Expansion des arbustes et pollen
Étude palynologique des sédiments lacustres récents de la région
de la rivière Boniface, Nunavik
Mémoire
Marie-Pier Denis
Maîtrise en biologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Marie-Pier Denis, 2013
iii
Résumé
L’analyse de photographies de la région de la rivière Boniface (Nunavik) a montré une forte expansion du couvert arbustif entre 1957 et 2008. L’objectif principal de ce projet était de déterminer si cette expansion est observable dans les assemblages polliniques récents des sédiments lacustres de cette région. Pour ce faire, une analyse pollinique à fine résolution temporelle a été effectuée sur les sédiments de deux lacs. Les pourcentages polliniques du bouleau glanduleux, la principale espèce responsable de la densification du couvert arbustif, sont plus élevés dans les sédiments récents de ces lacs. De plus, les influx polliniques calculés à l’aide de la datation au 210Pb montrent une augmentation des genres
Betula et Alnus depuis environ 1975. Ces influx polliniques doivent toutefois être
interprétés avec prudence en raison de difficultés liées à l’élaboration de la chronologie des sédiments. L’analyse pollinique à fine résolution semble néanmoins permettre de détecter l’expansion récente des arbustes.
v
Table des matières
Résumé ... iii
Table des matières ... v
Liste des tableaux ... vii
Liste des figures ... viii
Liste des annexes ... x
Remerciements ... xi
Chapitre 1. Introduction ... 1
1.1 Expansion des arbustes ... 1
1.1.1 Télédétection ... 1
1.1.2 Analyse de photographies aériennes ... 3
1.2 Le climat comme moteur principal de l’expansion des arbustes... 4
1.2.1 Études expérimentales ... 4
1.2.2 Études dendrochronologiques ... 4
1.3 Processus régissant l’expansion des arbustes ... 5
1.4 Analyse pollinique et strate arbustive ... 7
1.4.1 Changements historiques au sein de la strate arbustive ... 8
1.4.2 Apport de l’analyse pollinique ... 8
1.5 Objectifs du projet ... 10
Chapitre 2. Matériel et méthodes ... 11
2.1 Région d’étude ... 11
2.1.1 Le bouleau glanduleux ... 14
2.1.2 L’aulne crispé... 16
2.2 Caractéristiques des lacs sélectionnés ... 17
2.2.1 Lac Cliff ... 18
2.2.2 Lac Biscuit ... 20
2.3 Échantillonnage ... 20
2.4 Datation au plomb ... 22
vi
2.5.1 Pourcentages polliniques ... 24
2.5.2 Rééchantillonnage ... 24
2.5.3 Concentrations et influx polliniques ... 25
Chapitre 3. Résultats ... 27
3.1 Pourcentages polliniques ... 27
3.1.1 Lac Cliff ... 27
3.1.2 Lac Biscuit ... 30
3.2 Chronologie, taux de sédimentation et influx polliniques ... 32
3.2.1 Lac Cliff ... 32 3.2.2 Lac Biscuit ... 35 Chapitre 4. Discussion ... 41 4.1 Pourcentages polliniques ... 41 4.2 Influx polliniques ... 44 Conclusion ... 48 Références ... 50 Annexes ... 61
vii
Liste des tableaux
Chapitre 2.
Tableau 2.1. Principales caractéristiques des lacs Cliff et Biscuit. ... 18 Chapitre 3.
Tableau 3.1. Moyennes, écart types, maxima et minima des pourcentages polliniques et de la
concentration pollinique totale pour les lacs Cliff et Biscuit. ... 28
Tableau 3.2. Moyennes des pourcentages polliniques du bouleau dans le lac Cliff (groupées par 5 à
partir de la surface) et p-values obtenus lors du rééchantillonnage de moyennes aléatoires. Seuls les échantillons ayant une épaisseur entre 0,22 et 0,44 cm ont été utilisés. Un p-value plus petit que 0,05 (en gras dans le tableau) indique que la moyenne réelle des pourcentages polliniques des échantillons de l’intervalle de profondeur est plus élevée que les moyennes obtenues aléatoirement par rééchantillonnage. ... 29
Tableau 3.3. Moyennes des pourcentages polliniques du bouleau dans le lac Biscuit (groupées par
5 à partir de la surface) et p-values obtenus lors du rééchantillonnage de moyennes aléatoires. Seuls les échantillons ayant une épaisseur entre 0,22 et 0,4 cm ont été utilisés. Un p-value plus petit que 0,05 (en gras dans le tableau) indique que la moyenne réelle des pourcentages polliniques des échantillons de l’intervalle de profondeur est plus élevée que les moyennes obtenues aléatoirement par rééchantillonnage. ... 31
Tableau 3.4. Inventaire de plomb et date de dépôt des sédiments du lac Cliff ainsi que leurs erreurs
selon la profondeur des échantillons. ... 33
Tableau 3.5. Année de dépôt et taux de sédimentation (cm/an) des échantillons du lac Cliff en
fonction de la profondeur selon la modélisation quadratique. Les échantillons en gras sont ceux qui ont été envoyés au comptage pour le 210Pb. L’année de dépôt qui leur a été attribuée par le modèle CRS est entre parenthèses. ... 34
Tableau 3.6. Inventaire de plomb et date de dépôt des sédiments du lac Biscuit ainsi que leurs
erreurs selon la profondeur des échantillons. ... 36
Tableau 3.7. Année de dépôt et taux de sédimentation (cm/an) des échantillons du lac Biscuit en
fonction de la profondeur selon la modélisation quadratique. Les échantillons en gras sont ceux qui ont été envoyés au comptage pour le 210Pb. L’année de dépôt qui leur a été attribuée par le modèle CRS est entre parenthèses. ... 38
Tableau 3.8. Moyennes, écart types, maxima et minima des influx polliniques pour les lacs Cliff et
viii
Liste des figures
Chapitre 1.
Figure 1.1. Évolution du couvert arbustif sur une terrasse de la rivière Boniface entre 1957
(photographie aérienne, à gauche) et 2008 (image satellitaire WorldView-1, à droite). Modifié de Ropars et Boudreau (2012). ... 3
Figure 1.2. Expansion des arbustes : boucle de rétroaction positive
neige-arbustes-sol-microorganismes engendrée par le réchauffement climatique. Toutes les flèches représentent une interaction positive. La flèche pointillée relie les deux cases entre lesquelles la relation n’a pas encore été établie avec certitude. Traduit de Sturm et al. 2005. ... 6
Figure 1.3. Changements de patrons de dominance d’arbustes dans l’Arctique depuis le dernier
maximum glaciaire. Traduit de Naito et Cairns 2011. ... 9
Chapitre 2.
Figure 2.1. Principales zones de végétation du Québec et du Labrador. L’écotone forêt
boréale-toundra (ou boréale-toundra forestière) est représenté en vert pâle. L’étoile représente la région d’étude du projet. Carte originale de Payette et Rochefort (2001), modifiée par Mathieu Frégeau. ... 11
Figure 2.2. Vues en plan (gauche) et transversale (droite) de l’EFT du Québec. La vue en plan
montre la répartition des forêts (en gris) et de la toundra (en blanc) selon la latitude, jusqu’à la limite des arbres. La vue transversale montre la répartition des îlots forestiers à partir de la forêt boréale continue jusqu’à la limite arctique des arbres. Sous un scénario de réchauffement
climatique, les arbres colonisent les sommets. Sous un scénario de refroidissement du climat et en absence de feu, l’EFT reste stable, alors qu’en présence de perturbations par le feu, les arbres disparaissent du sommet des collines. Traduit de Payette et al. 2001. ... 12
Figure 2.3. Nombre de degrés-jours de croissance enregistrés entre 1971 et 2009 à la station
météorologique d’Inukjuak. La régression est significative (F[1,36] = 12,059, p < 0,001). Modifié de Dufour-Tremblay et Boudreau 2011. ... 13
Figure 2.4. Aires de répartition de deux espèces arbustives dominantes à l’écotone forêt toundra : a)
le bouleau glanduleux (Betula glandulosa Michx.) et b) l’aulne crispé (Alnus crispa (Aiton) Pursh). Modifié de Furlow 1993. ... 15
Figure 2.5. Image satellite d’une section de la rivière Boniface montrant les deux lacs
échantillonnés pour cette étude : le lac Biscuit et le lac Cliff (noms non officiels). ... 17
Figure 2.6. Bassins versants a) du lac Cliff et b) du lac Biscuit. Les bassins versants sont délimités
par la ligne noire continue. Les points verts représentent les points d’échantillonnage
(approximatifs). Les peuplements d’épinette noire, les milieux humides et les secteurs de palses sont délimités respectivement par les contours verts, bleus et bruns. ... 19
Figure 2.7. Dominance du bouleau glanduleux dans les bassins versants. A) Vue du sud-est du lac
Cliff et B) vue du nord-est du lac Biscuit. Dans les deux cas, les arbustes au premier plan sont des bouleaux glanduleux... 20
ix
Figure 2.8. Photographies des carottes du lac Cliff (A) et du lac Biscuit (B) avant le
sous-échantillonnage. ... 21
Chapitre 3.
