Survie et croissance des boutures de Salix sous divers
régimes hydriques dans une perspective de
stabilisation des berges par génie végétal - Approche
expérimentale établie dans le but d'améliorer les
connaissances et les pratiques du génie végétal
Mémoire
Naren Keita
Maîtrise en biologie végétale - avec mémoire
Maître ès sciences (M. Sc.)
Survie et croissance des boutures de Salix sous
divers régimes hydriques dans une perspective de
stabilisation de berges par génie végétal
Approche expérimentale établie dans le but d’améliorer les
connaissances et les pratiques du génie végétal
Mémoire
NAREN KEITA
Sous la direction de :
Monique Poulin, directrice de recherche
André Evette, codirecteur de recherche
Résumé
Le dynamisme des cours d’eau peut avoir des effets dévastateurs sur les infrastructures à proximité et la stabilisation des berges. L’enrochement a longtemps été utilisé comme technique d’aménagement des berges, jusqu’à une période récente où le génie végétal a été reconnu comme une technique de replacement. Toutefois, les connaissances sont limitées concernant la réponse des espèces pionnières des zones riveraines soumises à différents régimes hydriques. Dans le cadre de la présente étude, la survie et la croissance végétative des boutures de saule (Salix) sous divers régimes hydriques ont été étudiées. Les résultats indiquent un taux de survie supérieur à 88% pour l’ensemble des trois espèces, peu importe le régime hydrique. Les trois espèces testées, soit S. discolor, S. eriocephala et S. interior, ont présenté une bonne tolérance au stress hydrique, bien que S. eriocephala ait produit la plus grande biomasse aérienne et racinaire des trois espèces. La présente étude jette ainsi les premières bases pour assurer une bonne utilisation de ces espèces dans les ouvrages à partir des techniques de génie végétal. Sur la base de la présente étude, l’usage de ces espèces et plus particulièrement du S. eriocephala pourrait contribuer à la réussite des projets de génie végétal pour la stabilisation des berges au Québec.
Table des matières
Résumé ... ii
Table des matières ... iii
Liste des tableaux ... v
Liste des figures ... vi
Remerciements ... vii Avant-propos ... ix Introduction : ... 1 CHAPTER 1 ... 9 1.1 Résumé ... 10 1.2 Introduction ... 11 1.3 Méthode ... 14 1.3.1 Design expérimental ... 14
1.3.2 Mise en place des traitements... 15
1.3.2.1 Récolte des boutures ... 15
1.3.2.2 Plantation des boutures ... 17
1.3.3 Application des traitements de régimes hydriques ... 17
1.3.3.1 Traitement en conditions à la capacité au champ ... 17
1.3.3.2 Traitement en conditions de sécheresse ... 18
1.3.3.3 Traitement en conditions d’inondation ... 18
1.3.4 Conditions de croissance ... 18
1.3.5 Suivi ... 19
1.3.5.1 Conditions hydriques du substrat de sol ... 19
1.3.6 Variables réponses correspondant à la croissance... 20
1.3.7 Analyse des données ... 21
1.4 Résultats ... 21
1.4.1 Survie des saules ... 21
1.4.2 Croissance des saules ... 21
1.4.2.1 Longueur racinaire ... 21
1.4.2.2 Production de biomasse ... 23
1.4.2.3 Volume racinaire ... 27
1.5 Discussion ... 29
1.5.1 Effet des régimes hydriques ... 29
1.6 Remerciements ... 33 Conclusions ... 34 Bibliographies ... 35
Liste des tableaux
Tableau 1 : Les techniques de génie végétal pour la stabilisation des berges. ... 6 Tableau 2 : Origine de 36 boutures de S. discolor, S. eriocephala et S. interior collectées. ... 16 Tableau 3 : Effet du régime hydrique et du type d’espèce sur la longueur racinaire totale. ... 23 Tableau 4 : Effet du régime hydrique sur les biomasses aériennes, racinaires, totales et ratio de
biomase sèche des trois espèces (S. discolor, S.eriocephala, S.interior) sous trois régimes
hydriques…. ... 26
Tableau 5 : Effet du régime hydrique sur le volume racinaire des trois espèces (S. discolor,
S.eriocephala, S.interior) sous trois régimes hydriques, soit en conditions de sécheresse, de capacité au champ et d’inondation ... 28
Liste des figures
Figure 1. A. Disposition des six sacs de sable dans chacune des parcelles principales correspondant
à un régime hydrique; B. Exemple d’un bac contenant six sacs (en sous-parcelles) avec chacun un échantillon de chacune des trois espèces de saules (total de six boutures par bac) ; C. Exemple d’un bloc avec les trois régimes hydriques. ... 15
Figure 2. Dispositif expérimental établi dans la serre haute performance . ... 19
Figure 3. Suivi de la condition hydrique du substrat de sol pour les trois traitements hydriques . 20
Figure 4. Longueur racinaire totale en fonction des régimes hydriques et en fonction des espèces
de Saules... 22
Figure 5. Biomasse aérienne totale (A), racinaire (B), aérienne/racinaire (C) et totale (D).. ... 25
Remerciements
Je remercie chaleureusement ma directrice de recherche Mme Monique Poulin, professeure et chercheure au Département de phytologie de la Faculté des Sciences de l’Agriculture et de l’Alimentation pour son encadrement émérite dans ma formation scientifique et au-delà du cadre professionnel. Son dynamisme et sa rigueur pour la recherche m’ont permis de développer des compétences remarquables tout au long de mon parcours à l’Université Laval. J’ai été très sensible à son sens d’humanisme et à l’originalité de ses initiatives. Par sa présence et son accompagnement, je n’ai jamais senti le poids de la distance avec les miens qui se trouvent à des milliers de kilomètres du Canada.
Je tiens aussi à remercier sincèrement mon codirecteur, M. André Evette . Je vous remercie beaucoup pour vos conseils, votre disponibilité, et votre support durant mon parcours de maîtrise au programme de biologie végétale.
Je voudrais remercier chaleureusement Maxime Tisserant pour son appui et sa contribution sans faille à la réussite de mes cours et de mon intégration à l’Université Laval. Mes remerciements vont aussi à Sandrine Hogue-Hugron, Léo Jeanne Paquin, Jean François Rioux et à Jérôme Cimon-Morin pour leurs précieux appuis et conseils.
Je remercie Dr Steeve Pépin professeur au Département des Sols et de génie agroalimentaire, Cintia Racine (étudiante au doctorat de Sylvio Demers) et Isabelle Clermont pour leur contribution à la réussite de mon expérimentation, notamment via leur aide dans l’apprentissage de la manipulation de l’appareil TDR (évalue la constance diélectrique du sol).
Je tiens aussi à remercier Audrey Auclair, Jennifer Brodeur, Pierre Mathieu Charest, Daniel Campeau, Marie-Michèle Thibaudeau et Hélène Servais, tous les membres du projet de formation pour la sécurité alimentaire au Mali pour leurs accompagnements irréprochables et l’Affaire Mondiale Canada de m’avoir offert la bourse d’études et pour m’avoir donnée l’opportunité de réaliser l’un de mes rêves le plus précieux, qui était de venir poursuivre mes études supérieures au CANADA.
Finalement, je voudrais exprimer toute ma reconnaissance à mes parents Seriba KEITA et Safiatou Doumbia, mon mari Ousmane Z TRAORÉ et ma famille à Québec Lansénou KEITA et Bintou CISSÉ qui m’ont apporté un soutien moral sans faille tout au long de cette aventure.
