Disparition des acides resiniques... 57
4.2.2 Suivi des parametres physiologiques de 1'experience de degradation de 1'ADA... 59
4.2.3 Discussion du test de degradation desacidesresiniques... 59
4.3 TESTS DE TOXICITEANAEROBIE... 64
4.3.1 Essai preliminaire de sensibilite de laboue anaerobie... 65
4.3.2 Etude de la stabilite abiotique de 1'inhibition de 1'activite methanogenique... 68
4.3.3 Suivi du traitement fongique de 1'ADA a 1'aide du test de toxicite anaerobie... 69
4.3.4 Discussion des tests de toxicite anaerobie... 76
4.4 DISCUSSION GENERALEDESRESULTATS... 80
5. CONCLUSION... 82
Annexe 1 Protocole de dosage de laperoxydase de lignine... 84
Annexe 2 Protocole de dosage de la peroxydase de manganese ... 87
Annexe 3 Explication du calcul pour Ie dosage des AR... 89
Annexe 4 Donnees du test de toxicite fongique des AR ... 94
Annexe 5 Donnees de 1'essai de degradation fongique des AR... 97
Annexe 7 Donnees de 1'essai de stabilite abiotique de 1'inhibition methanogenique des AR.... 105
Annexe 8 Donnees du TTA effectue sur les cultures fongiques (0 et 200 mg/L d'ADA) ... 108
Annexe 9 Donnees du test d'activite hydrogenothrophique ... 113
Annexe 10 Resultats du premier TTA effectue sur des cultures fongiques traitant 1'ADA... 114
Annexe 11 Experience de degradation in vitro de 1'ADA... 116
Annexe 12 Experience sur 1'effet de 1'acetone envers P. chrysosporium... 118
LISTE DES FIGURES
Figure 1.1 Schema de la structure du memoire et de la demarche experimentale ... 4
Figure 2.1 Schema simplifie de la digestion anaerobie... 7
Figure 2.2 Principaux acides resiniques retrouves au sein des effluents papetiers... 10
Figure 2.3 Alcools phenoliques precurseurs de la lignine... 20
Figure 2.4 Cycle catalytique de la LiP ... 22
Figure 2.5 Cycle catalytique de la MnP... 23
Figure 2.6 Mecanisme propose de production du radical hydroxyle par la LiP... 24
Figure 3.1 Schema du protocole de preparation des echantillons soumis au TTA... 43
Figure 3.2 Schema de la burette utilisee pour la mesure de la production de biogaz... 49
Figure 4.1 Croissance de P. chrysosporium expose a differentes concentrations d'AAetd'ADA... 52
Figure 4.2 Concentration specifique en proteines extracellulaires au sein de cultures de P. chrysosporium exposees a differentes concentrations d'AA et d'ADA... 52
Figure 4.3 Evolution de la concentration residuelle en glycerol au sein de cultures de P. chrysosporium exposees a differentes concentrations d'AA et d'ADA... 53
Figure 4.4 Evolution de la concentration residuelle en ammonium dans les cultures de P. chrysosporium exposees aux AR... 53
Figure 4.5 Activite LiP specifique obtenue en presence de differentes concentrations d'AA et d'ADA... 55
Figure 4.6 Activite MnP specifique obtenue en presence de differentes concentrations d'AA et d'ADA... 55
Figure 4.7 Disparition des AR au sein des cultures de P. chrysosporium contenant initialement 100 mg/Ld'AR... 58
Figure 4.8 Suivi des parametres physiologiques des cultures de P.chrysosporium utilisees pour Ie traitement de 1'ADA (100 mg/L).. ... 60
Figure 4.9 Suivi de 1'activite enzymatique et du taux d'enlevement de 1'ADA observe dans des cultures de P.chrysosporium contenant 100 mg/L d'ADA... 60
Figure 4.10 Courbes de production cumulative de methane du test de sensibilite de la boue anaerobie effectue avec de 1'AA et de 1'ADA a une concentration de 60 mg/L sous
differentes concentrations en MVS... 66 Figure 4.11 Productions normalisees de methane obtenues lors du TTA effectue sur les cultures
fongiques (0 et 200 mg/L d'ADA)... 74 Figure 4.12 Consommation d'hydrogene gazeux lors du test d'activite hydrogenothrophique
LISTE DES TABLEAUX
TABLEAU 3.1 Composition du milieu de culture fongique 33
TABLEAU 3.2 Methode de dosage de I'ammonium €NN4 )... 36TABLEAU 3.3 Methode de dosage du glycerol... 37 TABLEAU 3.4 Methode de dosage des AR....„,..,.,„...„,..„,.„.,..„„,.,..„..„.„„„„„.,„„„.„„. 39
TABLEAU 3.5 Distribution des solutions necessaires au TTA 42
TABLEAU 3.6 preparation du milieu defini pour Ie TrTA^^^^^^^ ^^^^ ^^^^^ ^^^^ 44
TABLEAU 3.7 Solutions meres du milieu defmi 44
TABLEAU 3.8 Methode de dosage des acides gras volatils, 48
TABLEAU 3.9 Methode d'analyse du biogaz... 49 TABLEAU 4.1 Variation de 1'inhibition methanogenique en fonction de la charge
specifique en AR... 68
TABLEAU 4.2 Effet de 1'incubation abiotique des AR sur 1'inhibition de 1'activite
methanogenique ^.,. ,.... .,...^.^..^...^...„„...„....„..„...„.„„„..69
TABLEAU 4.3 Identification et description des echantillons du TTA... 73
1. INTRODUCTION
1.1 ProblematiqueL'activite industrielle et agricole s'est souvent faite au detriment de la qualite de 1'environnement. Les composes chimiques synthetiques produits par 1'homme sont souvent pointes du doigt puisque plusieurs d'entre eux, difficilement biodegradables, toxiques, mutagenes eVou cancerigenes, peuvent etre accumules a tous les niveaux de la chaine alimentaire. Certains composes d'origine naturelle sont egalement responsables de problemes environnementaux lorsqu'ils sont deverses dans des milieux recepteurs. Des composes sont deverses intentionnellement, comme 1'insecticide DDT (dichloro-diphenyl-trichloroethane) ou 1'herbicide atrazine, d'autres proviennent de divers rejets industriels deverses dans les cours d'eau, tandis que d'autres sont problematiques a cause de leur rejet dans Ie sol (pentachlorophenol, biphenyls polychlrores, solvants chlores).
Une batterie de precedes physico-chimiques est disponible et peut etre utilisee afin de traiter ces polluants. Malheureusement Ie cout souvent prohibitif de ces precedes peut representer 1'obstacle majeur de leur application. Cependant, la biodegradation des composes polluants par des micro-organismes constitue une solution qui peut souvent s'averer plus avantageuse. Citons a titre d'exemple les industries papetieres, qui pour la plupart en Amerique du Nord, traitent leurs effluents liquides par procede biologique aerobie a boues activees. Ce procede, tres repandu, porte toutefois Ie fardeau d'une importante production de boue dont les couts de disposition eleves peuvent nuire a la competitivite des compagnies sur les marches mondiaux. A cela, il faut egalement ajouter les inevitables depenses energetiques necessaires au maintien de niveaux suffisants d'oxy gene dissous au sein des bassins de traitement. Le traitement de ces effluents par precede anaerobie, produisant de faibles quantites de biomasse et ne necessitant aucune aeration peut toutefois en reduire les couts de depollution. Malheureusement, certains composes des effluents des usines de pates et papiers peuvent avoir un effet adverse sur les bacteries du traitement anaerobie. Tel est Ie cas des acides resiniques (AR), substances naturelles des coni&res qui sont relachees lors des operations de mise en pate et qui, s'ils ne sont pas traites, sont egalement une menace pour les systemes aquatiques recepteurs.
1.2 Couplage fongique et bacterien
La presente etude propose 1'etude de la degradation des AR par un champignon de la poumture blanche du bois Phanerochaete chrysosporium. Depuis les quinze demieres annees, un interet croissant a ete porte a ce champignon connu pour sa capacite naturelle a degrader la lignine. Isole au debut des annees 70, ce champignon, par la polyvalence et la puissance de ses capacites oxydatrices, s'est egalement revele capable de degrader une vaste gamme de composes recalcitrants. La presente etude traite de differents aspects de 1'enlevement des AR par P. chrysosporium dans 1'optique d'un nouveau type de couplage impliquant en premier lieu Ie traitement fongique des AR suivi d'un traitement anaerobie de 1'effluent detoxifie.
1.3 Objectifs
Le premier objectifde ce projet de recherche etait de determiner la compatibilite entre Ie champignon et deux representants de AR: les acides abietique et dehydroabietique. Dans cette optique. Ie champignon a ete mis en presence de concentrations variables des AR et Ie suivi de certains parametres physiologiques pouvant etre affectes par la presence des AR a ete effectue.
Le second objectif etait de verifier la capacite de P. chrysosporium a degrader les deux AR choisis au sein d'un milieu de culture liquide et d'identifler les parametres regulateurs de cette degradation. Le suivi de la teneur en AR a ete effectue par chromatographie en phase gazeuse et par spectrometrie de masse.