Figure 3.1. Contenu en matière organique (%) de 11 échantillons de sédiments répartis dans la
carotte du lac Cliff, selon leur profondeur (cm). ... 27
Figure 3.2. Pourcentages polliniques des taxons de plantes vasculaires dans le lac Cliff. Les
échantillons de surface ont une épaisseur de 0,22 cm (à l’exception des deux échantillons supérieurs dont les épaisseurs sont de 0,4 cm et 0,44 cm), alors que ceux situés sous 10,98 cm ont une
épaisseur de 0,92 cm. ... 28
Figure 3.3. Contenu en matière organique (%) de 12 échantillons de sédiments répartis dans la
carotte du lac Biscuit, selon leur profondeur (cm). ... 30
Figure 3.4. Pourcentages polliniques des taxons de plantes vasculaires dans le lac Biscuit. Les
échantillons de surface ont une épaisseur de 0,22 cm (à l’exception de l’échantillon supérieur dont l’épaisseur est de 0,4 cm) et ceux situés sous 10,76 cm ont une épaisseur de 0,92 cm. ... 31
Figure 3.5. Modélisation de l’âge des sédiments selon leur profondeur pour le lac Cliff à l’aide du
210Pb. Les 6 points les plus bas sont les échantillons pour lesquels le modèle CRS a été appliqué. L’erreur associée à ces points est présentée dans le tableau 3.4. Le point supérieur (0 cm, 2011) a été ajouté pour la modélisation. La ligne continue est la courbe de tendance quadratique qui décrit le mieux la relation âge-profondeur. Elle a été utilisée pour intrapoler l’âge des sédiments qui n’ont pas été soumis à l’analyse du 210Pb. ... 33
Figure 3.6. Taux de sédimentation, influx polliniques des principaux taxons et concentration
pollinique totale pour les échantillons du lac Cliff datant des 130 dernières années. ... 35
Figure 3.7. Modélisation de l’âge des sédiments selon leur profondeur pour le lac Biscuit. Les 9
points les plus bas sont les échantillons pour lesquels le modèle CRS a été appliqué. L’erreur associée à ces points est présentée dans le tableau 3.6. Le point supérieur (0 cm, 2011) a été ajouté pour la modélisation. La ligne continue est la courbe de tendance quadratique qui décrit le mieux la relation âge-profondeur. Elle a été utilisée pour intrapoler l’âge des sédiments qui n’ont pas été soumis à l’analyse du 210Pb. ... 36
Figure 3.8. Taux de sédimentation, influx polliniques des principaux taxons et concentration
pollinique totale pour les échantillons du lac Biscuit datant des 130 dernières années. ... 39
Chapitre 4.
Figure 4.1. Relation entre la taille d’une étendue d’eau sans affluent et la proportion relative de
pollen provenant de différentes aires autour du bassin. Les lignes rouge pointillées représentent l’endroit approximatif où se situent les lacs Cliff et Biscuit selon leur taille. Figure traduite et modifiée de Jacobson et Bradshaw (1981). ... 43
x
Liste des annexes
Annexe 1. Pourcentages polliniques des arbres et arbustes du lac Cliff. Les abréviations sont
présentées à la fin du tableau. ... 61
Annexe 2. Pourcentages polliniques des herbacées du lac Cliff. Les abréviations sont présentées
à la fin du tableau. ... 63
Annexe 3. Pourcentages polliniques du pollen inconnu et des grains hors-totaux, et
concentration pollinique totale du lac Cliff. Les abréviations sont présentées à la fin du tableau. .... 65
Annexe 4. Pourcentages polliniques des arbres et arbustes du lac Biscuit. Les abréviations sont
présentées à la fin du tableau. ... 67
Annexe 5. Pourcentages polliniques des herbacées du lac Biscuit. Les abréviations sont
présentées à la fin du tableau. ... 69
Annexe 6. Pourcentages polliniques du pollen inconnu et des grains hors-totaux, et
concentration pollinique totale du lac Biscuit. Les abréviations sont présentées à la fin du
tableau. ... 71
Annexe 7. Activité du 137Cs selon la profondeur pour les échantillons a) du lac Cliff et b) du lac Biscuit... 73
xi
Remerciements
De nombreuses personnes ont contribué de près ou de loin à la réalisation de mon projet de maîtrise et au dépôt de ce mémoire. Je tiens d’abord à remercier Stéphane Boudreau, mon directeur, pour sa disponibilité et son aide au cours des deux dernières années et ce, peu importe sa localisation géographique. C’est grâce à lui que j’ai pu développer sur le terrain mon intérêt pour l’écologie. Je lui dois également mon coup de cœur pour le nord québécois. Un merci spécial à Serge Payette, sans qui les étés à Boniface n’auraient jamais existé.
Je n’aurais pas pu mener à bien mon projet sans l’aide d’Élisabeth Robert, qui m’a assistée dans l’apprentissage de l’analyse pollinique. Merci aussi à Martin Lavoie, à M. Pierre Richard, de qui les conseils au début de mon parcours m’ont permis d’avancer, ainsi qu’à Reinhard Pienitz, Claudia Zimmermann et Isabelle Laurion pour le prêt de matériel. L’expertise et la patience d’Émilie Saulnier-Talbot et de Frédéric Bouchard ont été essentielles à ma survie lors du long processus d’analyse du 210Pb de mes échantillons,
et la générosité de Christian Tardif a beaucoup simplifié mes (quelques) analyses statistiques. J’aimerais également remercier Hugo Asselin d’avoir partagé ses données avec moi.
Échantillonner des lacs dans le nord avec pas d’hélicoptère et pas de quatre-roues est une aventure en soi. Merci à Caoline et Sanda pour les kilomètres marchés dans la toundra avec sur le dos un carottier et ses poids, un extracteur, un zodiac gonflable, une pompe, deux rames, un fusil, les trousses de premiers soins, alouette! Merci aussi à Reinhard Pienitz, Jean Carignan et Robert Beauregard pour leur aide à Kuujjuarapik.
Je remercie du fond du cœur le LABoudreau et le labo Payette, de qui les différents membres ont été d’une importance capitale tout au long de mon cheminement, autant du point de vue personnel que professionnel. Merci pour votre regard critique et vos conseils, sans oublier votre capacité hors du commun à détecter une boîte de texte mal alignée ou un double espace glissé malencontreusement dans une présentation Powerpoint. Un merci spécial à tous mes collègues du 4052. Mel, le « pollen louche du courage » a ensoleillé mes bonnes et moins bonnes journées. Mathieu, les heures « perdues » à discuter féminisme,
xii
psychologie ou politique ont au moins autant contribué à préserver ma santé mentale que celles passées à discuter pollen, charbons, statistiques et rédaction.
Je remercie ma famille et mes amis biologistes, qui ont su poser juste assez de questions au sujet de mon projet, tout en réussissant à me changer les idées le moment venu. Merci à mes « autres amis » pour votre intérêt réel malgré vos domaines éloignés du mien. Les conversations nocturnes entre chercheurs m’ont aidée à mettre mon projet en perspective, et vos encouragements ont été plus précieux que ce que vous pouvez imaginer. Finalement, les mots me manquent pour exprimer ma gratitude envers Emile. Merci d’avoir été présent du début à la fin de cette aventure, de ma première session psychologiquement catastrophique à mon été d’absence nordique, en passant par les trois semaines de travail permanent qui ont précédé mon séminaire, au cours desquelles les repas sont apparus par magie dans mon assiette et la vaisselle s’est lavée d’elle-même. Je t’en dois une.
Enfin, la recherche nordique ne serait rien sans le soutien financier et les installations de nombreux organismes. Je remercie le Centre d’études nordiques, la Chaire de recherche en écologie des perturbations, le Conseil de recherches en sciences naturelles et génie du Canada, le Fonds de recherche du Québec – Nature et technologies et le Programme de formation scientifique dans le nord.
1
Chapitre 1. Introduction
Au cours des 100 dernières années, la température moyenne à la surface terrestre a augmenté de 0,74 °C ± 0,18 °C, avec une accélération notable du réchauffement dans la seconde moitié du dernier siècle (Trenberth et al. 2007). La plus grande part de ce réchauffement est attribuable à l’émission de gaz à effet de serre d’origine anthropique (Hegerl et al. 2007; Gillett et al. 2008; Christidis et al. 2012). Dans l’Arctique, l’augmentation de la température moyenne est presque le double de celle de l’ensemble du globe (Trenberth et al. 2007).
L’augmentation de la température a des répercussions particulièrement importantes dans les régions froides des hautes latitudes puisqu’on y retrouve des écosystèmes à l’intérieur desquels la température est limitante, notamment pour la reproduction sexuée et la croissance des espèces. Par conséquent, on s’attend à ce que la composition et la structure des écosystèmes arctiques et subarctiques se modifient en réponse aux changements du climat (Callaghan et Jonasson 1995). Ce type de réponse est déjà observable dans les écosystèmes toundriques. On y observe entre autres une expansion des arbustes depuis un peu plus d’une décennie, et ce dans toute la région circumpolaire (Myers-Smith et al. 2011a).
1.1 Expansion des arbustes
L’expansion des arbustes résulte d’une performance accrue des espèces arbustives (croissance latérale et horizontale et reproduction) qui se traduit par la densification de peuplements existants et la colonisation de nouveaux sites suite à l’établissement de nouveaux individus (Myers-Smith et al. 2011a). Alors que l’analyse de photographies satellitaires (télédétection) et aériennes est surtout utilisée pour décrire la dynamique spatio-temporelle du phénomène, l’identification des déterminants climatiques et écologiques repose plutôt sur des études expérimentales et dendrochronologiques.