Avant-propos
Le présent mémoire se divise en deux principales sections. La première section représente l’introduction générale. Elle consistait à l’élaborer un cadre conceptuel en lien avec la stabilisation des berges par la technique de génie végétal et à traiter la capacité de bouturages de Salix soumis à différents régimes hydriques. La section a été rédigée sur la base des connaissances existantes dans la littérature en rapport avec le sujet de recherche. Pour cette section, exclusivement théorique, je me suis inspirée des expériences des études antérieures, afin de pouvoir identifier dans un premier temps, la problématique liée à la pratique du génie végétal, ensuite d’identifier les limites de la recherche sur le sujet. Ainsi, cela nous a permis de pouvoir développer la question de recherche, dont la réponse apporte sa part de contribution à certains problèmes rencontrés dans les projets de génie végétal. Pour cela, nous nous sommes inspirés des études faites en Europe, en Amérique du Nord, aux États-Unis et au Mexique. Monique Poulin (directrice de recherche), André Evette (codirecteur) et Maxime Tisserant (étudiant au doctorat dans le laboratoire de Monique Poulin) m’ont recommandé plusieurs articles et ouvrages qui traitent de mon sujet de recherche.
La deuxième section a été rédigée sous forme d’article. Je suis l’auteure principale. L’article présenté au chapitre 1 est intitulé « Survie et croissance des boutures de Salix sous divers régimes hydriques dans une perspective de stabilisation des berges par de génie végétal». Pour cet article, j’ai cherché les populations naturelles de saules et effectué la collecte des boutures sur le terrain à l’automne 2017. Aussi, j’ai élaboré et mis en place le dispositif expérimental et procédé à la prise de données durant l’expérimentation. J’ai effectué les analyses statistiques et rédigé le manuscrit. Les deux coauteurs, Monique Poulin et André Evette ont contribué à l’élaboration du protocole de recherche, au choix des espèces, à l’élaboration du dispositif expérimental, au suivi de l’expérience et à la révision du manuscrit. Aussi Maxime Tisserant a pleinement contribué à toutes les étapes de la présente étude. Enfin, Sandrine Hogue-Hugron (professionnelle de recherche) a contribué à la révision du manuscrit.
Introduction :
Cadre conceptuel de la stabilisation des berges par la
technique de génie végétal
Problématique
Les cours d’eau sont des milieux très complexes et se distinguent par un état d’équilibre non statique (Choné, 2013). Cela indique que les cours d’eau sont des milieux très dynamiques et peuvent occuper dans le temps différentes zones en migrant dans leur plaine d’inondation (Marcoux-Viel, 2015). Cette variabilité spatiale s’accompagne d’une variabilité temporelle des débits, notamment au cours du cycle hydrologique annuel. Par exemple, le chenal d’écoulement d’eau perceptible en période estivale disparait complètement au printemps à cause des effets de la crue causée par la fonte des glaces et des neiges. La dynamique naturelle des cours d’eau, bien qu’elle soit souhaitable pour la diversité des habitats riverains et la recharge solide, peut représenter un risque pour la sécurité de la population localisée à proximité. C’est pourquoi la prise en compte des milieux riverains et de tout l’espace de liberté des rivières est capitale (Gagnon, 2007). Elle permet d’assurer la résilience des berges des cours d’eau et d’amoindrir les risques sur les infrastructures (Biron et al., 2013).
Par ailleurs, les populations humaines vivant près des cours d’eau ne cessent d’augmenter. Les raisons de cette densité près des cours d’eau sont assez nombreuses : la présence de la ressource en eau, les services écosystémiques offerts par les milieux riverains et l’étalement urbain (Klein, 1979). Ce dernier favorise une augmentation des surfaces imperméables par le développement des infrastructures routières et des constructions immobilières. La localisation des routes et des infrastructures à proximité des rivières amène des besoins en stabilisation de berges afin d’assurer la sécurité publique.
Le génie civil a longtemps été admis comme la méthode traditionnelle d’aménagement des berges des cours d’eau, afin d’éviter leur érosion et assurer leur stabilisation. Le génie civil se base sur l’utilisation de matériaux inertes (l'enrochement de pierre, chaussée en béton, gabions de roche, cloisons en acier, béton ou aluminium) comme moyens de protection des berges contre les phénomènes érosifs des sols dans les milieux riverains. L’avantage des techniques de génie civil réside notamment dans leur forte potentialité de résistance mécanique et leur capacité à stabiliser les berges, juste après l’établissement de l’ouvrage (Li & Eddleman,
2002). Au Québec, la technique de génie civil est utilisée très fréquemment et son application est de plus en plus critiquée puisque les ouvrages de génie civil créent un effet de structure artificielle le long des cours d’eau (Paquette, 2010), menant à une dénaturalisation du paysage qui se trouve appauvri en végétation et en fonctions écologiques (Cavaille et al., 2013). Selon la Société de la faune et des parcs du Québec (2003), le recours aux techniques de génie civil n’est valable uniquement que lorsque le contexte l’impose. Il est maintenant reconnu que le recours aux matières végétales constitue une solution de remplacement socio-économique et écologiquement viable (Rey et al., 2004).
Définition du génie végétal et historique
Le génie végétal est l’une des composantes du génie biologique (Labonne et al., 2007). Il se caractérise par les techniques utilisant les capacités morphologiques et fonctionnelles de la couverture végétale, pour des fins de protection et conservation des berges des milieux riverains (Rey, 2004). Il s’agit notamment des parties aériennes et souterraines de certaines espèces riveraines, qui maintiennent le sol face aux contraintes mécaniques érosives exercées par l’eau. La pratique du génie végétal existe depuis des temps immémoriaux. Depuis des siècles, les ouvrages de génie ont été réalisés en Chine et en Rome antique (Frossard & Evette, 2009). À cette époque, le recours aux techniques de génie végétal reposait essentiellement sur les nécessités économiques et sociales auxquelles faisaient face les populations établies près des rives. Par exemple en France, pendant très longtemps, les populations se servaient des techniques de génie végétal afin de résoudre des problématiques d’érosions torrentielles pouvant menacer la sécurité de la population humaine riveraine. Aussi, elles permettaient de renforcer la stabilité des infrastructures (routes, maison) contre les problèmes d’érosion (Labonne et al., 2007).
Aujourd’hui, le génie végétal est utilisé dans une perspective de vision encore plus large. En effet, l’une des raisons principales qui justifient son usage concerne les services écologiques offerts par les végétaux (Kuzovkina & Volk, 2009). Il existe dans la littérature plusieurs références, qui ont évoqué les avantages offerts par le génie végétal. Notamment, sur le plan écologique, les végétaux servent de filtre et permettent de prévenir la pollution diffuse en provenance des surfaces agricoles situées à proximité des cours d’eau. Ainsi, ils contribuent à la purification de la qualité de l’eau. De plus, les végétaux riverains participent à l’augmentation et au maintien de la biodiversité. Aussi, ils jouent des fonctions de corridor écologique et alimentent en énergie les petits cours d’eau . Enfin sur le plan social, les paysages végétalisés des milieux riverains offrent un espace de loisirs (marche, ornithologie) au service des personnes vivant à proximité des berges (Dufour & González, 2019). Les végétaux utilisés dans les ouvrages de génie végétal maintiennent les sols contre les différentes formes de dégradation (l’action des glaces, la pression du courant d’eau, etc.). Ils agissent à deux niveaux pour contrer les phénomènes d’érosion. D’une part, ils empêchent l’ablation du substrat en retenant le sol, d’autre part, ils diminuent la vitesse de l’eau en augmentant la
rugosité et favorisent la sédimentation, en retenant les sédiments érodés plus en amont (Rey et al, 2004). Selon une étude comparative entre des surfaces diversement végétalisées, le niveau de l’érosion devient très élevé dans les zones presque dénudées (partiellement végétalisées) par rapport aux zones végétalisées (Reid et al., 1999). Autrement dit, le niveau de l’érosion décroît en fonction de l’augmentation de la couverture végétale (Battany & Grismer, 2000). Le génie végétal est ainsi reconnu comme une technique dont la fonction de stabilisation des berges augmente avec le temps suite au développement de la végétation.