Le troisieme et demier objectifetait de determiner, a 1'aide du test de toxicite anaerobie, si Ie pretraitement fongique des AR pouvait permettre de reduire 1'effet adverse que peuvent avoir ces substances sur les bacteries du traitement anaerobie.
1.4 Plan du memoire
Le present memoire comporte deux grands volets. Le premier volet, presente au Chapitre 2, tente premierement de dresser un portrait global de la problematique engendree par les AR. La stmcture et la provenance des AR y sont traites. Les problemes enviroimementaux que peuvent causer les AR, en premier lieu au niveau ecologique et en deuxieme lieu au niveau des systemes de traitement anaerobies sont expliques. Differents moyens de prise en charge des AR sont
ensuite discutes. La derniere partie du premier volet porte sur Ie champignon Phanerochaete chrysosporium.
Le deuxieme volet de ce memoire est de nature experimentale. L'equipement et les protocoles experimentaux utilises au cours du present travail de recherche sont decrits en detail au Chapitre 3. La premiere serie de resultats experimentaux du Chapitre 4 porte sur la compatibilite physiologique entre les AR et P. chrysosporium, dans Ie but de jeter un premier regard sur la faisabilite du traitement fongique des AR. La deuxieme serie de resultats traite de la disparition des AR et des facteurs pouvant possiblement la declencher ou la reguler. La demiere serie de resultats a pour but, a 1'aide du test de toxicite anaerobie, 1'evaluation de 1'efficacite du traitement fongique des AR par Ie suivi de la toxicite anaerobie exercee par ces produits. Les resultats experimentaux sont discutes separement au fur et a mesure de leur devoilement, et sont mis en perspective d'une fa9on plus globale au cours de la discussion generale a la fin du Chapitre 4. La conclusion (Chapitre 5) constitue un rappel general des resultats et suggere egalement differents aspects qui meriteraient d'etre couverts lors de recherches ulterieures afin d'eclairer certains points n'ayant pu etre traites au cours de la presente recherche.
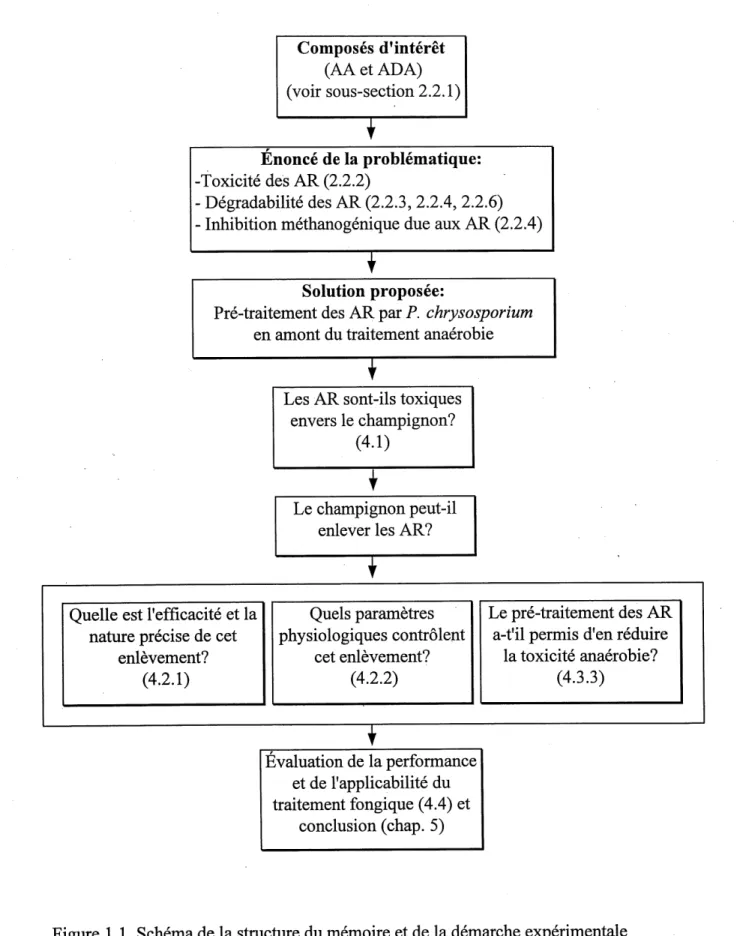
La Figure 1.1 schematise la structure globale du memoire et explique la demarche experimentale. Cette meme figure est egalement un guide de navigation au sein du memoire puisque Ie renvoi thematique a certaines sections et sous-sections y est effectue.
Composes d'interet
(AAetADA)
(voir sous-section 2.2.1)T
Enonce de la problematique: -Toxicite des AR (2.2.2) . Degradabilite des AR (2.2.3, 2.2.4, 2.2.6)Inhibition methanogenique due aux AR (2.2.4)
^
Solution proposee:
Pre-traitement des AR par P. chrysosporium en amont du traitement anaerobie
T
Les AR sont-ils toxiques envers Ie champignon?
(4.1)
T
Le champignon peut-il enlever les AR?
T
Quelle est I'efficacite et la
nature precise de cet enlevement?
(4.2.1)
Quels parametres physiologiques controlent cet enlevement?(4.2.2)
Le pre-traitement des AR a-t'il permis d'en reduirela toxicite anaerobie?
(4.3.3)
T
Evaluation de la performanceet de 1'applicabilite du
traitement fongique (4.4) et
conclusion (chap. 5)Figure 1.1 Schema de la stmcture du memoire et de la demarche experimentale
2. REVUE DE LITTERATURE
Tel que mentionne en introduction, la prise en charge de composes polluants par des micro-organismes peut dans beaucoup de cas s'averer une solution que les imperatifs economiques favoriseront, par opposition aux precedes de traitement physico-chimiques. Les champignons, les bacteries ou les algues peuvent etre utilises a ces fins et ont en commun trois differents modes de transformation possibles qui sont lies directement au metabolisme cellulaire:
• Le cometabolisme: Ce mode de transformation implique une modification structurale du compose polluant qui ne sert cependant pas de source d'energie. La presence d'un co-substrat permettant la croissance du micro-organisme est done necessaire.
• La biodegradation: Ce mode se distingue du cometabolisme par Ie fait que Ie compose polluant peut etre utilise comme source de carbone et d'energie servant les fins de croissance et de maintien du micro-organisme.
• La synthese: Les molecules du compose polluant et ses produits de transformation peuvent etre lies entre eux (polymerisation) ou a d'autres composes (conjugaison) par des mecanismes
enzymatiques.
La transformation microbienne peut permettre une reduction de la toxicite d'un polluant, effet qui doit etre verifie par des essais ecotoxicologiques, tandis que la mineralisation d'un polluant, impliquant sa conversion en dioxyde de carbone et en produits inorganiques, donne lieu a la detoxification du polluant. La mineralisation d'un polluant peut etre verifiee et quantifiee a 1'aide de molecules cibles isotopiquement marquees (carbone 14 a titre d'exemple), dont la mineralisation est source de dioxyde de carbone radioactif, lui-meme capture par piegage (NaOH par exemple) et quantifie a 1'aide d'un compteur a scintillation.
Ces prmcipes de base du traitement biologique sont applicables autant pour les micro-organismes associes au traitement aerobie, que pour ceux prenant part a la digestion anaerobie, sur laquelle la prochaine section porte.