1.1.1 Télédétection
L’indice normalisé de végétation (NDVI, Normalized Difference Vegetation Index) est un indice de la quantité de tissus photosynthétiques au sol basé sur la réflectance de la
2
radiation solaire par la végétation. Les premières études ayant utilisé le NDVI ont rapporté une augmentation de la productivité des communautés végétales sur de grandes superficies en milieu subarctique (Jia et al. 2009). Dans ce type d’analyse, la valeur du NDVI dérivée des images satellitaires est utilisée comme estimateur de la productivité de la végétation au sol (Olthof et Pouliot 2010). Le NDVI est également corrélé à la biomasse végétale aérienne et au pourcentage de recouvrement de la végétation (Raynolds et al. 2006; Raynolds et al. 2008).
Pour la période 1981-1991, les régions du globe situées entre 45°N et 70°N ont montré une importante augmentation du NDVI (Myneni et al. 1997; Myneni et al. 1998). L’augmentation du NDVI en milieu toundrique dans toute la région circumpolaire a par la suite été corroborée à plusieurs reprises pour les années 1982-2008 (Bunn et Goetz 2006; Bhatt et al. 2010; Beck et Goetz 2011). L’étude de régions plus restreintes en Alaska (Jia et
al. 2003; Stow et al. 2004; Verbyla 2008) et au Canada (Goetz et al. 2005; Jia et al. 2009;
Bhatt et al. 2010; McManus et al. 2012) a montré des résultats similaires à ceux observés pour la région circumpolaire. L’augmentation du NDVI semble être associée fortement aux zones où l’on retrouve des arbustes érigés (Silapaswan et al. 2001; Raynolds et al. 2006; Olthof et Pouliot 2010; Blok et al. 2011a; McManus et al. 2012), bien que le NDVI de certaines zones qui abritent peu ou pas d’arbustes ait aussi augmenté (Bunn et Goetz 2006; Bhatt et al. 2010; Beck et Goetz 2011).
Les résultats obtenus suite à l’analyse d’images satellites à basse résolution doivent cependant être corroborés par des études de terrain puisque de telles analyses ne permettent pas de déceler les transformations que subissent les peuplements arbustifs à l’échelle locale (Lantz et al. 2010). De plus, le NDVI peut être affecté par des facteurs géomorphologiques, hydrologiques, écologiques ou anthropiques (Walker et al. 2009). Une validation sur le terrain ou l’utilisation d’images à résolution élevée sont donc de mise lorsque l’on veut mettre en relation l’augmentation du NDVI avec celle d’un certain groupe fonctionnel ou d’une espèce en particulier (Myers-Smith et al. 2011a). Par exemple, en Sibérie, le couvert arbustif de Betula nana L. est positivement corrélé à l’augmentation du NDVI (Blok et al. 2011a).
3 1.1.2 Analyse de photographies aériennes
La comparaison de photos aériennes ou satellitaires permet d’analyser le paysage à une échelle plus fine que la télédétection, bien qu’elle nécessite aussi une validation rigoureuse sur le terrain afin de corroborer l’interprétation des images analysées.
En Alaska, la comparaison de photos obliques a montré que la strate arbustive s’est étendue au cours des 50 dernières années (Sturm et al. 2001a). L’aulne crispé (Alnus crispa (Aiton) Pursh), le bouleau nain (B. nana) et différentes espèces de saule (Salix sp.) en sont les principaux responsables (Tape et al. 2006). À Kangiqsualujjuaq (nord-est du Québec), à la limite entre les toundras forestière et arbustive, les zones de couvert ligneux érigé dense (<90%) ont augmenté de 10% entre 1964 et 2003 dans une aire de 7 km2 analysée par comparaison de photos verticales. Le bouleau glanduleux (Betula glandulosa Michx.) domine dans les parcelles où il y a eu augmentation (Tremblay et al. 2012). Cette espèce joue également un rôle prépondérant dans les changements à l’est de la baie d’Hudson, près de la limite latitudinale des arbres. Dans une région d’étude de 137 km2, l’analyse de
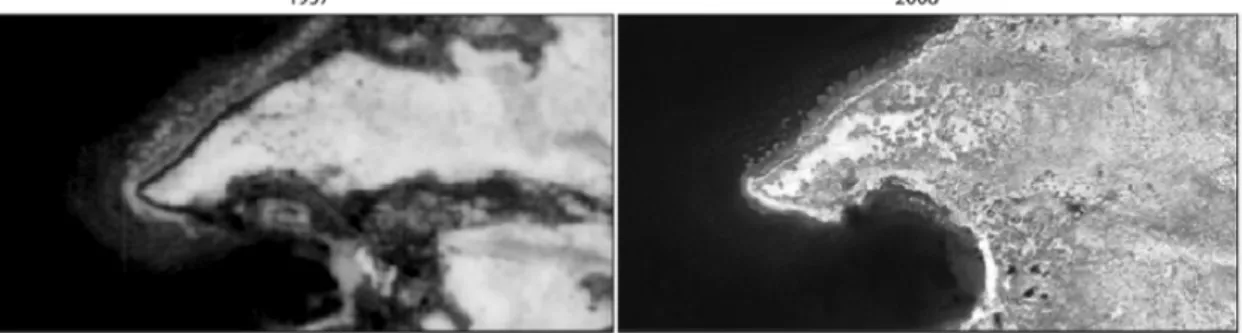
photographies aériennes et satellitaires par Ropars et Boudreau (2012) a permis d’observer que le couvert arbustif a augmenté de 21,6% sur les terrasses de basse altitude et de 11,6% sur les sommets dénudés entre 1957 et 2008 (Figure 1.1). La lacune principale de cette méthode est que l’utilisation de photos prises à des moments différents ne fournit aucun renseignement sur la variabilité interannuelle au cours du laps de temps qui sépare les photos (Callaghan et al. 2011).
Figure 1.1.Évolution du couvert arbustif sur une terrasse de la rivière Boniface entre 1957 (photographie
aérienne, à gauche) et 2008 (image satellitaire WorldView-1, à droite). Modifié de Ropars et Boudreau (2012).
4
1.2 Le climat comme moteur principal de l’expansion des arbustes
Puisque la densification et l’expansion des espèces arbustives sont des phénomènes observés à l’échelle circumpolaire, il est probable que les déterminants qui les sous-tendent agissent également à cette échelle. Pour cette raison, il a été suggéré que le climat serait le principal responsable des transformations en cours. L’effet allégué du réchauffement climatique sur la strate arbustive peut se vérifier par le biais d’études expérimentales ou dendrochronologiques.
1.2.1 Études expérimentales
De nombreux travaux ont simulé les changements climatiques afin de prédire ses effets à court, moyen et long terme sur la végétation. Les facteurs manipulés sont généralement la température, la disponibilité des nutriments et une réduction de la radiation solaire incidente pour simuler une fermeture de la voûte arbustive. Bien que les conclusions de ces études suggèrent une grande variabilité selon l’écosystème, le site et la région étudiés (Press et al. 1998; Jagerbrand et al. 2009), les arbustes décidus (particulièrement
B. nana) répondent fortement aux traitements d’augmentation de température et de
disponibilité des nutriments par une augmentation de leur biomasse (Arft et al. 1999; Bret-Harte et al. 2001; van Wijk et al. 2004; Aerts et al. 2006; Elmendorf et al. 2012). La croissance rapide de B. nana suite à ces traitements pourrait être due au fait que cette espèce possède une grande réserve de méristèmes et peut donc développer de nombreux axes végétaux (Arft et al. 1999; Bret-Harte et al. 2001). Un autre élément de réponse pourrait être qu’elle produit plus de biomasse ligneuse que les autres espèces arbustives pour un même investissement en azote (Aerts et al. 2006).
1.2.2 Études dendrochronologiques
Des études dendrochronologiques récentes ont également permis d’établir une relation entre la température au début et au milieu de la saison de croissance et la croissance annuelle des cernes et l’élongation axiale pour les espèces arbustives décidues suivantes:
Salix lanata L. (Forbes et al. 2010), Juniperus nana Willd. (Hallinger et al. 2010), Salix pulchra Cham., Betula nana (Blok et al. 2011b) et B. glandulosa (Boudreau et
5 de déterminer que la valeur du NDVI en été est fortement corrélée à la largeur des cernes de l’arbuste Salix lanata, ce qui confirme l’apport de cette espèce à l’augmentation observée du NDVI (Forbes et al. 2010; Blok et al. 2011b).
1.3 Processus régissant l’expansion des arbustes
Les corrélations établies entre le réchauffement climatique et la croissance des espèces arbustives ne nous renseignent cependant pas sur les mécanismes impliqués dans cette relation. L’augmentation de la température dans les écosystèmes nordiques pourrait promouvoir la croissance des arbustes en agissant directement sur les processus physiologiques tels que le taux de photosynthèse (Chapin 1983). De plus, une température plus élevée en début de saison accélère la fonte de la neige, ce qui allonge la saison de croissance et modifie nécessairement les conditions de recrutement et de croissance des arbustes (Myers-Smith et al. 2011a). L’allongement de la saison de croissance a d’ailleurs été observé en Amérique du Nord et en Russie (ACIA 2005; Chapin et al. 2005; Hansen et
al. 2010).