Les techniques du génie végétal
Selon la problématique, différents types de stabilisation sont envisageables pour prévenir l’érosion et protéger les infrastructures situées à proximité de la berge (Labonne et al., 2007). Chaque type de stabilisation peut faire appel à plusieurs techniques de génie végétal (Tableau 1). Les différentes techniques de génie végétal présentées ont été sélectionnées en fonction de leur résistance face aux contraintes hydriques et de leurs fonctions de protection contre l’érosion (Tableau 1). Les précisions ou descriptions font référence à des données de guides pratiques sur les techniques de génie végétal dans le Tableau
Tableau 1 Les techniques de génie végétal pour la stabilisation des berges.
Type de stabilisation Ouvrages de génie végétal Fonction de protection
Stabilisation du fond du lit
Les Garnissages des branches Ralentissent l’écoulement de l’eau, captent les sédiments et évitent leur transport par érosion hydrique.
Les barrages en caissons végétalisés
Prévient l’érosion post installation de l’ouvrage en bloquant le fond du lit et les berges du cours d’eau.
Les barrages en fascinages Prévention contre le courant du fond du cours d’eau
Les barrages en clayonnages Prévention contre le courant du fond du cours d’eau
Stabilisation des berges
Les plantations Empêche l’érosion des berges par les parties aériennes et racinaires des plantes sollicitées par l’érosion hydrique
Le bouturage
Utilisé pour protéger les rivages des cours d’eau à travers une bouture ligneuse d’une espèce pionnière des milieux riverains et ayant une forte capacité de production de biomasse racinaire et aérienne
Lit de plants et plançons Utilisé pour les stabilisations rapides avec une pénétration des racines très profondes
Tressage Protège les pieds de berges à faible hauteur soit maximum 40cm. Il donne un effet de mur végétal après installation.
Fascine Généralement accompagnée d’autres techniques de protection, elle est adaptée à la protection des niches d’érosion tout au long du cours d’eau Peigne Empêche les sapements de berge et les arrachements de niches, elle a un effet
immédiat de stabilisation Les techniques combinées ou
techniques mixtes
Les végétaux et les pierres combinés ensemble agissent comme des supports pour le substrat de sol
Canalisation de l’eau
Les épis en fascines
Les saucissons (fascine de gros diamètres associant terre, pierre et branches de saules)
Permettent d’orienter la circulation des cours d’eau vers un écoulement rectiligne. Il réduit aussi les méandres du cours d’eau. Ces techniques qui conduisent à l’artificialisation des milieux sont aujourd’hui en désuétude
Génie végétal et le génie civil
Le génie végétal comme le génie civil sont deux approches de stabilisation des berges avec des particularités et spécificités bien différentes. Le choix entre les deux approches dépend fortement de la condition écologique du milieu à stabiliser (Frossard & Evette, 2009). Par exemple, pour certains profils de cours d’eau avec des contraintes mécaniques très élevées, l’usage du génie végétal pour la stabilisation des berges sera difficile, voire impossible. Alors, le recours à l’approche par le génie civil peut être recommandé. Le génie civil permet donc de stabiliser les berges lorsque les méthodes de génie végétal sont inappropriées.
Les méthodes du génie civil visent une stabilisation immédiate, c’est-à-dire dès l’établissement de l’ouvrage (Li & Eddleman, 2002). Cela est important dans les situations où les besoins de stabilisation sont urgents (assurance immédiate de la sécurité des riverains face à la crue). Quant au génie végétal, la fonction de stabilisation n’est pas immédiate et dépend très fortement de la conservation du matériel végétal utilisé et de sa capacité de développement végétatif dans le temps (production des racines et des feuilles). Mais toutefois, la pratique du génie végétal présente plusieurs avantages. Ces avantages sont d’ordre écologique (améliorer la biodiversité, les fonctions de corridor, la dépollution, la fertilité du sol), physiologique (production de l’oxygène pendant la photosynthèse, absorption de l’eau par les racines) et physique (effet d’ancrage du système racinaire, et frein à l’écoulement plus effet tapis des parties aériennes) (Frossard & Evette, 2009). Aussi, les solutions offertes par les techniques du génie végétal voient leur résistance mécanique augmenter avec le temps. Cette augmentation graduelle de la résistance fait que les végétaux arrivent à préserver les berges dans un état proche du naturel, tout en bloquant l’érosion et en captant les sédiments (Ministère du Développement, de l’Environnement et des Parcs, 2005). Contrairement au génie végétal pour lequel les fonctions de stabilisation sont
avantages multiples souvent supérieurs au génie civil à savoir la stabilisation durable des berges, la consolidation des substrats de sol par les racines, la captation des sédiments (Lee et al., 1998), les effets positifs sur la biodiversité et les fonctions écologiques associées contribuant au caractère naturel des sites.
Parmi les facteurs qui limitent le développement du génie végétal, nous trouvons : l’intensité de la pente (trop escarpée), du débit ou des vagues (Rey et al., 2004). A cela peut s’ajouter aussi la variation des régimes hydriques des cours d’eau, qui représente un enjeu majeur pour l’application des techniques de génie végétal (Markus-Michalczyk et al., 2016). Ces contraintes hydriques peuvent se manifester par des sècheresses estivales ou par des inondations printanières, ces deux processus ont des influences très marquées sur la survie et la croissance végétative des espèces végétales utilisées à des fins de stabilisation (Doffo et al., 2017). La tolérance à la sécheresse est essentielle pour la survie des espèces utilisées durant la période estivale, surtout pendant les premières périodes de végétation. De même, les inondations printanières peuvent noyer les plantes pendant le début de leur cycle de végétation. Le rôle crucial des processus de sécheresse et d’inondation sur la réussite des ouvrages de génie végétal souligne ainsi la pertinence de travailler au développement des connaissances des propriétés biologiques, physiologiques et mécaniques des espèces riveraines face aux variations de stress hydrique (Kuzovkina & Volk, 2009).
Objectif et hypothèse de recherche
La présente étude a pour objectif d’enrichir les connaissances sur la survie et la croissance des boutures de ces trois espèces soumises à différents stress hydriques, dans une perspective d’application au domaine du génie végétal. J’ai ainsi émis l’hypothèse que la tolérance au stress hydrique d’inondation et de sécheresse entre les espèces différera en fonction de leur répartition dans le gradient de pente en milieu naturel. Ainsi, je prédis que S. eriocephala, qui se trouve sur tout le gradient de pente en milieu naturel, aura une plus grande tolérance à tous les régimes hydriques alors que le régime d’inondation sera favorable au S. interior et le régime de sécheresse au S. discolor, qui poussent respectivement en bas et en haut de berge.
CHAPTER 1
Survie et croissance des boutures de Salix sous divers régimes
hydriques dans une perspective de stabilisation des berges par
1.1 Résumé
Les zones riveraines sont soumises à une large variation de régimes hydriques. Ce changement de régime hydrique peut être un facteur limitant l’établissement des boutures de certaines espèces de Salix en condition d’inondation (en période de crue) ou de sécheresse (en période d’étiage) dans les ouvrages de génie végétal. Or, les connaissances sur la résistance au stress hydrique des espèces de Salix utilisées dans les ouvrages de génie végétal sont limitées. Afin d’enrichir les connaissances sur la capacité de reprise et de développement végétatif de trois espèces de Salix largement répandues en milieu riverain au Québec et très utilisées en génie végétal, nous avons établi un dispositif expérimental en tiroirs en serre à l’Université Laval comportant trois traitements hydriques (i) capacité au champ (pendant toute l’expérience à la suite de la période d’acclimatation), (ii) inondation (submersion totale pendant deux semaines suivie d’une semaine à la capacité au champ) répétée quatre fois (iii) sécheresse ( arrosage une fois par semaine). Des mesures de longueur racinaire, de biomasse aérienne et racinaire ont été prises après 3 mois. Les trois espèces ont présenté une forte capacité de reprise (supérieur à 88 % de survie). Il n’y avait pas d’effet du stress hydrique sur la production de biomasse racinaire pour les trois espèces, mais pour S. interior la longueur racinaire a été favorisée en conditions d’inondation par rapport aux deux autres conditions. S. eriocephala a produit une biomasse totale 3,14 fois plus élevée que ne l’a fait S. discolor et 1,85 fois plus grande que S. interior. Les trois espèces ont présenté une certaine tolérance aux deux types de stress. Néanmoins, entre les trois espèces, S. eriocephala a présenté une plus grande tolérance au stress hydrique puisqu’il a produit en général la plus grande racinaires ainsi que les plus fortes biomasses.