2.1 Le traitement anaerobie
2.1.1 Prmcipes et mecamsmes
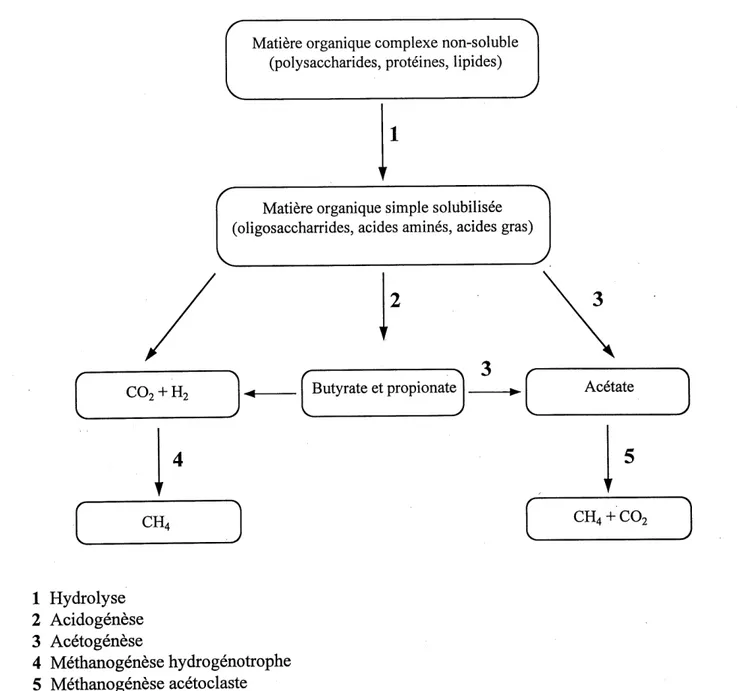
La biotransformation de la matiere organique en methane est un procede auquel prennent part differents groupes trophiques de bacteries anaerobies (facultatives ou obligees) PMOVAES, 1986]. Au cours de ce precede. Ie produit d'un groupe de bacteries devient Ie substrat d'un autre groupe pour donner ultimement lieu a la production de methane. La Figure 2.1 illustre de maniere simplifiee 1'ecologie du consortium anaerobie permettant la production de methane. Les deux etapes initiales de la digestion anaerobie sont 1'hydrolyse et 1'acidogenese qui sont effectuees par un groupement heterogene de bacteries, parmi lesquels on retrouve des anaerobes facultatifs. L'etape d'hydrolyse permet la degradation de la matiere organique complexe (polysaccharides, proteines, lipides) par les bacteries fermentaires primaires en composes tels que les oligosaccharides, acides amines et acides gras. L'acidogenese permet la prise en charge des
produits de 1'hydrolyse pour donner lieu a la production de differents acides gras volatils (AGV)
et d'alcools, d'H^ et de C02. Les AGV (autres que 1'acetate) produits lors des etapes preliminaires d'hydrolyse et d'acidogenese subissent une deshydrogenation et sont convertis en acetate par les bacteries acetogeniques productrices obligees d'H2 (OHPA). Le maintien de faibles concentrations d'Hz est essentiel a cette demiere reaction, tache accomplie par les bacteries methanogenes hydrogenothrophes qui utilisent l'H2 par la reduction du CO^, pour produire environ 30 % du methane provenant de la digestion anaerobie. La majorite du methane (70 %) est toutefois produite par les bacteries acetoclastes qui precedent par decarboxylation de 1'acetate. II est possible aux bacteries de la classe des homo-acetogenes, de produire 1'acide ac^tique a partir de I'H^ et du CO^. La Figure 2.1 montre la reaction de deshydrogenation (acetogenese) de 1'acide propionique et butyrique, la reduction du dioxyde de carbone par les methanogenes hydrogenotrophes, ainsi que la decarboxylation de 1'acetate par les methanogenes acetoclastes.
2.1.2 Avantages et desavantaees du traitement anaerobie.
L'utilisation du traitement anaerobie, et particulierement son application au secteur des p^tes et papiers, est un domaine relativement recent [LETTriMGA et coll., 1991]. Lorsque compare a son plus proche competiteur, Ie traitement aerobie en 1'occurrence, Ie traitement
CO.+H'2 -1 J-i-2
CH4
Matiere organique complexe non-soluble
(polysaccharides, proteines, lipides)
Matiere organique simple solubilisee
(oligosaccharrides, acides amines, acides gras)
Butyrate et propionate Ac^tate
CH4+C02
1 Hydrolyse
2 Acidogenese 3 Acetogenese 4 Methanogenese hydrogenotrophe 5 Methanogenese acetoclasteCH3CH2COOH + 2H^O ->• CHsCOOH + C02 +3H2 (deshydrogenation de 1'acide propionique)
CH3CH2CH2COOH + 2H20 -> 2CH3COOH + H2 (deshydrogenation de 1'acide butyrique)
C02 + 4H^ -> CH4 + H20 (methanogenese hydrogenotrophe) CH3COO- + H20 -> CH4 +HC03- (methanogenese acetoclaste)
anaerobie jouit d'avantages economiques et operationnels non negligeables [LETTLNGA et coll., 1991]. La technologie des systemes aerobies a boues activees est relativement maitrisee mais sa simplicite peut cependant etre assombrie par des couts prohibitifs relies a 1'aeration mecanique et a la disposition des importants volumes de boues generes. Le traitement anaerobie ne requiert aucune aeration, ce qui se traduit par des economies en frais de capital et d'operation (entretien et alimentation en electricite des aerateurs). De plus, la production de gaz methane par la biomasse anaerobie peut etre valorisee. La simplicite et la faible demande en espace des installations, en plus 1'economie de volume de reacteur permise par la possibilite d'utiliser de hautes charges organiques, sont egalement des avantages en la faveur des systemes anaerobies. La faible production de boue lors de la digestion anaerobie peut egalement permettre des economies au niveau des couts de stabilisation et de disposition de la biomasse produite. La biomasse anaerobie possede egalement la capacite de pouvoir rester inactive pour des periodes prolongees sans perte majeure de son activite. Le developpement de systemes anaerobie est limite par certaines contraintes pratiques: les systemes anaerobie peuvent necessiter de longues periodes de demarrage et de rodage et Ie temps d'acclimatation de la biomasse a certains composes recalcitrants peut egalement etre important. L'utilisation de systemes anaerobies est compliquee par la sensibilite des bacteries face a certains composes toxiques [SIERRA-ALVAREZ et coll., 1994]. Contrairement a 1'industrie agro-alimentaire qui genere dans la majorite des cas des effluents dont la digestion anaerobie est peu problematique, 1'industrie des pates et papiers par contre, represente un creneau d'application plus difficile a exploiter a cause de la presence de substances problematiques contenues au sein de certains effluents. Les tannins contenus dans les
effluents d'ecor9age [FIELD et LETTINGA, 1987; FIELD et coll., 1988], les lignines de faible
poids moleculaire [SIERRA ET LETTINGA, 1991b], les composes phenoliques chlores et non
chlores [SIERRA ET LETTINGA, 199 la], les acides gras a longues chames et tout specialement
les acides resiniques (AR) sont capables d'inhiber a differents degres 1'activite methanogenique des bacteries anaerobies. La prochaine section traite plus en detail de la nature des AR et des problemes environnementaux que peuvent causer ces produits.
2.2 Les acides resiniques
2.2.1 Description, provenance et structure
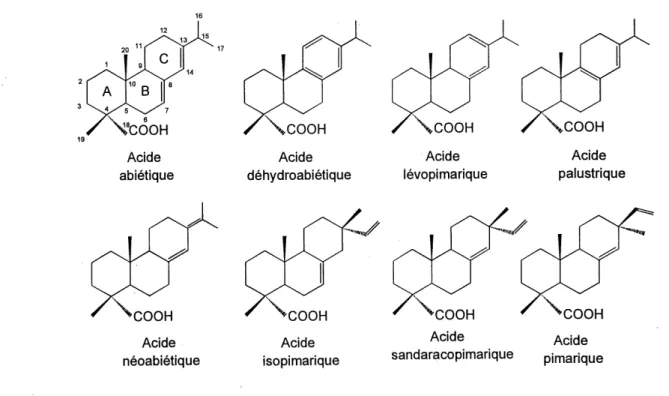
Les AR sont des acides faibles de nature hydrophobe naturellement produits par certaines especes d'arbres, les coniferes en majorite, et qui se retrouvent au sein de 1'oleoresine, de 1'ecorce et des aiguilles des arbres. Les AR sont des acides monocarboxyliques tricycliques provenant de la famille des diterpenes. Us sont presents dans la majorite des coniferes des regions temperees et comptent pour des pourcentages entre 0.2 et 0.8% de la masse seche du bois. On attribue aux AR un role de prevention contre les attaques des insectes et de leur larves [WAGNER et coll., 1983] et centre la decomposition causee par certains champignons, a 1'aide d'une combinaison d'effets
d'inhibition de croissance fongique et de controle de 1'humidite (impermeabilisation due a la
nat.ire hydrophobe des AR) [EBERHARDT et coll., 1994]. Les AR sont relaches lors des operations de mise en pate et sont une source tres importante de la toxicite des effluents d'usines de pate et papiers, comme il en sera discute a la section suivante. Les AR qui se retrouvent naturellement au sein des arbres et qui sont frequemment detectes dans les effluents de mise en
pate sont les acides abietique (AA), dehydroabietique (ADA), neoabietique, levopimarique,
pimarique, appartenant a la classe des abietanes, et les acides isopimarique, sandaracopimarique, et palustrique, qui appartiennent a la classe des pimaranes. Les membres de la classe des abietanes sont caracterises par une chame isopropyle (CH^-CH^CH-) en position C-13, tandis que ceux de la classe des pimaranes sont caracterises par des groupements methyl (-€N3) et vinyl (CH3=CH-) en cette meme position. La Figure 2.2 montre la structure des principaux AR. II peut y etre remarque que les membres de la classe des abietanes ne different d'entre eux que par la localisation des doubles liaisons, qui sont toutes endocy cliques saufpour 1'acide neoabietique, qui possede une*double liaison exocyclique. L'ADA se distingue de tous les AR par son noyau aromatique. Les membres de la classe des pimaranes different d'entre eux par la localisation de la double liaison et par la presence de a-methyl ou de P-methyl en C-13. Le present travail de recherche a porte sur deux membres de la classe des abietanes: 1'AA et 1'ADA. Les AR sont susceptibles a 1'oxydation pouvant etre causee par 1'oxygene atmospherique a temperature ambiante [LAWRENCE, 1959], en particulier 1'AA, qui au contact de 1'air, peut subir une
decarboxylation oxydative et une isomerisation causee par des reactions de radicaux libres
[SOLTES etZINKEL, 1989].