Le réchauffement observé en milieux arctiques et subarctiques a également un effet indirect important sur l’activité des microorganismes décomposeurs. L’augmentation de la température du sol entraîne un épaississement du mollisol, ce qui devrait augmenter l’activité microbienne (Blok et al. 2010). Dans des écosystèmes où la disponibilité des nutriments est un facteur limitant (Chapin et Shaver 1985), ce phénomène pourrait avoir des répercussions sur la productivité et la structure des communautés végétales.
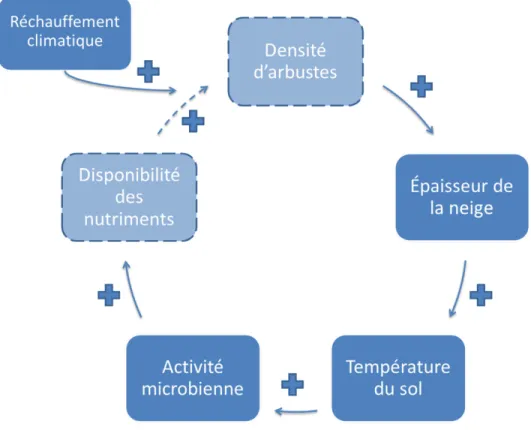
Le réchauffement climatique étant observé principalement en hiver et au printemps (Serreze et al. 2000), les événements hivernaux sont considérés comme déterminants pour expliquer l’expansion des espèces arbustives (Sturm et al. 2005). Pour cette raison, la neige est un élément clé dans la boucle de rétroaction proposée pour expliquer la performance accrue des espèces arbustives (Figure 1.2). Une température du sol élevée favorise l’activité microbienne, ce qui résulte en une disponibilité supérieure des nutriments. Ceci devrait se répercuter positivement sur la croissance des espèces arbustives au cours de la saison de croissance suivante, bien que ce lien reste à démontrer (Hallinger et al. 2010). Le développement d’une structure verticale plus complexe permet ensuite aux arbustes de
6
retenir plus de neige, ce qui se traduit par des températures du sol plus élevées en saison hivernale, favorisant ainsi une activité microbienne plus intense (Sturm et al. 2005; Cornelissen et al. 2007). Ce cycle forme une rétroaction positive qui pourrait contribuer à l’expansion des arbustes. Puisque l’épaisseur de neige requise pour isoler le sol est atteinte à une densité assez faible d’arbustes, le cycle pourrait s’enclencher rapidement (Sturm et al. 2001b).
Figure 1.2. Expansion des arbustes : boucle de rétroaction positive neige-arbustes-sol-microorganismes
engendrée par le réchauffement climatique. Toutes les flèches représentent une interaction positive. La flèche pointillée relie les deux cases entre lesquelles la relation n’a pas encore été établie avec certitude. Traduit de Sturm et al. 2005.
En plus de cette rétroaction positive, l’expansion des arbustes diminue l’albédo des surfaces terrestres (Chapin et al. 2005; Loranty et al. 2011). En effet, les arbustes absorbent l’énergie solaire à cause de leur couleur foncée, ce qui diminue la quantité d’énergie réfléchie dans l’espace et, par conséquent, augmente la température de l’air (Chapin et al. 2005).
D’autres facteurs peuvent cependant tempérer les effets d’amplification décrits ci-haut. Les événements hivernaux extrêmes (périodes de plusieurs jours avec une température
7 élevée provoquant une fonte importante de la neige) peuvent diminuer la croissance des arbustes (Bokhorst et al. 2009), alors que des précipitations nivales trop importantes écourtent la saison de croissance (Hallinger et al. 2010). L’ombrage créé par l’expansion des arbustes en été peut diminuer la température au sol, ce qui diminue les taux de décomposition et la libération des nutriments (Cornelissen et al. 2007). De plus, l’accumulation de la litière des arbustes pourrait ralentir l’activité microbienne car elle est relativement difficile à décomposer (Cornelissen et al. 2007).
Finalement, les herbivores peuvent agir comme agent modérateur dans les boucles de rétroaction positive. En effet, ils retirent une partie de la biomasse aérienne des espèces végétales, ce qui peut altérer la productivité, la structure des plantes et la composition de la communauté végétale (Tape et al. 2010). Puisque certains écosystèmes nordiques soutiennent de fortes densités d’herbivores, la pression d’herbivorisme pourrait avoir un effet opposé à celui du réchauffement climatique (Olofsson et al. 2009; Tape et al. 2010). Au contraire, le piétinement du sol par les herbivores peut accélérer l’incorporation de la matière organique et la fixation de l’azote par les microorganismes (Danell et al. 2006) en plus de fournir des lits de germination propices à l’établissement de plantules (Munier et al. 2010; Dufour-Tremblay et Boudreau 2011).
Dans certaines régions, des agents autres que le climat semblent être les moteurs de l’expansion du couvert arbustif. Par exemple, le feu, le réseau hydrographique, la dégradation du pergélisol et les activités humaines peuvent créer des microsites propices à l’établissement des arbustes et ainsi contribuer à leur expansion (Myers-Smith et al. 2011a; Myers-Smith et al. 2011b).
1.4 Analyse pollinique et strate arbustive
Dans les régions où les arbustes se densifient, la production pollinique totale de ces derniers devrait augmenter, puisque le pollen pourra être produit par un plus grand nombre d’individus. Si tel est le cas, l’expansion des arbustes devrait s’observer dans les assemblages polliniques conservés dans les séries stratigraphiques, que ce soit dans les colonnes de tourbe ou les sédiments lacustres.
8
1.4.1 Changements historiques au sein de la strate arbustive
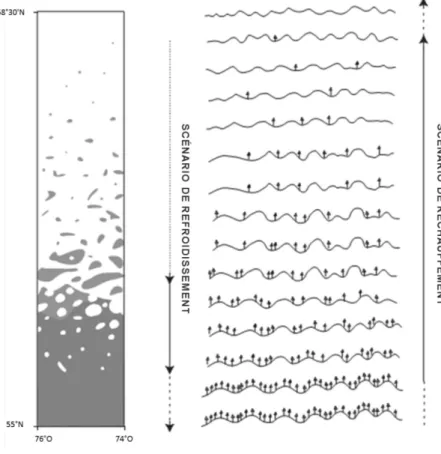
Différentes études paléoécologiques ont permis de mettre en relief des épisodes d’expansion des arbustes reliés aux conditions climatiques au cours des derniers millénaires. Une mise en commun des analyses polliniques dans toute la région circumpolaire a permis de retracer les fluctuations d’abondance de la strate arbustive depuis la dernière glaciation jusqu’à aujourd’hui (Figure 1.3). Le registre fossile montre par exemple qu’à la suite du dernier maximum glaciaire, la toundra herbacée et arbustive s’est installée en Alaska. Le paysage a ensuite été dominé successivement par le bouleau et l’aulne avant d’en arriver aux assemblages polliniques actuels d’aulne et de toundra herbacée (Anderson et Brubaker 1994; Naito et Cairns 2011). Dans la même région, les analyses polliniques et de macrofossiles montrent que les feux ont déjà été fréquents dans les étendues de bouleau glanduleux (Higuera et al. 2008; Higuera et al. 2011).
Ces exemples illustrent bien la capacité de l’analyse pollinique à détecter des changements régionaux importants au sein des communautés végétales, incluant ceux qui impliquent les espèces arbustives. L’interprétation des données palynologiques doit toutefois être réalisée avec prudence.
1.4.2 Apport de l’analyse pollinique
En palynologie que les pourcentages de représentation d’un taxon à une époque donnée ne reflètent pas directement son abondance (Lavoie 2001). En effet, la production pollinique varie énormément d’une espèce à l’autre selon son mode de pollinisation. La production montre également des variations interannuelles, et dépend notamment de l’âge de l’individu, de son rythme de floraison et des conditions climatiques (Pons 1970; Faegri et Iversen 1975; Lavoie 2001; Lezine 2008). La quantité de pollen déposée par unité de surface dépend donc de l’abondance de l’espèce dans la région, mais aussi du mécanisme de dissémination du pollen et de la production absolue de pollen (Seppa 2007). De plus, les pourcentages polliniques calculés pour une espèce dans un échantillon donné dépendent directement de la quantité de pollen déposé dans les sédiments par les autres espèces. Cependant, si l’on procède à la datation des sédiments, on peut utiliser le taux de sédimentation pour calculer les influx polliniques des différents taxons d’intérêt. L’influx
9
Figure 1.3.Changements de patrons de dominance d’arbustes dans l’Arctique depuis le dernier maximum glaciaire. Traduit de Naito et Cairns 2011.
Espèces arbustives dominantes Couvert végétal non différencié
Toundra herbacée Toundra de mousses et lichens
Toundra
10
pollinique (taux d’accumulation pollinique) est une mesure du nombre de grains de pollen par unité de surface et de temps qui présente l’avantage de rendre les données polliniques de chaque taxon indépendantes de celles des autres taxons (Lavoie 2001; Lezine 2008).