1.2 Introduction
Plusieurs espèces de la famille des Salicacées sont des plantes pionnières associées aux écosystèmes riverains (Doffo et al., 2017). Elles sont adaptées au régime de perturbation par les crues, notamment grâce à leur capacité à produire de nombreuses graines voyageant par le vent et sur l’eau, leur conférant un bon potentiel d’établissement sur les rives en absence de compétition (Karrenberg et al., 2002). Ces mêmes espèces sont de plus en plus utilisées dans les projets de génie végétal pour la stabilisation des berges de cours d’eau grâce à leur grande reprise après bouturage, particulièrement pour le genre Salix (Bergeron & Roy, 1985; Francis et al., 2005). Elles sont ainsi utilisées dans plusieurs techniques de génie végétal, soit en tressage ou en fascines ainsi que dans des matelas de branches afin de protéger la berge contre l’érosion, assurant ainsi la stabilité des infrastructures adjacentes ( Michalczyk et al., 2016). Plusieurs raisons permettent de justifier l’utilisation des espèces de Salix dans les projets de génie végétal. Entre autres, certaines espèces sont pourvues d’un réseau racinaire allant jusqu’à 1,3 m de profondeur (Kopp et al., 2001) et elles peuvent ainsi stabiliser le sol en profondeur (Dimitriou & Aronsson, 2005). Les saules disposent aussi d’une grande capacité de production végétative assurant un établissement rapide (Plante, 2012).
Le choix des espèces de Salix à utiliser dans les ouvrages de génie végétal doit tenir compte de la tolérance de chaque espèce aux conditions très variables dans le temps et au sein du gradient de pente dans le talus riverain. La survie et la croissance des boutures de saules peuvent en effet être affectées par l’alternance de périodes de crue et d’étiage et les espèces sélectionnées doivent pouvoir résister aux forces tractrices de l’eau et aux conditions anoxiques en période de crue, mais également pouvoir tolérer des conditions de sécheresse en période d’étiage. Ces régimes de perturbations (arrachement de la biomasse) et de stress (anoxie et sécheresse) seront différents entre le bas et le haut de talus. Le choix des espèces
Quelques études réalisées en Europe et aux États-Unis et portant sur la stabilisation des berges par la technique de génie végétal ont porté sur l’effet des conditions hydriques sur la survie et les caractéristiques morphologiques des boutures de certaines espèces de peupliers et de Salix (Castagni, 2017; Lavaine, 2013; Parent et al., 2008). Ces études ont permis d’identifier quatre espèces de saules, S. alba, S. elaeagnos, S. triandra pour l’Europe et S. nigra pour les États-Unis, qui possèdent des caractéristiques d’intérêt pour le génie végétal, notamment leur haut potentiel de propagation végétative. Au Québec, bien que le génie végétal soit en essor depuis quelques décennies, peu de travaux ont porté sur l’identification d’espèces adaptées aux variations du régime hydrique et sur les caractéristiques morphologiques et anatomiques des espèces de saules dans les ouvrages de génie végétal. Les travaux expérimentaux de Castagni (2017) ont traité de trois espèces largement répandues en milieux naturels au Québec, soit S. eriocephala, S. interior et Cornus stolonifera. Toutefois, d’autres espèces de Salix non considérées dans ces travaux colonisent naturellement des zones riveraines des cours d’eau au Québec et pourraient être des espèces potentielles à exploiter dans les ouvrages de génie végétal pour la stabilisation des berges.
L’effet du stress hydrique (sécheresse et inondation) sur la croissance et la survie des espèces de Salix a été abordé dans plusieurs études (Imada et al., 2008; Li et al., 2004). Par exemple, le manque de tolérance de certaines espèces de saules au stress de sécheresse serait dû à leur taux de transpiration élevé et à la faible capacité du xylème à remplir ses fonctions à la suite d’une réduction du contenu en eau du sol (Wikberg & Ogren, 2004). Aussi, la production de biomasse aérienne est directement reliée à l’activité du potentiel de conductance stomatique. Or, l’effet du stress hydrique de sécheresse induit une augmentation de l’évapotranspiration, qui peut affecter négativement ce potentiel de conductance stomatique (Waldren et al,1987) et réduire par conséquent la biomasse produite. De plus, la durée du stress de sécheresse peut ralentir la croissance et compromettre la survie des individus de saule (Nakai & Kisanuki, 2011). Selon Nakai & Kisanuki (2011), au-delà de trois semaines consécutives en condition de sécheresse, la conductance stomatique est deux fois plus faible que celle en condition de capacité au champ. En revanche, le manque de tolérance des saules aux conditions d’inondation serait associé à la réduction du taux d’oxygène dans le sol qui crée des
conditions anaérobies compromettant la croissance des racines (Francis et al, 2005). Une inondation au-delà de 28 jours provoque une réduction considérable du potentiel d’oxydoréduction (Jackson & Attwood, 1996), qui se caractérise par une diminution de la quantité d’oxygène dans le sol. Un ralentissement de la croissance est possible après une période de 20 jours d’inondation (Imada et al., 2008; Imada et al., 2010).
Certaines espèces ont développé des mécanismes d’adaptation leur permettant de survivre en conditions de stress hydrique, en attendant le retour de conditions adéquates. Par exemple, un abaissement du niveau d’eau du sol peut induire le développement progressif des racines en profondeur dans le sol pour résister à la sècheresse (Pezeshki et al, 1998 ; Wikberg & Ogren, 2004). En revanche, face au stress d’inondation (Li et al, 2004; Sennerby-forsse & Zsuffa, 1995) certaines espèces tolèrent les conditions anaérobies grâce au développement de lenticelles hypertrophiées (pores favorisant les échanges d’oxygène) en surface de la zone racinaire et à la formation de nouvelles racines adventives (Nakai & Kisanuki, 2011). Cela implique que certaines espèces de Salix disposent des caractéristiques physiologiques d’intérêt qui pourraient être exploitées en génie végétal en plus des caractéristiques morphologiques.
Pour la présente étude, nous avons ciblé trois espèces de saules très répandues en Amérique du Nord et plus particulièrement dans les milieux riverains au Québec: S. discolor, S. eriocephala et le S. interior (Argus, 2010). Le choix de ces espèces repose, d’une part, sur leurs caractéristiques morphologiques qui en font des candidates intéressantes pour le génie végétal. En effet, les trois espèces produisent de la biomasse en grande quantité (Kuzovkina & Volk, 2009) et possèdent une capacité de reprise élevée à partir d’une bouture (supérieur à 84%; Castagni, 2017). D’autre part, ces trois espèces ont une répartition naturelle différente dans le talus riverain. En effet, chacune des trois espèces occupe une position distincte qui reflète probablement leur capacité de tolérance aux conditions de stress hydrique (anoxie et sécheresse (Pezeshki et al, 2007). Ainsi, une espèce comme le S. discolor qui est présente en
la réponse de ces trois espèces de saules à trois régimes hydriques différents, soit de sécheresse, de capacité au champ et d’inondation, dans des conditions contrôlées en serre.