Acide abietique ^COOH Acide neoabietique Acide dehydroabietique Acide levopimarique ^COOH Acide isopimanque "COOH Acide sandaracopimarique COOH Acide palustrique ^COOH Acide pimanqueFigure 2.2 Principaux acides resiniques retrouves au sein des effluents papetiers
Les acides dehydroabietique, isopimarique et pimarique sont significativement plus
stables face a 1'oxydation [SOLTES et ZINKEL, 1989]. Certains AR sont davantage susceptibles
a differentes reactions d'isomerisation et de polymerisation pouvant modifier leur structure. Par exemple, les acides levopimarique et palustrique peuvent, par des reactions couplees d'oxydatibn
et de deshydratation, se transformer en ADA [SOLTES et ZINKEL, 1989], ce qui explique que la
composition en AR de la matiere premiere d'une usine de pate a papier puisse differer de celle de 1'effluent produit [ZENDER et coll., 1994]. La polymerisation des AR, par 1'action combinee d'acides puissants et de hautes temperatures, donne majoritairement lieu a la formation de
dimeres faiblement solubles dans des solvants organiques [SOLTES et ZESTKEL, 1989]. Les
procedes de blanchiment au chlore donnent lieu a la formation de mono- et dichloroanalogues des
influencee par 1'espece de bois utilisee. Ie precede de mise en pate, la gestion de 1'eau au sein de
1'usine [MIETTINEN et RUOPPA, 1988], la saison [HABET8 et DE VEGT, 1991] et egalement
la methode de blanchiment utilisee. Les procedes de mise en pate chimique, en particulier les precedes alcalins, permettent en plus de la solubilisation de la lignine, la solubilisation des composes de la resine [SIERRA-ALVAREZ et coll., 1991]. Les AR sont faiblement solubles dans 1'eau a pH neutre, entre 3 et 6 mg/L, [NYREN et BACK, 1958] et peuvent par consequent se retrouver adsorbes sur les fibres de pate et sur les matieres en suspension presentes dans les effluents et peuvent egalement etre incorpores a ce meme effluent sous forme de micelles [DROBOSYUK et coll., 1982]. Les AR presentent egalement une forte tendance a s'adsorber sur la biomasse autant anaerobie qu'aerobie, 1'ADA etant 1'AR Ie moins fortement associe a la
biomasse [HALL et LIVER, 1996].
2.2.2 Toxicite des acides resimgues
Les effluents des usines de pate a papier contiennent une vaste gamme de composes toxiques pour les organismes aquatiques. Parmi ces composes, se retrouvent les AR qui ont ete reconnus dans certains cas comme etant la prmcipale source de lethalite des effluents d'usines de pate a papier [LEACH et THAKORE, 1978]. La concentration d'une substance causant la mortalite chez 50% des sujets en 96 heures (CL50-96h) est un parametre communement utilise pour evaluer la toxicite d'une substance. Pour 1'AA et 1'ADA respectivement, des valeurs de CL50-96h de 0.7 et 1.1 mg/L ont ete rapportees chez la truite arc-en-ciel [LEACH et THAKORE, 1976] et des valeurs similaires ont ete obtenues pour les analogues mono et di-chlores des AR [KENNEDY et coll., 1995]. Des etudes ont demontre que des concentrations superieures a 15 mg/L d'ADA avaient pour effet de reduire Ie niveau d'ATP cellulaire et la consommation d'oxygene, causant une augmentation de 1'hemolyse des erythrocytes (globules rouges) et Ie
developpement dejaunisse chez les poissons [MATT80FF et OIKARI, 1987]. Bien que les cas
de mortalite aigue de poissons causee par les effluents d'usine de pate soient peu frequents [TANA, 1988], les effets chroniques des polluants se retrouvant a des concentrations sous-lethales au sein des milieux recepteurs doivent toutefois etre consideres. Des tests effectues sur des truites arc-en-ciel ont montre que la plupart des AR et certains de leurs chloroanalogues sont bio-accumules, mais peuvent toutefois etre elimines par ces poissons (duree de demi-vie estimee
de 4 jours) [NIIMI et LEE, 1992]. L'ADA, a partir d'une concentration de 5 p-g/l, peut
s'accumuler au sein des tissus du poisson, en particulier au niveau du foie, et affecter differentes fonctions metaboliques [TANA, 1988]. Des echantillonnages effectues en aval d'usines de pate a papier ont montre de faibles concentrations en AR et en derives d'AR [BROWNLEE et coll., 1977; OWENS et coll., 1994; JUDD et coll., 1996] au niveau des sediments, avec une predominance de 1'ADA retrouve a des concentrations entre 5 et 100 p,g/g dans les premiers 10 cm de sediments [BROWNLEE et coll., 1977]. Des etudes effectuees avec des moules d'eau douce ont montre que les AR sont rapidement bio-accumules au sein des tissus, mais lorsque les moules sont reintroduites dans de 1'eau pure, la demi-vie moyenne des AR au sein de ces
demieres est d'environ 3 jours [BURGGRAAF et coll., 1996].
Les donnees sur la lethalite aigue et les effets chroniques des AR sur les poissons mettent en evidence la necessite de trailer les effluents afin d'en reduire la teneur. Les prochaines sous-sections proposent un aper^u des differents types de traitement utilises par 1'industrie papetiere a la lumiere de leur performance d'enlevement des AR.
2.2.3 Traitementaerobied'effluentscontenantdes acides resimgues
Les traitements biologiques aerobies conventionnels tels que les boues activees et les etangs aeres sont consideres comme des methodes efficaces pour 1'elimination des AR des effluents de mise en pate [LIU et coll., 1996] meme si ces precedes peuvent occasionellement afficher des performances variables d'enlevement des AR [ZENDER et coll., 1994]. A titre d'exemple. Ie traitement par etang acre (temps de retention hydraulique de 4.5 jours) d'un
effluent kraft a pennis la reduction de la concentration totale en AR de 95 % [STUTHRIDGE et
coll., 1991]. Un traitement par boues activees (temps de retention hydraulique de 8 heures) a pu effectuer un enlevement de 92% des acides gras et resiniques contenus dans 1'effluent d'une usine de mise en pate par precede chimico-thermomecanique (CTMP) [LIU et coll., 1996]. II a ete montre que 1'enlevement global des acides gras et resiniques lors de cet essai etait du majoritairement a une action biologique mais a egalement ete cause par des phenomenes d'adsorption sur la biomasse et d'oxydation chimique causee par 1'aeration [LIU et coll., 1996]. Des essais effectues en mode cuvee ont montre qu'une boue aerobie municipale non acclimatee aux AR a pu enlever completement 200 mg/L d'un melange d'AR en moins de 2 jours
(1'adsorption sur la biomasse a ete mesuree et comptabilisee), 1'ADA etant enleve a un taux semblable sinon superieur aux autres AR etudies [LIVER et HALL, 1996]. Une voie metabolique aerobie partielle de 1'AA passant par 1'hydrogenation et la saturation ainsi que par la formation de derives hydroxyles (en la position C-13) et decarboxyles a ete proposee [ZENDER
et coil, 1994].
2.2.4 Traitement anaerobie d'effluents contenant des acides resiniques
Bien que les AR soient presents dans un grand nombre d'effluents issus de 1'industrie fbrestiere, 1'effet precis de ces substances sur les bacteries anaerobies et leur traitabilite par les systemes anaerobies sont encore peu connus [MCCARTHY et coll., 1990]. II semble qu'aucun consensus n'ait €ie atteint quant au devenir des AR traites par voie anaerobie. MCFARLANE et CLARK [1988] ont montre que les AR peuvent etre degrades dans un reacteur anaerobie ascentionel a lit granulaire (Uplo^ anaerobic sludge blanket reactor, UASB) (2.5 ans d'acclimatation a 1'effluent) a 1'exception de 1'ADA, qui se trouve concentre au sein de la biomasse. Des resultats similaires ont egalement ete publics par PATOINE et coll. [1997]. La nature co-metabolique de la degradation de 1'AA par un consortium anaerobie a ete mise en evidence lors du retrait de la source de carbone (galactose), moment a partir duquel la
degradation de 1'AA s'est trouvee interrompue [MCFARLANE et CLARK, 1988]. Dans des
etangs de traitement a predominance anaerobie, la reduction de la concentration totale en AR d'un effluent kraft a ete limitee a 44% et des concentrations importantes en ADA et en un derive
hydroxyle de 1'AA ont ete mesurees a la sortie du traitement [STUTHRIDGE et coll., 1991].
LIVER et HALL [1996] n'ont pu observer aucune degradation significative des AR lors d'essais
en bouteille realises avec une boue anaerobie non acclimatee aux AR (test d'une duree de 24 jours).