S’il est possible de retracer les fluctuations passées des arbustes à l’aide du pollen, est-il possible de faire la même chose pour les fluctuations récentes, en utilisant une résolution temporelle très fine? Suivre l’évolution de l’expansion des arbustes avec l’analyse pollinique permettrait d’ajouter un type de preuves aux connaissances récentes dans ce domaine. Puisque l’analyse pollinique fournit de l’information sur les assemblages régionaux de la végétation (Lavoie 2001), l’échelle à laquelle elle nous renseigne est semblable à celle de l’analyse de photographies. Dans les régions où les photos historiques de qualité ne sont pas disponibles, l’analyse pollinique pourrait permettre de rendre compte de l’évolution locale de l’expansion des arbustes. Elle comporte en outre l’avantage de générer une information continue, contrairement aux analyses qui étudient deux ou trois photographies et doivent considérer que ces dernières sont représentatives de l’évolution du paysage. Finalement, l’analyse pollinique a le potentiel de renseigner directement sur les groupes fonctionnels impliqués dans les changements, et parfois même sur les espèces. 1.5 Objectifs du projet
L’objectif principal de ce projet est de déterminer si l’expansion récente des arbustes dans la région de la rivière Boniface est détectable dans les assemblages polliniques conservés dans les sédiments lacustres. Pour ce faire, une analyse pollinique a été effectuée à très fine résolution temporelle sur des sédiments provenant de deux lacs situés à proximité de la limite latitudinale des arbres. À notre connaissance, cette étude est la première analyse pollinique à utiliser une résolution temporelle aussi fine. Les objectifs spécifiques de ce projet sont 1) quantifier les variations des pourcentages polliniques des principaux taxons dans les 20 cm supérieurs des sédiments lacustres, et 2) quantifier les variations des influx polliniques des principaux taxons depuis les 150 dernières années. Puisqu’il a été démontré que le bouleau glanduleux est le principal responsable de l’expansion des arbustes dans cette région (Ropars et Boudreau 2012), notre hypothèse de recherche est que les pourcentages polliniques et les influx polliniques du genre Betula seront plus élevés dans les sédiments de surface.
11
Chapitre 2. Matériel et méthodes
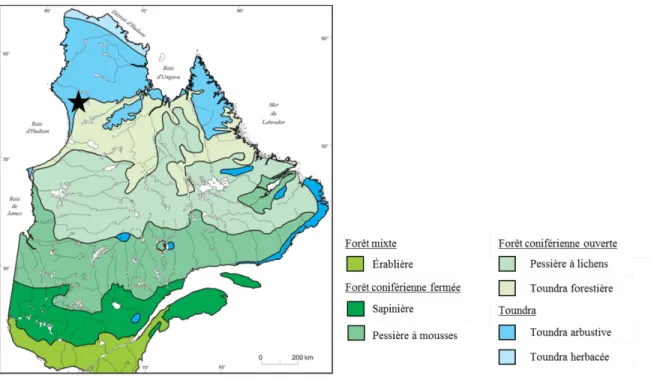
2.1 Région d’étude
L’écotone forêt boréale-toundra (EFT) est la zone de transition entre la forêt boréale et la toundra arbustive (Figure 2.1). Au Québec, il est caractérisé par une mosaïque de peuplements forestiers et de peuplements de toundra arbustive et herbacée dont l’abondance relative varie le long d’un gradient latitudinal, entre le 55ème et le 58ème
parallèle (Figure 2.2). Dans la portion méridionale de l’EFT, les peuplements forestiers dominent le paysage alors que les îlots de toundra sont retrouvés sur les sommets de haute altitude. À la limite nordique des arbres, les peuplements forestiers sont confinés aux sites humides et protégés du vent, alors que les peuplements de toundra occupent le reste du paysage (Payette et al. 2001).
Figure 2.1. Principales zones de végétation du Québec et du Labrador. L’écotone forêt boréale-toundra (ou
toundra forestière) est représenté en vert pâle. L’étoile représente la région d’étude du projet. Carte originale de Payette et Rochefort (2001), modifiée par Mathieu Frégeau.
L’épinette noire (Picea mariana (Mill.) BSP) domine les peuplements forestiers de l’EFT et forme la limite des arbres au nord-ouest du Québec. Cette espèce se régénère par graines dans les sites forestiers, alors que le marcottage (mode de croissance clonale associé à l’enracinement de branches basses) assure son maintien dans les sites plus exposés. Dans
12
ces sites, l’épinette noire est souvent trouvée sous forme de krummholz, i.e. sous forme arbustive (Payette 1993).
Figure 2.2.Vues en plan (gauche) et transversale (droite) de l’EFT du Québec. La vue en plan montre la
répartition des forêts (en gris) et de la toundra (en blanc) selon la latitude, jusqu’à la limite des arbres. La vue transversale montre la répartition des îlots forestiers à partir de la forêt boréale continue jusqu’à la limite arctique des arbres. Sous un scénario de réchauffement climatique, les arbres colonisent les sommets. Sous un scénario de refroidissement du climat et en absence de feu, l’EFT reste stable, alors qu’en présence de perturbations par le feu, les arbres disparaissent du sommet des collines. Traduit de Payette et al. 2001.
Les îlots de toundra sont plus diversifiés que les îlots forestiers. Ils sont dominés par différentes espèces de lichens, de bryophytes, d’herbacées et d’arbustes. Le bouleau glanduleux, l’aulne crispé (Alnus crispa (Ait.) Pursh) ainsi que différentes espèces de saule (Salix spp.) et d’éricacées (Vaccinium spp. Rhododendron spp. Empetrum nigrum L.) composent la strate arbustive (Gajewski 1991; Gamache et Payette 2005). De ces espèces arbustives, certaines dominent à la fois le paysage et les différents diagrammes polliniques de l’EFT produits jusqu’à ce jour (Gajewski 1991; Gajewski et al. 1993; King 1993). Dans la région de la rivière Boniface, c’est notamment le cas du bouleau glanduleux, espèce
13 structurante des îlots de toundra, et de l’aulne crispé (Gajewski 1991; Ropars et Boudreau 2012).
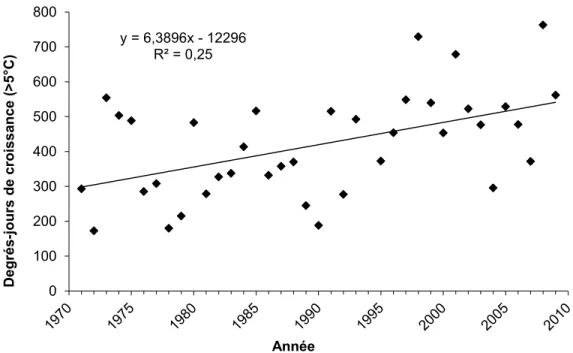
Les lacs à l’étude se situent dans la sous-zone arbustive de la toundra forestière, dans la région de la rivière Boniface (57°45’N, 76°20’O), à environ 10 km au sud de la limite des arbres (Figure 2.1). Les données climatiques à long terme disponibles pour cette région sont celles de la station d’Inukjuak, à 135 km au nord-ouest de la rivière Boniface. Elles sont représentatives des conditions climatiques de la région de Boniface (Wang et al. 2001). La température annuelle moyenne est de -7 °C pour la période 1971-2000 et les précipitations moyennes annuelles sont d’environ 460 mm, dont 45 % tombent sous forme de neige (Environnement Canada 2012). La durée moyenne de la période sans gel est de 60 jours (Vallée et Payette 2007). Selon ce que rapportent Dufour-Tremblay et Boudreau (2011), le nombre de degrés-jours de croissance à Inukjuak est passé de 333 ± 142 pour la période 1971-1979 à 517 ± 142 entre 2001 et 2009 (Figure 2.3). À la rivière Boniface, deux stations météorologiques installées en milieux forestier et toundrique révèlent une température annuelle moyenne de -5,2 °C entre 1988 et 2008.
Figure 2.3. Nombre de degrés-jours de croissance enregistrés entre 1971 et 2009 à la station météorologique
d’Inukjuak. La régression est significative (F[1,36] = 12,059, p < 0,001). Modifié de Dufour-Tremblay et
Boudreau 2011. y = 6,3896x - 12296 R² = 0,25 0 100 200 300 400 500 600 700 800 D eg rés -jo ur s d e cr oi ssan ce ( >5 °C) Année
14
Située sur le bouclier canadien, la région de la rivière Boniface est une pénéplaine qui s’est développée sur des affleurements granito-gneissiques d’âge précambrien (Allard et Rousseau 1999). Dans cette région, la déglaciation est survenue vers 7600 ans BP, puis la transgression marine de la mer de Tyrrell a eu lieu vers 7200 ans BP (Dyke 2004). La région est aujourd’hui caractérisée par des collines basses (sommets situés à environ 200 m au-dessus du niveau de la mer), ainsi que par des vallées et des lacs qui se jettent dans la rivière Boniface (Allard et Rousseau 1999).
Les communautés végétales actuelles de la région d’étude ont été modelées par l’interaction entre les feux et le refroidissement du climat depuis la fin de l’Holocène (Arseneault et Payette 1997). Au cours du dernier millénaire, l’absence de régénération après feu suite à la détérioration du climat aurait causé la conversion de peuplements forestiers en peuplements toundriques (Payette et Gagnon 1985). Pour cette raison, les pessières à mousses se retrouvent aujourd’hui dans les dépressions humides et protégées du vent, alors que les sommets exposés et les flancs de collines abritent les pessières à lichens, les krummholz et la toundra arbustive ((Payette et al. 2008)).