1.3 Méthode
1.3.1 Design expérimental
Nous avons évalué la survie et la croissance de trois espèces de saules, soit S.discolor, S.eriocephala, S.interior, à partir de boutures soumises à trois régimes hydriques. Un dispositif expérimental en tiroirs a été mis en place dans le complexe de serre de haute performance de l’Université Laval à l’hiver 2018, pour une durée de trois mois (05 février au 05 mai). Trois régimes hydriques ont été testés en parcelles principales soit la sécheresse, la capacité au champ et l’inondation, alors que les trois espèces de saules ont été établies en sous-parcelles. Chaque régime hydrique était appliqué à un bac (110 cm longueur x 72 cm de largeur x 100 cm (profondeur) contenant six sacs de sable de 30 L (35,6 cm x 29,8 cm) dans lesquels une bouture a été plantée, pour un total de deux boutures de chacune des trois espèces par bac. Les bacs étaient munis d’un système de drainage qui permettait de contrôler le niveau d’eau. Les bacs ont été disposés en blocs de façon à tenir compte des variations de température et d’humidité dues à la présence du chauffage et de l’orientation du brumisateur dans la serre (Figure 1 et 2).
Figure 1. A. Disposition des six sacs de sable dans chacune des parcelles principales correspondant à un régime hydrique; B. Exemple d’un bac contenant six sacs (en sous-parcelles) avec chacun un échantillon de chacune des trois espèces de saules (total de six boutures par bac) ; C. Exemple d’un bloc avec les trois régimes hydriques.
1.3.2 Mise en place des traitements
1.3.2.1 Récolte des boutures
Les boutures des trois espèces de saules ont été collectées dans la région de la Capitale nationale en novembre 2017, moment où les plantes étaient en dormance et dépourvues de feuilles. Les boutures ont été récoltées dans des populations naturelles en milieux riverains. Quatre populations avaient été préalablement sélectionnées au cours de l’été en fonction de la présence d’au moins une espèce, parfois jusqu’à la totalité des trois
A B
que pour les rameaux plus âgés. Pour chacune des trois espèces, 36 boutures ont été récoltées (12 plants mères x 3 boutures), pour un total de 108 boutures. Dans le but d’assurer la survie optimale des boutures, elles ont été immédiatement placées dans des sacs sous vide et gardées au froid à -4°C jusqu’au début de l’expérience (période d’entreposage de 2 mois).
Tableau 2 Origine de 36 boutures de S. discolor, S. eriocephala et S. interior collectées.
Espèces Populations Nbre
Boutures/ pied mère
S. discolor
Plein air Sainte Foy Rivière Cap Rouge Pointe St-Vallier Trait Carré 12/4 6 /2 12 /4 6/2 S. eriocephala
Plein air Sainte Foy Rivière Cap Rouge Parc chaudière Trait carré 15/5 6/2 9/3 6/2 S. interior
Plein air Sainte Foy Parc du Berger Parc chaudière Rivière St-Charles 3/9 3/1 6/2 18/6
1.3.2.2 Plantation des boutures
Avant l’instauration de l’expérimentation en serre, les boutures ont été submergées dans l’eau 24 heures afin d’assurer leur hydratation et d’optimiser l’émergence des parties aériennes et racinaires (Edwards et al., 1976; Pezeshki et al., 2007). Le diamètre de chaque bouture a été mesuré à l’aide d’un vernier électronique. Ensuite, le 5 février 2019, elles ont été transplantées dans un substrat de sable (densité apparente : 1,47 g; densité réelle : 2,66 g) contenu dans des sacs de tissus perméables à l’eau. Les deux tiers de la longueur de chaque bouture occupaient la partie souterraine et le reste consistait en la partie hors sol, qui comportait au moins trois bourgeons. Dans le but de maximiser la survie, les boutures ont été soumises à des conditions optimales de croissance (correspondant au régime hydrique de capacité au champ) durant une période d’acclimatation d’un mois. À la suite de l’expression de quelques symptômes de carence observés un mois après installation, les boutures ont été fertilisées avec 500 ml de NPK (20-20-20) liquide (pour chaque volume de pot correspondant à 50 ml). Les doses étaient faibles, afin de ne pas stimuler la croissance des boutures, mais simplement d’assurer leur établissement.
1.3.3 Application des traitements de régimes hydriques
À la suite de la période d’acclimatation d’un mois, les boutures des espèces du S. discolor, S. eriocephala et du S. interior ont été soumises aux trois régimes hydriques. Les traitements hydriques appliqués étaient inspirés de Castagni (2017) et Lavaine (2013).
1.3.3.1 Traitement en conditions à la capacité au champ
Le volume d’eau requis pour l’irrigation des boutures à la capacité au champ a été déterminé grâce aux notions agronomiques. La quantité à la capacité au champ correspond au volume d’eau dans le sol après 48 heures à la suite de la submersion totale du substrat de sol (Fortin, 2016). Concrètement, trois sacs de sable sec sans bouture ont été placés en submersion totale
quantité d’eau ajoutée était fonction du besoin en eau à la capacité au champ, soit un niveau d’eau qui variait entre 3 à 4 litres correspondant au 30% de volume d’eau du sac de sable.
1.3.3.2 Traitement en conditions de sécheresse
Pour le traitement en conditions de sécheresse, les substrats étaient soumis à un contenu en eau du sol très faible, en deçà du seuil optimal pour la croissance des plantes, mais au-dessus du seuil du point de flétrissement (la réserve en eau du sol qui n’est pas utilisable par la plante). Pour ce traitement les boutures étaient irriguées une seule fois par semaine avec un volume d’eau de 1 à 2 litres par irrigation soit 15 % du volume d’eau du sac de sable (Castagni, 2017).
1.3.3.3 Traitement en conditions d’inondation
Pour le traitement en conditions d’inondation, le niveau d’eau dans le sol était maintenu à 5 cm au-dessus de la surface du substrat durant une période de deux semaines. L’eau du bac était ensuite évacuée et les boutures soumises aux conditions de capacité au champ pendant une semaine. Ce cycle fut répété quatre fois durant la période d’expérimentation soit un intervalle de 7 jours.
1.3.4 Conditions de croissance
Pendant toute la période de l’expérimentation, la température de l’air de la serre était toujours supérieure à 17,0 °C avec un maximum de jour de 28,0 °C. La moyenne était de 23,5 °C. L’humidité relative dans la serre était supérieure à 35%, avec une humidité maximum de 75% le jour et de 78% la nuit soit une moyenne hebdomadaire de 57,8%. Enfin, le rayonnement photosynthétique actif (Photosynthèses actives Radiation; PAR) de jour étaient supérieur à 6 mol/m2 avec un maximum était de 45 mol/m2et, pour une moyenne totale de 22 mol/m2.
Figure 2. Dispositif expérimental établi dans la serre haute performance de l’Université
LAVAL, 2575 rue Marie-Fitzbach, Québec.