Les AR, en plus de poser une menace aux organismes vivants des milieux recepteurs, possedent egalement une toxicite intrinseque envers Ie consortium bacterien du traitement anaerobie. Les nouveaux procedes CTMP utilisent des quantites reduites d'eau, ce qui resulte en des effluents plus concentres et davantage propices a un traitement anaerobie [WELANDER et ANDERSSON, 1985]. Certains de ces effluents se sont montres inhibiteurs envers la production de methane, en raison de la solubilisation chimique de composes resineux toxiques [LETTINGA
et coll., 1991; MCCARTHY et coll., 1990]. II a ete demontre que la toxicite anaerobie des effluents CTMP est causee a la fois par des composes presents dans la phase aqueuse, et egalement par des composes adsorbes aux fibres contenues dans 1'effluent [MCCARTHY et coll., 1990; RICHARDSON et coll., 1991], d'ou la necessite d'un controle adequat des matieres solides
pour un traitement anaerobie efficace [HABETS et DE VEGT, 1991; RICHARDSON et coll.,
1991]. MCCARTHY et coll. [1990] ont demontre que Ie lavage de ces fibres avec du methanol a permis d'en eliminer completement la toxicite anaerobie. Les memes auteurs desirant connaitre la part de responsabilite des AR pour la toxicite anaerobie des effluents blanchis CTMP, out conclu que les AR inhibent 1'activite methanogene mais que la toxicite anaerobie de ces effluents ne peut etre expliquee qu'en partie par les AR: ils postulent un effet de synergie cause par la combinaison des AR a d'autres composes non identifies [MCCARTHY et coll., 1990]. II a egalement ete montre que la toxicite anaerobie de certains effluents de mise en pate peut etre eliminee par une extraction a 1'ether (solubilisation de composes non polaires) ou par adsorption sur resine polymerique, qui sont deux methodes d'enlevement selective des composes de la resine des arbres [SIERRA-ALVAREZ et coll., 1991]. Plusieurs auteurs ont remarque une baisse de 1'activite methanogenique des systemes anaerobies en presence de divers representants des AR.
WELANDER et ANDERSSON [1985] et FIELD et coll. [1988] ont rapporte des inhibitions
jusqu'a 50% de 1'activite de controle causees par la presence d'AA tandis que des essais en reacteur (UASB) avec 1'acetate comme seule source de carbone, out montre que d'importantes concentrations en AA maintenues a long terme n'ont pas eu d'effet significatif sur 1'activite des bacteries methanogenes acetoclastes et sur la performance globale du reacteur [KENNEDY et coll., 1992]. Des essais effectues en bouteille ont demontre que 1'inhibition de 1'activite methanogenique etait proportionnelle a la charge specifique en AR (masse d'AR par masse deMVS) [MCCARTHY et coll., 1990; KENNEDY et coll., 1992; LIVER et HALL, 1996]. A titre
d'exemple, 1'inhibition de 50% de 1'activite methanogenique a ete obtenue a une concentration
specifique en AR de 80 mg/g MV8 [KENNEDY et coll., 1992]. Le caractere irreversible de
1'inhibition methanogenique cause par les AR a ete montre par HABETS et DE VEGT [1991]
ainsi que par PATOBSTE et coll. [1997]. Certains auteurs out suggere une certaine capacite
d'adaptation du consortium anaerobie face aux AR [MCFARLANE et CLARK, 1988] et a un
effluent CTMP initialement inhibiteur envers la microflore methanogene [WELANDER et
ANDERSSON, 1985; RICHARDSON et coll., 1991]. 11 a ete demontre que les bacteries
methanogenes acetoclastes sont inhibees par les AR, contrairement aux bacteries methanogenes hydrogenotrophes dont 1'activite est restee inchangee au sein d'un reacteur UASB soumis a des AR [PATODSTE et coll., 1997]. La prochaine sous-section traite de certains procedes physico-chimiques permettant 1'enlevement des AR dans 1'optique d'une reduction de leur toxicite prealablement au traitement anaerobie.
2.2.5 Modification et pretraitement phvsico-chimiaue des acides resmiques
Parmi les methodes physico-chimiques d'enlevement des AR, la precipitation par sel de
calcium (CaC^) en condition alcaline (pH 11) a permis 1'enlevement de 1'AA [FIELD et coll.,
1988] et de differents composes inhibant la methanogenese [SIERRA-ALVAREZ et coll., 1991].
Un rapport inteme de la compagnie Paques B.V. cite dans HABETS et DE VEGT [1991] a
montre que 1'abaissement du pH d'un effluent CTMP a 1.5 a permis la precipitation des AR, reduisant ainsi leur concentration de 95.3 mg/L a 0.2 mg/L, permettant une reduction significative de 1'inhibition de la methanogenese causee par les AR. Des resultats similaires ont ete obtenus pour des effluents de mise en pate a la soude [SIERRA-ALVAREZ et coll., 1991]. Pour 1'operation d'un reacteur anaerobie industriel, la dilution d'un effluent CTMP avec des eaux blanches et la recirculation de 1'effluent du reacteur ont ete citees comme etant des methodes efficaces permettant la reduction de la toxicite pour les bacteries anaerobies [HABETS et DE
VEGT, 1991].
2.2.6 Degradation bjologiaue des acides resmiaues par des cultures pures
Les sous-sections 2.2.3 et 2.2.4 ont traite de la degradation des AR par des procedes aerobies et anaerobies utilisant une biomasse renfermant un large spectre de micro-organismes. Des etudes de biodegradation des AR ont aussi ete effectuees a 1'aide de souches bacteriennes pures. La degradation bacterienne de 1'ADA par Flavobacterium resinovorum [BIELLMANN et coll., 1973a] et par des bacteries du genre Pseudomonas et Alcaligenes [BIELLMANN et coll., 1973b] qui utilisent 1'ADA comme seule source de carbone a ete rapportee. Les auteurs de ces travaux ont identifies plusieurs sous-produits oxydes provenant de 1'ADA permettant ainsi la
suggestion de votes metaboliques d'interet secondaire puisque ces micro-organismes n'ont pas ete isoles a partir d'effluent ou de systeme de traitement propres a 1'industrie papetiere. COTE et
OTIS [1989] ont etudie la bacterie Bacillus psychrophilus qui a ete isolee a partir de clariflcateurs
de papeterie et qui a demontre la capacite de degrader 1'ADA par co-metabolisme. Recemment, des bacteries pouvant croitre a partir de 1'ADA ont ete isolees dans un reacteur sequentiel traitant des eaux de precede concentrees d'une usine de mise en pate dans Ie but de mieux identifier et comprendre les facteurs pouvant agir sur la biodegradation des AR au sein des systemes de traitement aerobies [MOHN, 1995]. Les bacteries isolees (semblent correspondre a Sphingomonas yanoikuyae et Zoogloea ramigera) ont egalement ete capables de croitre en utilisants d'autres AR de la classe des abietanes tels que 1'acide abietique et palustrique mais ont ete incapables de croitre a partir des membres de la classe des pimaranes, comme les acides pimarique et isopimarique [MOHN, 1995]. Deux autres types de bacteries capables de croitre en utilisant les acides pimariques et isopimariques comme seule source de carbone ont ete isoles a partir du meme reacteur sequentiel, ce qui laisse croire une certaine specificite lors de 1'enlevement des AR par les systemes aerobies sequentiels [WILSON et coll., 1996]. D'autres souches bacteriennes capables d'effectuer 1'enlevement des AR ont ete isolees a partir de bassins
d'effluent de mise en pate kraft blanchie [MORGAN et WYNDHAM, 1996; BICHO et coll.,
1995]. Les deux souches les plus performantes isolees par BICHO et coll. [1995] ont ete etudiees pour leur capacite a degrader les AR contenus dans un effluent CTMP. L'effluent CTMP a permis la croissance des deux bacteries mais les nutriments qui se sont averes necessaires a la degradation des AR par ces bacteries (NH4T, ?0^~, mineraux et vitamines) se trouvent en trop
faibles quantites dans ce type d'effluent [ZHANG et coll., 1997].