2.1.1 Le bouleau glanduleux
Le bouleau glanduleux est l’espèce arbustive structurante de la toundra à l’EFT. C’est un arbuste pouvant atteindre une hauteur de 2,5 m. Son aire de répartition couvre les régions nordiques de l’Amérique du Nord, ainsi que la côte ouest nord-américaine jusqu’au nord de la Californie (Figure 2.4a; Furlow 1993). Il forme des fourrés plus ou moins denses et croît généralement dans des sols acides et pauvres en nutriments. Le bouleau glanduleux est intolérant à l’ombre et peut supporter des pressions soutenues d’herbivorisme (de Groot et Wein 1999; Champagne et al. 2012). Il est un proche parent du bouleau nain, un arbuste qui répond fortement aux expériences de réchauffement et de fertilisation (de Groot et al. 1997).
La croissance clonale du bouleau glanduleux se fait par le biais de racines adventives qui se développent sur les tiges (Weis et Hermanutz 1988) et de bourgeons dormants qui se développent sur les racines et les rhizomes (Tollefson 2007). La
15 reproduction sexuée chez le bouleau glanduleux est fréquente, mais diminue le long d’un gradient latitudinal jusqu’à la limite nord de sa répartition (Weis et Hermanutz 1988). a)
b)
Figure 2.4. Aires de répartition de deux espèces arbustives dominantes à l’écotone forêt toundra : a) le
bouleau glanduleux (Betula glandulosa Michx.) et b) l’aulne crispé (Alnus crispa (Aiton) Pursh). Modifié de Furlow 1993.
Le bouleau glanduleux est une espèce monoïque. Les chatons femelles apparaissent quand les feuilles commencent à s’ouvrir au printemps, après la fonte de la neige, alors que
16
les chatons mâles sont formés vers le milieu de la saison de croissance (de Groot et al. 1997; Tollefson 2007). Le pollen est anémophile et est produit en grande quantité (Weis et Hermanutz 1988). Le genre Betula est d’ailleurs l’un des taxons dominants des spectres polliniques modernes dans la toundra forestière québécoise (Gajewski 1991). Il n’y a pas d’autopollinisation chez cette espèce (Weis et Hermanutz 1993). Les graines se forment entre juillet et octobre et peuvent persister sur l’individu pendant l’hiver. La dissémination des graines commence à l’automne et se termine au printemps (Tollefson 2007).
2.1.2 L’aulne crispé
L’aulne crispé est lui aussi répandu dans l’EFT. Cet arbuste peut atteindre jusqu’à trois mètres de hauteur. En Amérique du nord, son aire de répartition couvre l’est et le centre du Canada (Figure 2.4b). On le retrouve aussi dans l’est de la Russie (Furlow 1993). Il croît individuellement ou en petits bosquets, principalement le long des cours d’eau et sur les affleurements rocheux, mais également en toundra ou en forêt lorsque le couvert forestier est relativement ouvert. La capacité de l’aulne crispé à s’associer à des bactéries fixatrices d’azote en fait une espèce pionnière. On le retrouve souvent sur les sols acides des régions récemment déglacées (Houle 2001; Rhoades et al. 2001).
L’aulne crispé se reproduit de façon clonale à partir d’organes souterrains, particulièrement suite à des perturbations comme le feu (Rook 2002). La reproduction sexuée reste toutefois le mode principal de régénération de l’espèce. Les graines germent bien sur des sols perturbés. L’aulne crispé est monoïque. Les chatons femelles s’allongent en même temps que l’ouverture des feuilles au printemps. Les chatons mâles sont produits en août, vers la fin de la saison de croissance. Ils s’allongent rapidement au printemps lorsque la température augmente et libèrent leur pollen anémophile en quelques jours, puis sèchent et tombent. Le pollen du genre Alnus est produit en grande quantité, ce qui mène à une surreprésentation systématique de ce taxon dans les assemblages polliniques modernes (King 1993). Après la pollinisation, les chatons femelles grossissent jusqu’à la fin de l’été. Il y a chevauchement entre la production de fruits et la production des chatons pour l’année suivante. Les graines sont disséminées de septembre à avril (Houle 2001; Rook 2002).
17
2.2 Caractéristiques des lacs sélectionnés
Avant l’échantillonnage sur le terrain, l’analyse d’une image satellite de la région de la rivière Boniface a permis de repérer une dizaine de lacs potentiels pour le carottage des sédiments lacustres. Afin d’être identifiés comme potentiels, les lacs devaient être exempts d’affluents, facilement accessibles et de petite taille. Les lacs ainsi pré-identifiés ont été visités afin de vérifier s’ils répondaient à tous ces critères de sélection. Seuls les lacs ayant une profondeur de plus de 2 m ont été retenus afin de s’assurer que les sédiments de surface ne sont pas perturbés par le gel complet de la colonne d’eau en saison hivernale. Suite à cette vérification sur le terrain, deux lacs situés au sud de la rivière Boniface et à 4 km l’un de l’autre ont été sélectionnés pour cette étude : le lac Cliff et le lac Biscuit (noms non officiels, Figure 2.5).
Figure 2.5. Image satellite d’une section de la rivière Boniface montrant les deux lacs échantillonnés pour
cette étude : le lac Biscuit et le lac Cliff (noms non officiels).
Toutes les données relatives aux bassins versants des lacs ont été obtenues à partir d’une image satellitaire WorldView-1 Standard Ortho-Ready panchromatic prise le 15 juillet 2008 et couvrant une région de 151 km2. L’image a été orthorectifiée afin de corriger les déformations topographiques (résolution : 0,5 m). Les mesures ont été effectuées avec le logiciel ArcGIS 9 (ESRI).
18
2.2.1 Lac Cliff
Le lac Cliff se situe à une altitude de 132 m selon le modèle numérique d’élévation de terrain de la région généré par Ressources naturelles Canada (résolution : 15 m; Tableau 2.1). Il a une superficie totale de 17 146 m2 et un périmètre de 660 m. Son bassin versant
couvre une superficie de 302 320 m2. Le lac Cliff est un lac encaissé qui ne possède pas d’affluents. Son seul exutoire se trouve au nord-ouest et traverse un milieu humide avant de se déverser dans un second lac de taille plus importante. Le lac est entouré de collines sur lesquelles le roc nu affleure par endroits. La colline la plus élevée culmine à 194 m d’altitude, au sud-ouest du lac. En excluant la superficie du lac, les peuplements d’épinette noire et les milieux humides occupent respectivement 10 % et 1 % du bassin versant (Figure 2.6a; Tableau 2.1). Toutefois, il est difficile de différencier la toundra arbustive, la toundra herbacée et les affleurements rocheux sur l’image satellitaire. Les observations effectuées sur le terrain montrent néanmoins que le bouleau glanduleux est répandu dans tout le bassin versant, excepté dans les peuplements forestiers denses (Figure 2.7a).
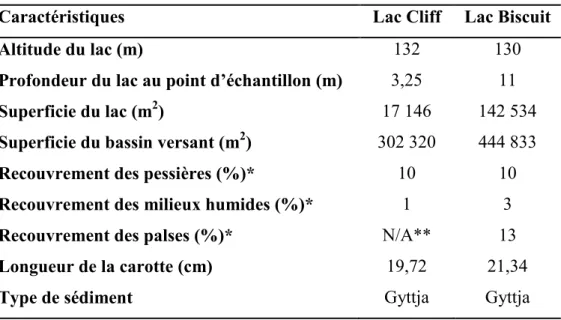
Tableau 2.1.Principales caractéristiques des lacs Cliff et Biscuit.
Caractéristiques Lac Cliff Lac Biscuit
Altitude du lac (m) 132 130
Profondeur du lac au point d’échantillon (m) 3,25 11
Superficie du lac (m2) 17 146 142 534
Superficie du bassin versant (m2) 302 320 444 833
Recouvrement des pessières (%)* 10 10
Recouvrement des milieux humides (%)* 1 3
Recouvrement des palses (%)* Longueur de la carotte (cm)
N/A** 19,72
13 21,34
Type de sédiment Gyttja Gyttja
*La surface recouverte par chaque type de milieu a été calculée sur ArcGIS à l’aide d’une image satellite de la région d’étude et de reconnaissance sur le terrain. Le pourcentage de recouvrement a été calculé par rapport à la superficie du bassin versant excluant la superficie du lac.
19 a)
b)
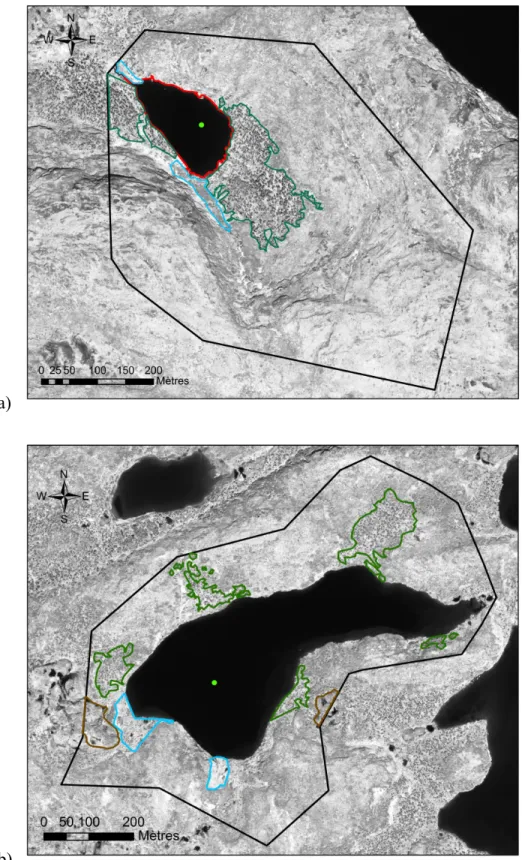
Figure 2.6. Bassins versants a) du lac Cliff et b) du lac Biscuit. Les bassins versants sont délimités par la
ligne noire continue. Les points verts représentent les points d’échantillonnage (approximatifs). Les
peuplements d’épinette noire, les milieux humides et les secteurs de palses sont délimités respectivement par les contours verts, bleus et bruns.