1.3.5 Suivi
1.3.5.1 Conditions hydriques du substrat de sol
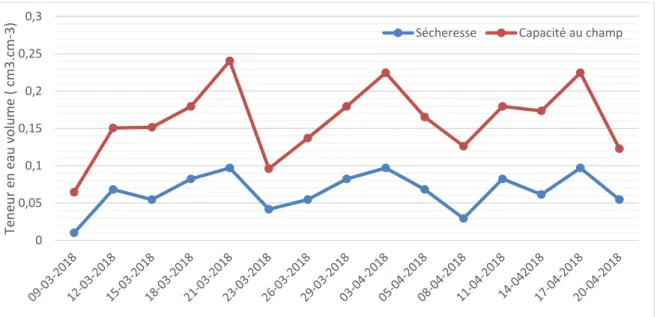
La teneur en eau volumique du substrat de sol était suivie tous les deux jours, dans le but de maintenir les niveaux d’eau correspondant à la capacité au champ et à la sécheresse. En effet, la masse sèche du substrat de sol initialement établie au début de l’expérience, le volume du sol et la teneur en eau du sol, ont servi de paramètres de base, afin de pouvoir vérifier la teneur en eau volumique du sol à l’instant T. La teneur en eau du sol dans les trois régimes hydriques a été déterminée grâce à un TDR (“Time Domain Reflectometry” / Réflectomètre temporel), muni de trois sondes de 15 cm. Les sondes émettent des ondes électromagnétiques faisant des mouvements circulaires (entre le substrat de sol et l’appareil)
puisque le sol était saturé en eau. Le but de ce dernier traitement était justement de mettre les boutures dans une condition de submersion totale pendant une semaine suivie d’une période de drainage. Durant la durée de l’expérience, la teneur en eau moyenne à la capacité au champ était de 10% alors que celle en conditions de sécheresse était de 6%, ces teneurs pouvant varier au cours de la période de l’expérimentation (Figure 3). Toutefois, compte tenu de la condition climatique de la serre, il était difficile d’atteindre le point de flétrissement temporaire (manque d’eau dans le sol) en tension.
Figure 3. Suivi de la condition hydrique du substrat de sol pour les trois hydrique ( traitements capacité au champ et à la sécheresse).
1.3.6 Variables réponses correspondant à la croissance
Aux termes de l’expérimentation, les boutures ont été soigneusement retirées du substrat et séparées en parties aérienne (feuilles et tiges) et souterraine (racines). Les différentes parties ont été séchées dans une étuve à 105 °C jusqu’à poids constant (48 heures). La biomasse aérienne et racinaire ainsi que la biomasse totale ont par la suite été déterminées avec une balance de précision. Le ratio du poids sec des feuilles versus celui des racines a aussi été calculé. Enfin les données racinaires (longueur racinaire totale et le volume racinaire) ont été déterminées à l’aide d’un scanner WinRhizo. Dans le but d’effectuer le balayage (scan), les racines de chaque bouture ont été préalablement nettoyées avec de l’eau, détachées des
0 0,05 0,1 0,15 0,2 0,25 0,3 Te n eu r en eau v o lu m e ( cm 3.cm
parties aériennes puis trempées dans une solution à colorant rouge pour une période de deux heures. Ensuite les racines (par bouture) ont été déposées sur le plateau pour les balayer.
1.3.7 Analyse des données
Nous avons testé l’effet des traitements sur la survie des boutures et les diverses mesures de croissance à l’aide d’une analyse de variance (ANOVA), dans laquelle, dans laquelle les régimes hydriques initialement mesurés en début d’expérience ont été considérés comme variables explicatives et la biomasse initiale des bouture, et la longueur des boutures (20 cm) en covariable. Les différences significatives étaient considérées à P < 0,05. L’homogénéité et la normalité de la variance ont aussi été vérifiées. Le logiciel d’analyse statistique SAS (version 9.4) a été utilisé pour l’analyse des variables réponses suivi d’un test de comparaisons multiples LSD a posteriori (Fisher's least significant différence).
1.4 Résultats
1.4.1 Survie des saules
Les boutures ont montré un établissement efficace, avec un taux de survie très élevé (supérieur à 88%) pour l’ensemble des trois espèces, quel que soit le régime hydrique considéré. En effet, toutes les boutures de S. eriocephala se sont établies, alors que celles de S. discolor ont montré un taux de survie variant de 88% (en conditions de sècheresse) à 94% (inondation) et celles de S. interior un taux entre 97% (sécheresse et capacité au champ) et 100% (inondation).
1.4.2 Croissance des saules
1.4.2.1 Longueur racinaire
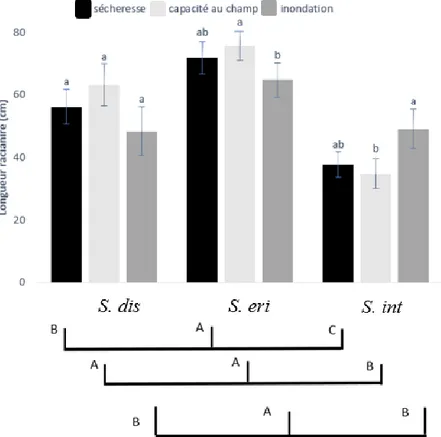
L’influence du régime hydrique sur la croissance en longueur des racines variait entre les espèces (interaction espèces*traitements significatifs; Tableau 3, Figure 4). Généralement la
racinaire plus faible par rapport à celle obtenue en condition de capacité au champ alors que les résultats inverses ont été trouvés pour S. interior (Figure 4). La longueur racinaire de S. discolor ne variait pas entre les régimes hydriques.
Figure 4. Longueur racinaire totale en fonction des régimes hydriques et en fonction des espèces. Les barres indiquent l’erreur type de la moyenne, les différentes lettres indiquent une différence significative à α = 0,05 entre les espèces (lettres minuscules) et entre les traitements (lettres majuscules), selon le test de comparaison multiple LSD.
Tableau 3 Effet du régime hydrique et du type d’espèce sur la longueur racinaire totale. Les espèces testées étaient S. discolor, S. eriocephala et le S. interior sous trois régimes hydriques, soit en conditions de sécheresse, de capacité au champ et d’inondation.
Sources
de variation
Degré de liberté ? Longueur des racines
F P Bloc 5 - Régimes hydriques Erreur A 2 10 0,51 0,61 Espèces 2 52,82 0,001*
Espèces* Régimes hydriques 4 3,32 0,04*
Erreur B 30 - Total (UE) Erreur échantillonnage Total échantillonnage 53 54 107 - - -
Le signe « * » indique une différence statistique à α = 0,05.
UE = Unité expérimentale
1.4.2.2 Production de biomasse
Les différents régimes hydriques n’ont pas eu d’impact sur la production de biomasse, que ce soit la biomasse aérienne, racinaire ou totale. Toutefois, la biomasse variait entre les espèces, pour toutes les parties de la plante (Tableau 4). Le S. discolor est l’espèce ayant produit le moins de biomasses aériennes alors que le S. eriocephala a produit une biomasse aérienne la plus forte, soit 3,5 et 1,6 fois plus élevée respectivement que celle de S. discolor et S. interior (figure 5.A). D’autre part, la plus faible biomasse racinaire a été produite par
biomasse du S. discolor était quasiment équivalente pour les feuilles et les racines, résultant en un ratio respectivement 2 fois et 3 fois inférieur à celui du S. interior et S. eriocephala (Figure 5.C).
Par ailleurs, la biomasse des feuilles et des tiges, qui constitue la biomasse aérienne, était presque identique chez le S. eriocephala (respectivement 55% et 45 %), comparativement aux S. interior et S. discolor pour lesquels la biomasse des feuilles était 2 fois supérieure à celle de la tige (Figure 5.A).
Figure 5. Biomasse aérienne totale (A), racinaire (B), aérienne/racinaire (C) et totale (D). Les barres représentent la moyenne de biomasse pour chaque espèce de saule ± l’erreur type. Les lettres minuscules indiquent les différences significatives à α = 0,05.
Tableau 4 Effet du régime hydrique sur les biomasses aériennes, racinaires, totales et ratio de la biomasse sèche des trois espèces ( S. discolor, S. eriocephala et S. interior) sous trois régimes hydriques, soit en conditions de sécheresse, de capacité au champ et d’inondation.