II existe egalement plusieurs champignons capables de transformer les AR, autant les abietanes que les pimaranes, mais il n'existe aucune preuve de degradation fongique de ces composes [WILSON et coll., 1996]. De plus, ce type de substrat ne serait pas preferentiel aux champignons de la poumture du bois en comparaison a la lignine, les sucres simples et les polysaccharides egalement contenus au sein de cette matrice [EBERHARDT et coll., 1994]. Le zygomycete Mortierella isabellina est Ie champignon dont la capacite a transfonner les AR a ete Ie plus abondamment documentee. Ce champignon agit prmcipalement par hydroxylation (en
C-2 sur Ie cycle A et en C-15 et C-16 sur la chaine isopropylique) des AR (AA, ADA et acide
isopimarique) [KUTNEY et coll., 1981] et egalement de leurs mono et di-chloroanalogues, sans
toutefois attaquer les cycles, ni effectuer de dechlorination [KUTNEY et coll., 1982; KUTNEY et
coll., 1983]. Bien qu'il n'y ait pas de degradation proprement dite de la structure des AR, il a ete verifie que 1'hydroxylation de ces composes en reduit significativement la toxicite aigue (especes testees: Daphnia pulex et tmite arc-en-ciel) [SERVIZI et coll., 1986]. II a ete demontre que Ie
degre et Ie site de 1'hydroxylation sont des facteurs affectant la reduction de la toxicite [SERVIZI
et coll., 1986]. Toutefois, la germination des spores de ce champignon est impossible au sein des effluents des usines de pate kraft (carences en azote et en sucres) et Ie champignon est un faible competiteur dans des systemes microbiens tels que les etangs de traitement [KUTNEY et coll., 1985]. D'autres souches fongiques peuvent enlever les AR a partir de la matrice meme du bois lorsque celui-ci est coupe. Parmi celle-ci, se retrouvent des champignons saprophytiques du bois, notamment ceux de la tache de 1'aubier (sap staining fungus) comme les especes Ceratocystis et Ophiostoma, capables de coloniser rapidement les canaux de resine sans alterer les elements stmcturaux du bois (lignine, cellulose et hemicellulose). Jusqu'a tout demierement, ces souches etaient jugees indesirables a cause de la coloration bleutee qu'elles pouvaient apporter aux copeaux de bois destines a la mise en pate mais il a etc montre que ces champignons peuvent egalement se reveler utiles a cause de leur propension a utiliser certains composes de la resine (triglycerides, acides gras et resiniques). Ces substances, composant la majeure partie des matieres extractibles du bois, connues sous Ie nom de poix (pitch en anglais), peuvent causer des problemes majeurs lors de la fabrication du papier a cause de leur tendance a former des depots sur les machines a papier pouvant ainsi nuire a la qualite du produit fmi [BLANCHETTE et coll.,
1992]. Dans Pindustrie forestiere et papetiere, on defmit par matieres extractibles, 1'ensemble des composes du bois solubilises (cires, alcools, acides gras et resiniques, triglycerides) par des solvants organiques non polaires (acetone ou dichloromethane). Des souches incolores
disponibles sur Ie marche sous Ie nom de CARTAPIP (souche albinos de Ophiostoma
piliferum) (Sandoz Chemicals Corp., Charlotte, NC) peuvent etre utilisees sur les piles de copeaux de bois prealablement a la mise en pate afin d'en enlever les substances extractibles dont font partie les AR. Des essais effectues avec Ophiostoma piceae sur des copeaux de bois ont
permis de determiner que la reduction de la teneur en matieres extractibles causee par ce champignon etait en partie due a 1'enlevement des AR: pour une periode d'incubation de trois semaines, Ie traitement fongique a permis un enlevement global de 63% des AR, dont des enlevements de 50 et 57% pour 1'ADA et 1'AA respectivement [GAO et coll., 1994]. D'autres resultats ont montre la capacite des champignons CARTAPIPMC, 0. picea, 0. ainoae (preleves sur des coniferes) et Lecythophora sp. (preleve sur du peuplier) a enlever plus de 67% des AR contenus dans des copeaux de pin en deux semaines, sans qu'un traitement prolonge ne permette de completer 1'enlevement [WANG et coll., 1995]. Lors de cet essai, la teneur en AA et en ADA des copeaux a ete reduite de 80% et de 40% pour les quatre souches etudiees. Les mecanismes, les sous-produits et les effets ecotoxicologiques de ce traitement ne sont cependant pas connus.
La presente etude a pour but 1'evaluation de la performance de 1'enlevement des AR par Ie champignon de la pourriture blanche du bois P. chrysosporium, dont les caracteristiques physiologiques different significativement de celles des champignons de la tache de 1'aubier. La prochaine section dresse un portrait succinct de ce champignon, en particulier ses capacites oxydatrices et ses potentialites au niveau environnemental et papetier.
2.3 Phanerochaete chrysosporium
2.3.1 Orifiine et description
Environ 1600 souches de champignons prenant part a la poumt-ire du bois ont ete identifiees. Elles sont divisees en trois grandes categories: la pourriture molle, blanche et brune, faisant reference non pas a 1'aspect visuel exterieur du champignon, mais plutot aux residus de leur action [GLASER, 1988]. P. chrysosporium est un champignon microscopique filamenteux causant la poumt.ire blanche du bois et il est retrouve partout sur 1'hemisphere nord [LEWANDOWSKI et coll., 1990]. P. chrysosporium appartient a la sous-classe des basidiomycetes, cette demiere etant caracterisee par la production de spares extemes portees au sommet d'un pedoncule nomme baside [BUR8DALL et ESLYN, 1974]. Les champignons sont des organismes eukaryotes, c'est a dire pourvus d'une membrane nucleique, et bien que compris dans Ie regne des plantes, ne possedent aucune activite photosynthetique puisqu'ils sont depoiu-vus de chlorophylle [GLASER, 1988]. Les champignons de la pourrit-ire blanche causent un type particulier de decomposition au cours de
laquelle Ie bois est blanchi ou decolore par 1'action du champignon sur la lignine [BARCLAY, 1991]. Au niveau microscopique, les hyphes de ces champignons colonisent les lumens du bois et penetrent d'abord de cellule en cellule et ultimement via des reseaux de cavites qui sont pratiques a travers Ie bois [BARCLAY, 1991]. Afin d'acceder a la cellulose, qui represente sa principale source de carbone. Ie champignon est capable de secreter des enzymes extracellulaires qui lui permettent
de degrader la lignine, qui est un compose tres resistant a la biodegradation [LEWANDOWSKI et
coll., 1990]. La structure tres complexe et variable de la lignine requiert de tout micro-organisme aspirant a la degrader un systeme enzymatique extracellulaire dote d'une faible specificite. C'est d'ailleurs cette faible specificite pour une large gamme de substrat (autres que la lignine) qui a permis a P. chrysosporium de se meriter 1'interet qui lui est voue dans Ie domaine de la biodegradation des polluants recalcitrants [FERNANDO et coll., 1990]. Les prochaines sous-sections decrivent brievement la structure de la lignine ainsi que les mecanismes connus par lesquels P. chrysosporium s'y attaque.
2.3.2 Description de la structure de la lienine
La lignine est un polymere naturel tridimensionnel servant a rigidifier la structure principale de plusieurs vegetaux (arbres en particulier). La structure des plantes superieures est egalement constituee de cellulose (compose organique Ie plus repandu sur la planete) et d'hemicellulose. La formation de la lignine est catalysee par la polymerisation oxydative des radicaux libres des alcools p-hydroxycinnamyliques de structure phenolique: alcools p-coumarylique, coniferylique et sinapylique, illustres a la Figure 2.3. A cause de la variabilite de la quantite et du type des alcools precurseurs ainsi que de leur biosynthese non stereospecifique, la lignine est un polymere dont la sti-ucture peut s'averer heterogene [BUMPUS et AUST, 1987]. Ce manque d'homogeneite stmcturale couple a la nature racemique du polymere font de la lignine un compose resistant a la plupart des systemes enzymatiques [BUMPUS et AUST, 1987]. Les liaisons stables carbone-carbone (C-C) et carbone-carbone-oxygene (C-0), ainsi que les liaisons entre les noyaux aromatiques constituent Ie maillage de la lignine. La taille des molecules de lignine oscille entre 600 et 1000 kDa, rendant impossible son transfert au sein des membranes cellulaires de micro-organismes par les mecanismes connus d'echange [GLASER, 1988]. P. chrysosporium degrade la lignine via un systeme enzymatique complexe et encore assez mal caracterise.
OCHa HaCC
CHzOH CH CH
-OCH
Alcool p-coumarylique Alcool coniferylique Figure 2.3 Alcools phenoliques precurseurs de la lignine
OH
Alcool sinapylique
Les enzymes responsables de 1'action preliminaire du champignon (appelees enzymes ligninolytiques) sont de nature extracellulaire, tandis que les mecanismes intracellulaires pennettant la mineralisation des fragments produits lors de la depolymerisation sont peu coimus. La prochaine sous-section traite des mecanismes d'action des enzymes ligninolytiques lignine et manganese peroxydases.