20
Figure 2.7. Dominance du bouleau glanduleux dans les bassins versants. A) Vue du sud-est du lac Cliff et B)
vue du nord-est du lac Biscuit. Dans les deux cas, les arbustes au premier plan sont des bouleaux glanduleux.
2.2.2 Lac Biscuit
Le lac Biscuit est situé à une altitude d’environ 130 m selon le modèle numérique d’élévation de terrain de Ressources naturelles Canada (résolution : 15 m; Tableau 2.1). Sa superficie totale est de 142 534 m2 (soit plus de huit fois celle du lac Cliff) et son périmètre
est de 2098 m.Son bassin versant couvre 444 833 m2, soit environ 1,5 fois la superficie du bassin versant du lac Cliff. Le lac Biscuit est un lac de tête et possède un seul exutoire. Il se déverse à l’est dans un second lac. Le lac Biscuit est entouré de plusieurs sommets (point culminant : 156 m) ainsi que de quelques peuplements forestiers dans les vallées. Si l’on exclut l’aire du lac, les peuplements d’épinette noire et les milieux humides occupent respectivement environ 10 % et 3 % du bassin versant (Figure 2.6b; Tableau 2.1). Des chapelets de palses au sud-est et au sud-ouest du lac couvrent 3 % du bassin versant. Comme pour le lac Cliff, l’analyse de l’image satellite n’a pas permis de différencier la toundra arbustive, la toundra herbacée et le roc nu, mais les observations sur le terrain ont confirmé que le bouleau glanduleux domine le paysage (Figure 2.7b).
2.3 Échantillonnage
Les deux lacs ont été carottés à l’aide d’un carottier à gravité (Aquatic Research Instruments, diamètre interne des tubes : 6,8 cm) à partir d’une petite embarcation pneumatique immobilisée sur l’eau à l’aide de deux ancrages. Une carotte courte a été
21 prélevée dans chacun des lacs. L’inspection des carottes à travers le tube d’échantillonnage ne laissait pas présumer de brassage de sédiments (aucune stratification évidente; Figure 2.8).
Figure 2.8. Photographies des carottes du lac Cliff (A) et du lac Biscuit (B) avant le sous-échantillonnage. Le Lac Cliff a été carotté le 11 juillet 2011. La carotte de 19,72 cm a été prélevée à une profondeur de 3,25 m (57°43’24,3344’’N, 76°10’20,5466’’O), à l’endroit qui semblait être le plus profond (Tableau 2.1; Figure 2.6a). La transparence du lac a été évaluée à 2,75 m à l’aide d’un disque de Secchi. La carotte a été sous-échantillonnée aux 0,22 cm jusqu’à 10,3 cm, puis aux 0,92 cm jusqu’à 19,72 cm, à l’exception des deux premiers sous-échantillons qui mesurent 0,4 et 0,44 cm en raison de la quantité trop importante d’eau dans les sédiments de surface qui rendait impossible un échantillonnage plus fin.
Le carottage du lac Biscuit a eu lieu le 1er août 2011. Au point d’échantillonnage (57°43’28,898’’N, °15’6,1492’’O), le lac a une profondeur de 11 m et une transparence de 5,5 m (Tableau 2.1; Figure 2.6b). La carotte de sédiments mesure 21,34 cm. La carotte du lac Biscuit a été sous-échantillonnée aux 0,22 cm jusqu’à 10,08 cm de profondeur, puis aux 0,92 cm jusqu’à 21,34 cm, à l’exception du premier sous-échantillon qui mesure 0,4 cm.
Les sous-échantillons des deux lacs ont été conservés dans des sacs de plastique hermétiques de type Whirlpak à l’abri de la lumière et au frais jusqu’au moment d’être congelés pour la lyophilisation.
22
2.4 Datation au plomb
Pour chacune des carottes, 11 ou 14 échantillons (lac Cliff et lac Biscuit, respectivement) répartis sur toute la longueur ont été envoyés au laboratoire de radiochronologie de l’Université Laval pour comptage gamma du 210Pb et 137Cs. Puisque
cette analyse est non destructrice, ces échantillons ont aussi été traités pour l’analyse pollinique comme tous les autres échantillons. Les échantillons analysés pour le 210Pb ont ensuite été mis à l’étuve à 375°C pour une heure, puis à 600°C pour 6h afin de calculer leur contenu en matière organique (Carter 1993). À partir ce celui-ci, le volume des échantillons a pu être calculé (Équation 2.4).
Parmi les échantillons envoyés au comptage gamma, seuls ceux qui avaient des concentrations de 210Pb supérieures au bruit de fond ont été retenus, soit 6 échantillons pour le lac Cliff et 10 pour le lac Biscuit. Ce sont ces échantillons qui ont été utilisés pour obtenir la chronologie des deux carottes. Suite à l’analyse des courbes d’activité du 210Pb
selon la profondeur des sédiments, le modèle CRS (« constant rate of supply »), qui tient compte d’un flux atmosphérique de 210Pb non supporté constant et d’un taux de
sédimentation variable dans le temps, a été utilisé pour les deux chronologies (Appleby 2001).
La concentration de 137Cs dans les échantillons a été comptée en même temps que
celle du 210Pb. Le 137Cs est un radio-isotope artificiel issu des essais nucléaires du milieu du
20ème siècle. On détecte sa présence dans les sédiments entre 1954 et 1963, avec un pic en
1963. Il arrive que l’on observe un second pic d’activité vers 1986 en raison de la radioactivité émise lors de l’explosion de Tchernobyl (Appleby 2001). Le profil de la concentration en 137Cs permet généralement de valider la chronologie obtenue par le 210Pb,
en s’assurant que l’année 1963 telle que définie par les deux méthodes concorde. Toutefois, dans les lacs de petite superficie du nord du Canada, il arrive fréquemment que le profil de
137Cs soit déformé par la remobilisation du césium suite à son dépôt dans les sédiments
(Blais et al. 1995). Lorsque cette technique de validation de la chronologie est inutilisable, il existe une autre option. Selon les modèles théoriques, la profondeur à laquelle l’inventaire de plomb est réduit de 90% par rapport à la valeur de surface devrait correspondre à un âge d’environ 75 ans (Appleby et Oldfield 1990; Sorgente et al. 1999).
23 À partir de l’année de dépôt associée aux échantillons, la relation âge-profondeur des sédiments de chacun des lacs a été modélisée à l’aide d’une équation quadratique dans le but d’associer un âge à chacun des sédiments échantillonnés. Pour ce faire, le point 0 cm, qui correspond à l’année 2011, a été ajouté aux échantillons comptés. À partir de l’âge obtenu par la modélisation, le taux de sédimentation de chacun des échantillons a été calculé en utilisant la formule suivante :
(𝑴𝑺𝒊 − 𝑴𝑺𝒔)/(𝑨𝒔 − 𝑨𝒊) (Équation 2.1)
Où MSi : Mi-section de l’échantillon d’intérêt (cm)
MSs : Mi-section de l’échantillon situé directement au-dessus de l’échantillon d’intérêt) (cm) As : Âge de l’échantillon directement au-dessus de l’échantillon d’intérêt (années)
Ai : Âge de l’échantillon d’intérêt 2.5 Analyse pollinique
Puisqu’une résolution temporelle très fine était désirée dans le cadre de ce projet, les sous-échantillons contenaient peu de sédiments (diamètre de 6,8 cm et épaisseur de 0,22 cm pour les échantillons les plus petits). Cet aspect a compliqué l’analyse pollinique, puisqu’il rendait impossible l’utilisation traditionnelle d’un volume d’un centimètre cube de sédiments. La lyophilisation (déshydratation par la congélation suivie d’une évaporation sous vide) de l’ensemble des échantillons a donc été privilégiée, puisqu’elle permettait l’analyse pollinique et la datation au plomb des mêmes échantillons. Cette technique a déjà été utilisée avec succès pour des sédiments lacustres du Québec méridional (Houle et al. 2012). Chaque sous-échantillon humide a été pesé sur une balance de haute précision (±0,0001 g). Les échantillons ont ensuite été congelés, lyophilisés et pesés à nouveau.