Sources Variation Degrés de liberté Feuilles F P Tiges F P Aérienne F P Racinaire F P Totale F P Ratio F P Bloc 5 - - - - - R.h. 2 Erreur A 10 Espèces 2 1,75 0,22 - 18,67 0,0027* 0,30 0,86 0,28 0,75 - 36,62 <,0001* 0,60 0,66 0,52 0,60 -25,80 0,001* 0,23 0,91 0,59 0,57 - 20,32 <.0001* 1,25 0,32 31,32 0,27 - 31,32 0,001* 0,57 0,68 0,05 0,95 - 39,12 0,001* Erreur B 30 - - - - Total (UE) 53 Erreur échant 54 Total échéant 107 - - - - - Le signe « * » indique la source de différence statistique à α = 0.05. Avec le test de Fisher et un LSD means a posteriori pour identifier la différence significative entre les moyennes. (ns : Non significatif. R.h. : régimes hydriques, B : Biomasse et échant : Échantillonnage. Espèces *R.h 4
0,44 0,77
1.4.2.3 Volume racinaire
Pour tous les diamètres racinaires confondus, le volume racinaire du S. eriocephala était près de deux fois supérieur à celui du S. discolor et S. interior (Figure 6.A). Le volume racinaire des trois espèces étudiées n’a pas été influencé par la variation des régimes hydriques (Tableau 5). Toutefois, il variait entre les espèces et en fonction du diamètre des racines (racines fines, moyennes, grosses). Par ailleurs, peu importe l’espèce, les racines fines (1,46 cm³) ont produit la plus grande partie du volume racinaire (67%), suivi par les racines de diamètre moyen (25%), alors que la plus faible proportion du volume racinaire (8%) était attribuable aux plus grosses racines (Figure 6.B).
Tableau 5 Effet du régime hydrique sur le volume racinaire des trois espèces S. discolor, S. eriocephala et S. interior sous trois régimes hydriques, soit en conditions de sécheresse, de capacité au champ et d’inondation.
Variables Degrés de liberté indépendante Volume racinaire F. value P. value Bloc 5 - - R.h. 2 0,67 0,53 Espèces 2 Erreur A 20 Grosseur 2 Espèces*R.h. 4 Grosseur *Espèces 4 Grosseur *R.h. 4 Grosseur*Espèces*R.h. 2 Erreur B 30 7,77 0,0006* - - 45,25 <,0001* 0,50 0,74 1.,59 0,17 0,66 0,62 0,63 0,75 Total (UE) 75 Erreur échant 32 Total échant 107 - -
1.5 Discussion
1.5.1 Effet des régimes hydriques
Contrairement à notre hypothèse, la majorité des variables de croissance n’ont pas été influencées par les régimes hydriques. En effet, seule la longueur racinaire a réagi à l’effet du régime hydrique, avec une réponse variable selon l’espèce. Les deux espèces associées au haut et milieu de talus (S. discolor et S. eriocephala) ont présenté des longueurs racinaires similaires, plus faibles en condition d’inondation comparativement aux conditions de sécheresse ou de capacité au champ alors que S. interior a montré un comportement contraire : les conditions d’inondation ont favorisé des racines plus longues chez cette espèce de bas de talus. Ce résultat contraste avec la littérature. En effet, plusieurs études ont montré que la longueur racinaire et la biomasse racinaire étaient influencées négativement par les conditions de sécheresse et d’inondation ( Parent et al., 2008; Pezeshki et al., 1998; Wikbergi & ögreni, 2007). En revanche, ces études ont généralement porté sur des espèces de Salix hybrides ainsi qu’une espèce de Salix présentant une tolérance certaine à l’inondation (S. nigra). Elles sont toutes des espèces arbustives et se localisent à différents niveaux de talus en milieux naturels.
Selon Glinski (1994), la condition d’inondation provoque une réduction de concentration d’oxygène dans le sol créant ainsi une condition anaérobique qui limiterait la croissance racinaire et, à la longue, celle de la plante entière. Elle empêche ainsi une bonne aération des racines, surtout les racines fines, qui alimentent la plante en eau et en éléments nutritifs (Imada et al., 2008). Ce sont probablement ces mécanismes qui expliquent pourquoi le S. eriocephala a présenté dans cette étude, une longueur racinaire réduite en condition d’inondation par rapport à la condition de capacité au champ. Toutefois, certaines espèces, comme le S. interior et le S. discolor, ont développé des mécanismes d’adaptation leur
la période d’inondation (Teresa et al., 2013). Toutefois, nous avons comparé la biomasse et longueur racinaire des racines fines, moyennes et grosses, en fonction de trois zones dans le sol sans trouver de différence significative : les espèces testées n’ont pas produit plus de racines fines dans la partie supérieure (résultats non présentés). Un autre mécanisme d’adaptation à l’inondation chez les saules consiste à accroître leur surface foliaire (Doffo et al., 2017). Cela permet d’augmenter la quantité de stomates afin de maximiser la captation d’oxygène dans l’air, en compensation du manque d’oxygène dans le sol généré par l’inondation. La durée ou les conditions d’expérimentation présentées ici n’ont pas permis d’observer une telle réponse chez les trois espèces de saules testées.
Pour sa part, le stress de sécheresse a pour effet de diminuer l’activité photosynthétique et la conductance stomatique, ce qui peut entraîner une diminution de la croissance de la plante (Beyschlag et al., 1990). Effectivement, la diminution du contenu en eau du sol en deçà du point de flétrissement temporaire (moins de 5% pour un sol sableux) et qui correspond à une sécheresse modérée peut réduire le potentiel photosynthétique de 25 % au-delà de 21 jours de sècheresse (Ambrose et al., 2015) par rapport à la condition de capacité au champ. Toutefois, l’effet du stress de sécheresse n’affecte pas de la même façon les biomasses aérienne et racinaire : la sécheresse a généralement un effet rapide sur la croissance des racines, alors que cet effet est plus long à se faire sentir au niveau de la croissance des parties aériennes (Li et al., 2004). Cela s’explique par la présence de réserve d’eau (eau facilement utilisable) que la plante peut utiliser lorsque le stress hydrique commence à se faire sentir. Dans le cadre de cette étude, l’absence de réponse pour toutes les variables de croissance au stress hydrique de sécheresse s’explique par le fait que nous n’avons pas réussi à créer des conditions assez « sèches ». En effet, le point de flétrissement n’a jamais pu être atteint en raison de la difficulté de drainer entièrement les bacs et de l’humidité relative élevée dans la serre (nécessaire à une autre expérience simultanément à la nôtre). De façon générale, nous estimons que l’absence de réponse des différentes variables de croissance aux stress hydriques est probablement due en grande partie à la durée de l’expérience qui n’a été que de deux mois. En effet, d’autres études qui ont pu constater les effets des stress hydriques sur la production des variables de croissances s’échelonnaient sur une durée de trois mois (Imada et al., 2008) et une année (Ambrose et al., 2015).
1.5.2 Variation entre les espèces
S. eriocephala . La biomasse la plus importante de toutes les trois espèces a été produite par le S. eriocephala. Ce résultat peut en partie s’expliquer par la dominance plus prononcée du primordium apical chez cette espèce comparée aux primordium latéraux (Doffo et al., 2017). Au niveau du primordium apical se produit une hormone de croissance appelé auxine, favorisant la croissance verticale d’où l’émergence de nouvelles tiges de façon régulière et rapide (Lapointe, 2007). Le S.interior qui dispose de la même caractéristique a cependant produit deux fois moins de biomasses aériennes que le S.eriocephala. Il reste que c’est l’espèce n’ayant pas de dominance du primordium apical, le S.discolor, qui a produit la plus faible biomasse et même de 4 et 1,5 fois inférieure à celle produite par le S. eriocephala et S. interior. Ce résultat concorde avec une autre étude dans laquelle les auteurs ont constaté que le S.eriocephala a produit une biomasse et une longueur racinaire nettement supérieure au S.interior et le S.discolor.