2.3.3 Mecanismes d'action des lienine et manganese peroxvdases
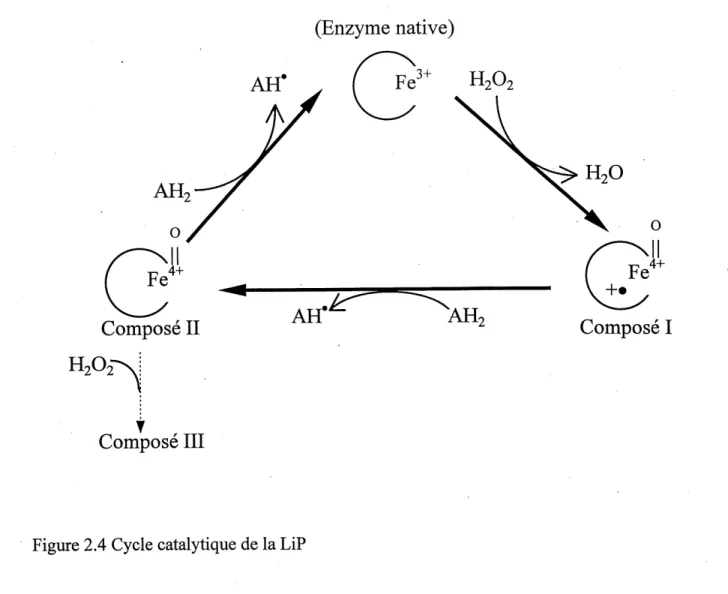
Le systeme ligninolytique de P. chrysosporium implique les enzymes lignine peroxydase (LiP) et manganese peroxydase (MnP). Ces demieres sont en partie responsables de 1'activite ligninolytique du champignon, c'est a dire de la capacite globale de degrader la lignine. Cette activite est difficilement quantifiable. Pour cette raison, des indicateurs tels que les activites enzymatiques LiP et MnP (dont Ie protocole de dosage est explique en detail aux Annexes 1 et 2) sont utilises. On a annonce en 1983 1'isolation de la LiP, hemoproteine (contenant du fer) obtenue a partir de cultures ligninolytiques de P. chrysosporium dont Ie poids moleculaire peut vaner entre 38
et 42 kDa (selon 1'isoenzyme) [TIEN et KIRK, 1983; GLENN et coll., 1983]. La LiP permet
1'oxydation des anneaux aromatiques non phenoliques [KERT8EN et coll., 1985] et egalement Ie
clivage de liens C-C et C-0 et possiblement C-C1 [BUMPUS et AUST, 1987] au sein de la lignine.
I'ouverture des amieaux aromatiques, la demethoxylation, la formation de radicaux libres
hydroxyles (par 1'action du H'^Pi) capables de degrader la lignine [FORNEY et coll., 1982], la
decarboxylation [AKAMATSU et coll., 1990; HARVEY et coil, 1986] et des reactions de
couplage phenolique [SCHOEMAKER et LEISOLA, 1990]. La LiP permet 1'oxydation par un
electron de substrats aromatiques en presence de peroxyde d'hydrogene. L'enzyme LiP peut effectuer a elle seule un certain nombre d'oxydations par elle-meme, mais Ie systeme LiP implique aussi 1'alcool veratrylique (AVE), 1'oxalate, Ie peroxyde d'hydrogene. Ie fer ferrique et ferreux ainsi
que des radicaux hydroxyles [BARR et coll., 1992]. L'activite de la LiP est dependante de la
presence de peroxyde d'hydrogene comme son nom 1'indique. Ce compose sert d'accepteur d'electron pour la LiP. HARVEY et coll. [1986] ont propose que 1'AVE, un metabolite secondaire du champignon [SHIMADA et coll., 1981], puisse semr de mediateur entre 1'enzyme LiP et les matrices-polymere moins accessibles a 1'enzyme et de ses sites actifs. Concretement, cela signifie que la LiP transforme 1'AVE en radical cationique, une espece oxydante active capable d'agir directement sur Ie substrat [HARVEY et coll., 1986]. L'AVE, lorsqu'ajoute a des cultures, a permis d'augmenter Ie taux initial de mineralisation de la lignine par P. chrysosporium [LEI80LA
et coll., 1984; FAISON et KIRK, 1985]. D'autres recherches [HARVEY et coll., 1989] ont permis
de mieux defmir Ie role de 1'AVE. La Figure 2.4 illustre Ie cycle catalytique de la LiP. En premier lieu, la LiP est oxydee (deux electrons) par Ie R^OI secrete par Ie champignon pour former un radical cationique (compose I). Ce radical cationique subit ensuite la reduction (un electron), entrainant ainsi 1'oxydation d'un substrat (AH^ sur la figure) et la formation du compose II de la LiP. Apres un certain nombre de reductions, 1'enzyme transforme possede un plus faible niveau d'activite catalytique. L'AVE (AH^ sur la figure) permet a 1'enzyme transfomie (compose II) de se reconvertir en son etat natifet de produire un radical cationique de 1'AVE (AVE). En absence de substmt et en presence de HzOz, 1'enzyme perdra son activite catalytique par sa conversion irreversible en compose III, qui peut egalement etre obtenu a partir de 1'enzyme native en exces de
H^.
La MnP (poids moleculaire d'environ 46 kDa) est une enzyme egalement produite par P. chrysosporium et plusieurs autres champignons de la pourritire blanche [KUWAHARA et coll.,
AH
Compose II
HzOr
Compose III
(Enzyme native)
AH*
G
Fe3+ H^O
.2^2AH.^"
VAH.H.O
0
^~^\
4+ +•Compose I
Figure 2.4 Cycle catalytique de la LiP
La MnP requiert du R^O^ qui oxyde 1'enzyme native en compose I, capable de catalyser 1'oxydation
-2+ -3+
du Mn en Mn. Le compose II ainsi obtenu peut ensuite retoumer sous la forme native de 1'enzyme par une autre oxydation du Mn^. Le MnJT peut, s'il est stabilise par des agents chelateurs (acides organiques), former des complexes capables de diffuser a partir du site actif de 1'enzyme et
d'oxyder des composes phenoliques de la lignine [TUOR et coll, 1992;WARIISHI et coll., 1989].
La particularite de la MnP est qu'elle permet 1'oxydation des sous-stmctures phenoliques de la lignine. La LiP et la MnP peuvent agir en synergie et oxyder une large gamme de composes. Les sous-produits de cette degradation sont des acides aromatiques et des aldehydes qui seront ensuite dives et mineralises ou qui subiront un cycle d'oxydoreduction les transformant en quinones et
des especes reactives de 1'oxygene comme Ie N202, Ie radical hydroxyle (OH') et 1'anion superoxyde (02 ) au cours de la degradation de la lignine par P. chrysosporium a ete mise en evidence avant meme 1'isolation de la LiP et de la MnP puisque la minemlisation de la lignine au sein de cultures fongiques a pu etre inhibee par 1'ajout de cytochrome c ou de superoxyde dismutase (connus pour stopper 1'activite du superoxyde) ainsi que par Fajout de benzoate et de salicylate (connus pour
stopper Factivite du radical hydroxyle) [FAISON et KIRK, 1983]. La production du radical
hydroxyle au sein de cultures de P. chrysosporium a ete demontree par BARR et coll. [1992].
AH"
CHELATEUR> , ( Fe3+ H^O,
(Enzyme native)
n^
H^O
L^;
Compose II
,...Mn CHELATEUR^
Mn
2+Compose I
AH-Figure 2.5 Cycle catalytique de la MnP
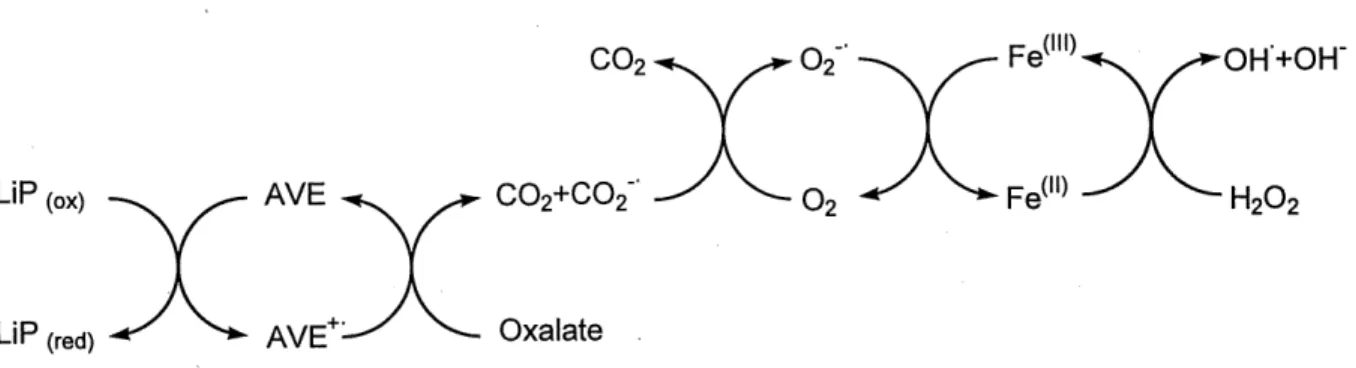
2.3.4 Rolede Facide oxaliaue
Les champignons de la pourriture du bois peuvent produire de 1 acide oxalique et 1'AVE. II a ete demontre lors d'essais in vitro que 1'oxalate inhibe la reaction de production de
veratraldehyde (par 1'oxydation de 1'AVE catalysee par la LiP) [AKAMATSU et coll., 1990].
Cette inhibition non-competitive resulte du fait que Ie radical cationique AVE est retransfomie en AVE (plutot qu'en veratraldehyde) apres avoir oxyde 1'oxalate. Ce resultat est en accord avec Ie role de mediateur propose pour 1'AVE lors de 1'oxydation de substrats par la LiP car la
presence d'AVE est necessaire a la mineralisation de 1'oxalate par la LiP. L'action indirecte de la LiP sur 1'acide oxalique explique aussi que ce produit ne soit pas accumule au sein des cultures de champignons de la pourriture blanche [AKAMATSU et coll., 1990]. II a ete propose que 1'acide oxalique puisse semr lors de la generation d'especes reactives de 1'oxygene, en particulier les radicaux hydroxyles comme Ie montre la Figure 2.6 [BARR et coll., 1992]. Cette reaction serait impossible sans la presence des metabolites secondaires AVE et oxalate.