Une quantité variable (entre 0,15 g et 0,60 g) de sédiments lyophilisés et bien mélangés a été traitée selon la méthode classique pour les analyses polliniques (Faegri et Iversen 1975). La masse analysée est variable en raison de la quantité variable des sédiments comprise dans chacun des sous-échantillons et de la masse volumique très différente d’un lac à l’autre. Un volume connu d’une suspension de grains d’Eucalyptus
globulus dont la concentration a été déterminée préalablement a été ajouté aux sédiments
avant les traitements, de façon à pouvoir calculer la concentration pollinique (grains/cm3)
24
Un minimum de 300 grains de pollen de plantes vasculaires terrestres a été compté au microscope à un grossissement de 400 x pour chacun des sous-échantillons et constitue la somme pollinique des échantillons. Les grains de pollen des taxons suivants ont été considérés dans le total : Picea, Pinus, Cupressacées (Thuja et Juniperus confondus),
Juglans, Acer, Populus, Larix, Salix, Betula (considéré comme arbustif en raison de la
latitude de la région d’étude), Alnus, Myrica gale, Ericaceae, Cyperaceae, Artemisia,
Ambrosia, Poaceae, Rubus chamaemorus, Polygonum sp., Chamerion angustifolium et Cornus sp. Le pollen des genres Picea et Pinus n’a pas été identifié à l’espèce. Les grains
de pollen des genres Betula ou Alnus ne pouvant être identifiés avec certitude ont été comptabilisés dans la catégorie « Betulacées non-identifiables » afin d’obtenir le plus d’information possible au sujet de la strate arbustive. Seuls les taxons les plus communs sont représentés dans les diagrammes polliniques.
2.5.1 Pourcentages polliniques
Pour calculer les pourcentages polliniques de chaque taxon, l’équation 2.2 a été utilisée :
𝑷𝑷(%) = �𝑺𝑷𝑺𝑻� × 𝟏𝟎𝟎 % (Équation 2.2)
Où PP : pourcentage pollinique
ST : somme des grains de pollen d’un taxon donné comptés dans l’échantillon SP : somme pollinique de l’échantillon
2.5.2 Rééchantillonnage
Suite au calcul des pourcentages polliniques et à l’analyse des résultats, nous étions intéressés à vérifier si les tendances d’augmentation décelées pour le bouleau à la surface étaient significatives. Pour ce faire, la moyenne des pourcentages polliniques a été calculée pour chaque groupe de cinq sous-échantillons à partir de la surface. Pour savoir si ces moyennes étaient significativement plus élevées que la variabilité observée au sein des échantillons, nous avons généré 10 000 moyennes aléatoires (groupes de 5) à partir des données de pourcentages polliniques. Seuls les pourcentages polliniques des 10 centimètres de surface ont été utilisés, car les échantillons situés plus profondément dans les carottes ont une épaisseur différente. À partir de ces moyennes aléatoires, nous avons calculé la probabilité d’obtenir une moyenne aussi élevée que la moyenne réelle de façon aléatoire.
25 Lorsque p était plus petit que 0,05, la moyenne était considérée significativement plus élevée.
2.5.3 Concentrations et influx polliniques
Pour calculer la concentration pollinique totale des échantillons, le nombre total de grains de pollen dans l’échantillon a été calculé avec l’équation 2.3 :
𝑵𝑷𝑭 =(𝑵𝑷𝑬 ×𝑺𝑷)𝑺𝑬 (Équation 2.3)
Où NPF : nombre de grains de pollen fossile dans l’échantillon
NPE : nombre de grains de pollen d’Eucalyptus ajoutés dans l’échantillon (calculé à partir de la concentration du mélange d’Eucalyptus)
SP : somme pollinique de l’échantillon (somme de tous les grains de pollen comptés dans l’échantillon, en excluant les grains hors-total)
SE : somme des grains de pollen d’Eucalyptus comptés dans l’échantillon
Le calcul de la concentration pollinique d’un échantillon exige de connaître le volume de l’échantillon humide total. Puisque l’analyse pollinique a été effectuée à partir d’une masse donnée de sédiments secs (et non pas d’un volume connu de sédiments humides), le volume humide de chaque échantillon a dû être calculé à partir de la densité des particules et des masses sèche et humide de chacun des échantillons (Avnimelech et al. 2001). Nous avons privilégié ce calcul à l’approximation d’un volume théorique à partir du diamètre et de l’épaisseur de chacun des échantillons puisque cela permettait de tenir compte de l’incertitude associée à l’extraction de sédiments très minces. En effet, puisque le carottier utilisé lors de l’extraction des échantillons n’était pas conçu pour les échantillons minces, il est probable que l’ensemble des échantillons n’ait pas une épaisseur exacte de 0,22 cm ou 0,92 cm. L’utilisation de la masse réelle de chacun des échantillons et de leur contenu en matière organique permet de contourner cette potentielle source d’erreur.
𝑽 =𝑴𝑺𝑫 + (𝑴𝑯 − 𝑴𝑺) (Équation 2.4)
Où MS : masse sèche de l’échantillon complet (g) D : densité des particules (grains/cm3)
26
Pour obtenir une concentration finale en grains de pollen/cm3, le calcul suivant a été fait selon la méthode de Houle et al. (2012) :
𝑪𝑷 = �𝑴𝑻𝑺𝑷�×𝑴𝑺
𝑽 (Équation 2.5)
Où CP : concentration pollinique (grains/cm3)
SP : somme pollinique de l’échantillon (somme de tous les grains de pollen comptés dans l’échantillon, en excluant les grains hors-total)
MT : masse des sédiments traités (g) MS : masse sèche totale de l’échantillon V : volume de l’échantillon humide
Les mêmes calculs ont été effectués pour connaître la concentration pollinique de chacun des taxons à l’intérieur des échantillons.
Finalement, les influx polliniques ont été calculés à partir de l’équation 2.6 :
𝑰𝑷 = 𝑻𝑺 × 𝑪𝑷 (Équation 2.6)
Où IP : influx pollinique (grains/cm2/année)
TS : taux de sédimentation (cm/année) CP : concentration pollinique (grains/cm3)
27
Chapitre 3. Résultats
3.1 Pourcentages polliniques
3.1.1 Lac Cliff
Les sédiments du lac Cliff sont constitués de gyttja uniforme. Aucun macroreste végétal n’a été trouvé dans les sédiments prélevés. Le contenu en matière organique moyen des 11 échantillons mesurés est de 30,1 ± 2,0 %. L’échantillon de surface a le plus haut taux de matière organique, soit 34,8 % (Figure 3.1).
Figure 3.1.Contenu en matière organique (%) de 11 échantillons de sédiments répartis dans la carotte du lac
Cliff, selon leur profondeur (cm).
Les pourcentages polliniques du genre Betula varient entre 21,6 % et 40,2 % ( = 30,4 ± 4,1 %) (Tableau 3.1; Figure 3.2; Annexes 1 à 3). Ils augmentent dans la portion supérieure de la carotte, entre 3 cm et la surface. D’ailleurs, la moyenne des pourcentages polliniques du bouleau des cinq échantillons de surface est significativement plus élevée que les moyennes calculées aléatoirement par rééchantillonnage (Tableau 3.2), ce qui prouve que la proportion du spectre pollinique occupée par le bouleau a augmenté récemment dans le lac Cliff. L’aulne est toutefois le taxon le plus représenté dans les assemblages polliniques du lac Cliff ( = 49,0 ± 4,6 %), bien que ses pourcentages polliniques diminuent faiblement dans les six centimètres superficiels. Le groupe « autres espèces arbustives », principalement constitué d’éricacées et de différentes espèces de saule, a des pourcentages polliniques relativement stables ( = 2,7 ± 1,4 %).
0 4 8 12 16 20 27 28 29 30 31 32 33 34 35 Pr of on de ur (cm ) Matière organique (%)
28
Tableau 3.1. Moyennes, écart types, maxima et minima des pourcentages polliniques et de la concentration
pollinique totale pour les lacs Cliff et Biscuit.
LAC CLIFF LAC BISCUIT
Pourcentages
polliniques (%) Moyenne ± écart type Maximum Minimum Moyenne ± écart type Maximum Minimum
Picea 6,5 ± 3,4 23,1 3,1 28,4 ± 10,8 50,2 7,7 Pinus 0,7 ± 0,5 1,9 0,0 2,2 ± 1,3 5,4 0,0 Autres arbres 0,1 ± 0,2 0,7 0,0 0,1 ± 0,1 0,3 0,0 Alnus 49,0 ± 4,6 57,7 33,8 26,1 ± 5,0 38,8 14,8 Betula 30,4 ± 4,1 40,2 21,6 24,2 ± 5,3 36,7 13,0 Autres arbustes 2,7 ± 1,4 6,3 0,3 4,6 ± 1,4 7,6 1,8 Cypéracées 3,4 ± 1,3 7,6 1,5 5,1 ± 1,6 9,3 2,1 Tubuliflores 2,3 ± 1,0 4,7 0,6 1,9 ± 1,0 4,1 0,3 Poacées 1,0 ± 0,6 2,5 0,0 1,2 ± 0,7 3,1 0,0 Autres herbacées 1,1 ± 0,7 2,9 0,3 2,0 ± 0,9 4,2 0,6 Pollen non-identifiable 4,1 ± 1,7 8,7 0,7 4,2 ± 2,0 8,8 0,3 Concentration pollinique totale (grains/cm3) 81 527 ± 26 674 172 732 33 775 66 219 ± 23 290 144 464 5 938
Figure 3.2. Pourcentages polliniques des taxons de plantes vasculaires dans le lac Cliff. Les échantillons de
surface ont une épaisseur de 0,22 cm (à l’exception des deux échantillons supérieurs dont les épaisseurs sont de 0,4 cm et 0,44 cm), alors que ceux situés sous 10,98 cm ont une épaisseur de 0,92 cm.