S. discolor. La plus petite biomasse des trois espèces a été produite par le S. discolor. Aussi, selon les observations faites au cours de la période de l’expérimentation, la reprise fut très lente chez cette espèce en comparaison avec les deux autres espèces. Nous attribuons ce résultat au manque de tolérance de cette espèce à s’établir dans un sol moins fertile, comme le sable (Mosseler & Major, 2014). Nous n’avons apporté dans le cadre de notre étude qu’une très petite dose de fertilisant une seule fois (500 mL de NPK 20-20-20 liquide), ce qui n’était probablement pas suffisant pour stimuler la croissance de la biomasse pour cette espèce. Par ailleurs, les stress hydriques n’ont pas eu d’effet sur la longueur racinaire du S.discolor, ce qui ne concorde pas avec l’écologie de l’espèce qui colonise préférentiellement les hauts de talus (Mosseler et al., 2017). Toutefois, nous pouvons constater une certaine tendance à avoir une longueur racinaire moins élevée en conditions d’inondation pour le S. discolor, quoique les différences n’étaient pas significatives. Il est aussi possible qu’il puisse pousser
apical (Doffo et al., 2017; Wikbergi & Ögreni, 2007). Toutefois, cette espèce est celle ayant produit le plus de biomasse aérienne par rapport à sa biomasse racinaire : elle a montré un ratio biomasse aérienne/racinaire 1,7 et 3,15 fois plus élevé que S.eriocephala et S.discolor. Nous attribuons ce résultat au fait que les conditions expérimentales de notre étude n’ont pas permis au système racinaire de se déployer pleinement puisque chez cette espèce, les racines se développent à l’horizontale (Mosseler & Major, 2015). Les racines des boutures du S. interior étaient ici soumises à un espace assez restreint ne favorisant pas leur développement maximal comme en milieu naturel où des colonies de S. interior peuvent se développer à la suite de longues ramifications racinaires issues des racines de base (Hilty, 2017). Nous avons tout de même trouvé une plus grande longueur racinaire chez le S.interior en conditions d’inondation comparativement aux conditions de sécheresse et de capacité au champ, ce qui correspond au fait que le S.interior colonise principalement les berges des rivières et les plaines inondables (Doffo et al., 2017). Cette espèce est confinée aux habitats riverains dans la majeure partie de l’Amérique du Nord où elle est d’ailleurs utilisée pour la stabilisation des milieux perturbés (Cerrillo et al., 2013).
1.6 Remerciements
Cette étude a été financée par la bourse d’études de Naren KEITA et les subventions de recherches de Monique Poulin. Nous voulons remercier le Projet de Formation pour la Sécurité Alimentaire au Mali (FASAM ) pour l’appui financier. Nous remercions aussi tous les assistants de recherche du Département de phytologie ainsi que celle du Département des sols et de génie agroalimentaire de la Faculté des Sciences de l’Agriculture et de l’Alimentation de l’Université Laval qui a participé à cette étude. Nous sommes reconnaissantes à Hélène Crépeau pour les conseils pour les analyses statistiques.
Conclusions
La présente étude a permis d’approfondir les connaissances concernant la capacité d’établissement et de croissance des boutures du S. eriocephala, S. discolor et le S. interior sous différents régimes hydriques. En effet, les trois espèces testées, soit S. discolor, S. eriocephala et S. interior ont une tolérance certaine au stress hydrique, ayant eu des taux de reprise élevés sous les trois régimes hydriques. Néanmoins, des trois espèces, le S. eriocephala est celle ayant montré le meilleur potentiel pour produire de longues racines sous les trois régimes hydriques. S. interior en revanche, est la seule espèce pour laquelle la longueur racinaire fut favorisée en conditions d’inondation par rapport aux deux autres conditions, même si cette longueur restait inférieure à celle mesurée pour S. eriocephala. Le S. eriocephala a aussi montré la plus grande production de biomasse, tant aérienne que racinaire, ce qui en fait une espèce à privilégier dans les ouvrages de génie végétal.
Bien que le S. discolor ait montré les plus faibles productions de biomasse aérienne et racinaire, son ratio biomasse aérienne versus racinaire était celui le plus bas, ce qui indique son fort potentiel à s’ancrer dans le substrat et ainsi stabiliser le sol. Enfin, le S.interior a montré clairement sa capacité d’adaptation aux conditions d’inondation, ayant produit des racines plus longues qu’en conditions de sécheresse ou de capacité au champ. Cette espèce serait ainsi bien adaptée au bas de talus et serait à préconiser en pied de berge dans les ouvrages de génie végétal. La plupart des études portant sur le génie végétal ont utilisé d’autres espèces de Salix, telles que : S. nigra, S. viminalis ou S. elaeagnos (Fillion et al, 2009; Turhan & Serdar, 2013; Van Splunder et al, 1995). Très peu d’études ont porté sur les espèces étudiées ici. Notre étude jette ainsi les premières bases pour assurer une bonne utilisation de ces espèces dans les ouvrages de stabilisation de berges à partir des techniques de génie végétal. Cela est particulièrement vrai pour le Québec, où la pratique du génie végétal regorge de défis.
Bibliographies
1. Andreu, V., Rubio, J. L., Gimeno-Garcia, E., & Llinares, J. V. (1998). Testing three Mediterranean shrub species in runoff reduction and sediment transport. Soil & Tillage Research, 45(3-4), 441-454. doi:10.1016/s0933-3630(97)00040-8.
2. Ambrose, A.R., Baxter, W. L., Christopher S Wong, Rikke R Naesborg, Cameron B. Williams, & Todd E. Dawson. (2015). Contrasting drought-response strategies in California redwoods. Tree Physiology, 35(5), 453-469. doi:10.1093/treephys/tpv016. 3. Aravanopoulos, F. A., & Zsuffa, L. (1998). Heterozygosity and biomass production
in Salix eriocephala. Heredity, 81, 396-403.
4. Argus, G.W. (2010). Flora of North America. FNA Vol. 7 Page 27, 37. http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=242417198
5. Battany, M.C., & Grismer, M.E. (2000). Rainfall runoff and erosion in Napa Valley vineyards: effects of slope, cover and surface roughness. Hydrological Processes, 14(7), 1289-1304. doi:10.1002/(sici)1099-1085(200005)14:7<1289::Aid-hyp43>3.0.Co;2-r.
6. Bergeron, N & Roy, A.G. (1985). Le rôle de la végétation sur la morphologie d’un petit cours d’eau. Géographie physique et quaternaire, 39(3). doi:10.7202/032613ar. 7. Bernatchez, P., & Lefaivre, Denis. (2008). Effets des structures rigides de protection sur la dynamique des risques naturels côtiers : érosion et submersion. Proceedings of the 4th Canadian Conference on Geohazards : From Causes to Management, pp.08. 8. Biron, P. M., Chone, G., Buffin-Belanger, T., Demers, S., & Olsen, T. (2013).
Improvement of streams hydro-geomorphological assessment using lidar dems. Earth Surface Processes and Landforms, 38(15), 1808-1821. doi:10.1002/esp.3425.
9. Castagni, M. 2017. Etude du potentiel de bouturage Cornus sericea, Salix eriocephala et de Cornus sericea selon différents paramètres hydriques. Rappport de stage de fin de master. Laboratoire d'écologie végétale. Département de phytologie. Université Laval, Québec-Canada.. p.31.
10. Cavaille, P., Dommanget, F., Daumergue, N., Loucougaray, G., Spiegelberger, T., 11. Cerrillo T., Rodríguez, M.E., Achinelli, F., Doffo, G., & Virginia MC (2013). Do
greenhouse experiments predict willow responses to long term flooding events in the field? Bosque, 34(1), 71-79. doi:10.4067/s0717-92002013000100009
12. Choné, G. (2013). L’espace de mobilité des rivières du sud du Québec : implications pour les écosystèmes et la gestion des rivières dans une perspective de changements climatiques. (Mémoire de maîtrise, M. Sc. ), Université Concordia, Montréal: . 13. Dimitriou, I., & Aronsson, P. (2005) Willows for energy and phytoremediation in
Sweden. work in the Department of Short-Rotation Forestry, Swedish University of Agricultural Sciences, Uppsala, Sweden.pdf. pp.5.