Fe( ")^ ^-OH'+OH"
C02+C02--^ ^-0.^ ^Fe(ll)-^ ^H.O12^2
LiP (red) -^ ^ AVE+:^ ^-- Oxalate
Figure 2.6 Mecanisme propose de production du radical hydroxyle par la LiP
2.3.5 Production du peroxyde d'hvdro gene
Puisque 1'activite des en2ymes LiP et MnP depend de la presence de H^, Ie champignon doit done Ie produire lui-meme car ce produit n'est pas disponible dans son milieu naturel et encore moins au sein des cultures en laboratoire (sauf si ajoute). Des auteurs proposent que Ie H^Oz est produit par 1'incorporation de 1'oxygene contenu dans 1'air aux radicaux fonnes par la catalyse de la LiP [HAMMEL et coll., 1985]. Plusieurs enzymes capables de catalyser la production de H^Oz ont ete detectees au sein de cultures de differents champignons de la pourriture blanche, incluant P.
chrysosporium [ERIK8SON et coll., 1986; DANIEL et coil, 1994].
2.3.6 Conditions de production des enzymes lieninolvtiaues
Bien que les mecanismes de regulation de la production des enzymes ligninolytiques ne soient pas completement elucides, il existe un grand nombre de publications portant sur 1'optimisation de la production d'enzymes ligninolytiques afin d'ouvrir la voie a leur utilisation
industrielle et environnementale. Parmi celles-ci se degagent trois axes prmcipaux de recherche: la selection des souches et des conditions physiologiques optimales. Ie choix de la configuration et du mode d'operation des bioreacteurs et finalement 1'etude de 1'ajout de substances permettant la regulation et 1'hyperproduction des enzymes ligninolytiques.
Les temperatures rapportees pour une croissance optimale de P. chrysosporium se situent entre 37 et 40 C mais on a rapporte une temperature optimale de production d'enzyme de 30°C
[ASTHER et coll., 1988]. Le pH optimal pour 1'activite ligninolytique du champignon se trouve
entre 3.5 et 5.5. Les enzymes ligninolytiques sont majoritairement produites lors de la phase de metabolisme secondaire (phase idiophasique) en reponse a des carences en azote [KEY8ER et coll.,
1978], en carbone ou en soufre [IEFFRIES et coll., 1981]. L'activite ligninolytique est declenchee par de restrictions en nutriments et ce, sans egard a la presence ou 1'absence de substances a degrader [KEYSER et coll., 1978]. II a toutefois ete montre que lorsque Ie glycerol est utilise comme source de carbone (contrairement au glucose), les peroxydases peuvent apparaitre au cours de la phase primaire de croissance, et ce dans des conditions non limitantes en azote [ROCH et coll., 1989].
Le choix de la configuration du reacteur joue un role primordial pour la production d'enzymes. II s'en degage que les reacteurs permettant I'immobilisation du champignon [ASTHER
et coll, 1990], la minimisation des contraintes de cisaillement [VENKATADRI et IRVINE, 1990],
tout en permettant la maximisation des transferts d'oxygene [FAISON et KIRK, 1985] doivent eti-e
privilegies.
II a ete montre que certains produits, lorsqu'ajoutes aux cultures de P. chrysosporium, permettent 1'augmentation des activites ligninolytiques LiP et MnP; parmi ceux-ci Ie surfactant
anionique Tween 80 et 1'acide oleique [ASTHER et coil, 1987], 1'AVE [FAISON et KIRK, 1985],
differents phospholipides du soja [ASTHER et coll., 1988; CAPDEVILA et coll., 1990] et I'inositol
[MOUKHA et coll., 1991]. L'utilisation de concentrations elevees de manganese (40 ppm) pennet
1'hyperproduction selective de la MnP cependant au detriment de la LiP [BONNARME et
JEFFRIE8,1990].
Tel que mentionne precedemment P. chrysosporium et/ou ses enzymes ligninolytiques sont 1'objet d'une large gamme d'etudes visant 1'integration de ces biotechnologies au sein de certains secteurs d'activite economique. La prochaine section traite de ce sujet.
2.4 Applications potentielles de P. chrysosporium
2.4.1 Degradation de conmosesrecalcitrantsnar P. chrvsQsporium
Une large gamme de composes recalcitrants peuvent etre degrades et mineralises par P. chrysosporium, mettant en evidence la non specificite de son systeme enzymatique. Les references suivantes proposent des revues exhaustives a ce sujet: BARCLAY [1991], REDDY
[1994], PASZCZYNSKI et CRAWFORD [1995]. Parmi les groupes de composes recalcitrants
pris en charge par P. chrysosporium, on retrouve les hydrocarbures polycycliques aromatiques
(HAP) [BUMPUS, 1989; BARCLAY, 1991] pouvant etre degrade a 70-100 % (au sein d'un
milieu liquide) en 27 jours entrainant la production de gaz carbonique (15 % de mineralisation en 7 jours) et de metabolites solubles dans 1'eau. Des composes chlorophenoliques comme Ie pentachlorophenol (PCP) et des guaiacols polychlores peuvent etre degrades par Ie champignon
via dechlormation [ALLEMAN et coll., 1994] et mineralises [BUMPUS et AUST, 1987;
LAMAR et DIETRICH, 1990]. Les tristement celebres BPC (biphenyles polychlores) peuvent
etre significativement mineralises par des cultures ligninolytiques de P. chrysosporium [BATON,
1985; THOMAS et coll., 1992]. Les herbicides atrazine [MOUGIN et coll., 1994] et alachlor
[FERREY et coll., 1994] ainsi que I'insecticide DDT [FERNANDO et coll., 1989] sont d'une
autre classe de composes industriels pouvant etre biodegrades par P. chrysosporium, tout comme les teintures azo commerciales qui peuvent etre degradees et decolorees efficacement [PASZCZYNSKI et coll., 1991]. Finalement, P. chrysosporium compte parmi 1'un des rares micro-organismes connus capable de mineraliser a des niveaux eleves (jusqu'a 50 %) des composes energetiques recatcitrants tel que Ie trinitrotoluene (TNT) [FERNANDO et coll.,
1990].
II a ete montre que la LiP peut catalyser 1'oxydation de certains composes phenoliques
s'attaquer a certains HAP qui ne sont pas des substrats de la LiP [BOGAN et coll., 1996]. Certaines etudes on montre la necessite du maintien de conditions ligninolytiques pour la degradation de certains composes [FERNANDO et coll., 1990] alors que la biodegradation d'autres composes par P. chrysosporium peut etre effectuee meme lorsque les activites LiP et MnP sont supprimees [YADAV et REDDY, 1993]. La connaissance et Ie controle des mecanismes enzymatiques et physiologiques impliques lors de la biodegradation fongique des composes recalcitrants pourrait ouvrir la voie a des systemes efficaces et polyvalents de depollution. Cependant, il s'avere que les mecanismes de detoxification par voie fongique, encore peu caracterises, sont de nat.u-e multiple et
ne se limitent pas uniquement a la LiP et a la MnP [BARR et AUST, 1994]. Certains composes qui
ne sont pas des substrats de la LiP et de la MnP peuvent etre mineralises par P. chrysosporium. C'est Ie cas du TNT, un compose hautement oxyde, substrat peu probable pour des enzymes de nature oxydative comme la LiP et la MnP. II a ete montre que Ie TNT est initialement reduit au sein de la membrane plasmique du champignon, pour former des metabolites servant de substrat a la UP
[BUMPUS et TATARKO, 1994; MICHELS et GOTTSCHALK, 1994]. 11 a aussi ete montre qu'un
metabolite fongique principal du DDT est obtenu par dechlorination reductive [BARR et AUST,
1994]. Tout comme pour la lignine, 1'implication de radicaux libres et d'especes fortement reactives de 1'oxygene dans les reaction de biodegradation des polluants est connue [BARR et AUST, 1994]. Le systeme enzymatique non specifique et en partie extracellulaire de P. chrysosporium lui confere la capacite de degrader une vaste gamme de produits recalciti'ants, solubles ou non dans 1'eau. Le systeme enzymatique est controle par les conditions de culture et possede done un meilleur potentiel pour la degradation jusqu'a disparition presque complete des polluants, contrairement a d'autres micro-organismes qui utilisent les polluants comme source de carbone. Cette propriete est avantageuse pour la degradation de produits toxiques a de faibles concentrations. Les desavantages de 1'utilisation du champignon sont relies aux conditions optimales de culture. La temperature optimale de croissance du champignon est de 37-40 "C et Ie champignon requiert une source alternative de carbone et un apport maximal en oxygene. Puisque Ie systeme ligninolytique du champignon est active par la restriction au niveau de certains nutriments, la croissance du champignon et la degradation des polluants doivent etre des etapes separees et distinctes. Cette restriction peut compliquer Ie design et 1'operation d'unites de traitement en plus de poser certaines